
Precursor-Boosted Production of Metabolites in Nasturtium officinale Microshoots Grown in Plantform Bioreactors, and Antioxidant and Antimicrobial Activities of Biomass Extracts

 ,
,  , , , ,
, , , ,  ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Preliminary Results Obtained in Agitated Cultures
2.2. The Appearance of Microshoot Cultures and Growth Rate
2.3. Phytochemical Analyses of Metabolites
2.3.1. Total Soluble Saccharide Content
2.3.2. Photosynthetic Pigment Content
2.3.3. Total GSL Content
2.3.4. UHPLC-DAD-MS/MS Analysis of GSLs
2.3.5. Total Flavonoid Content
2.3.6. Total Polyphenol Content
2.3.7. HPLC-DAD Analysis of Polyphenol Compounds
2.4. Antioxidant Activity
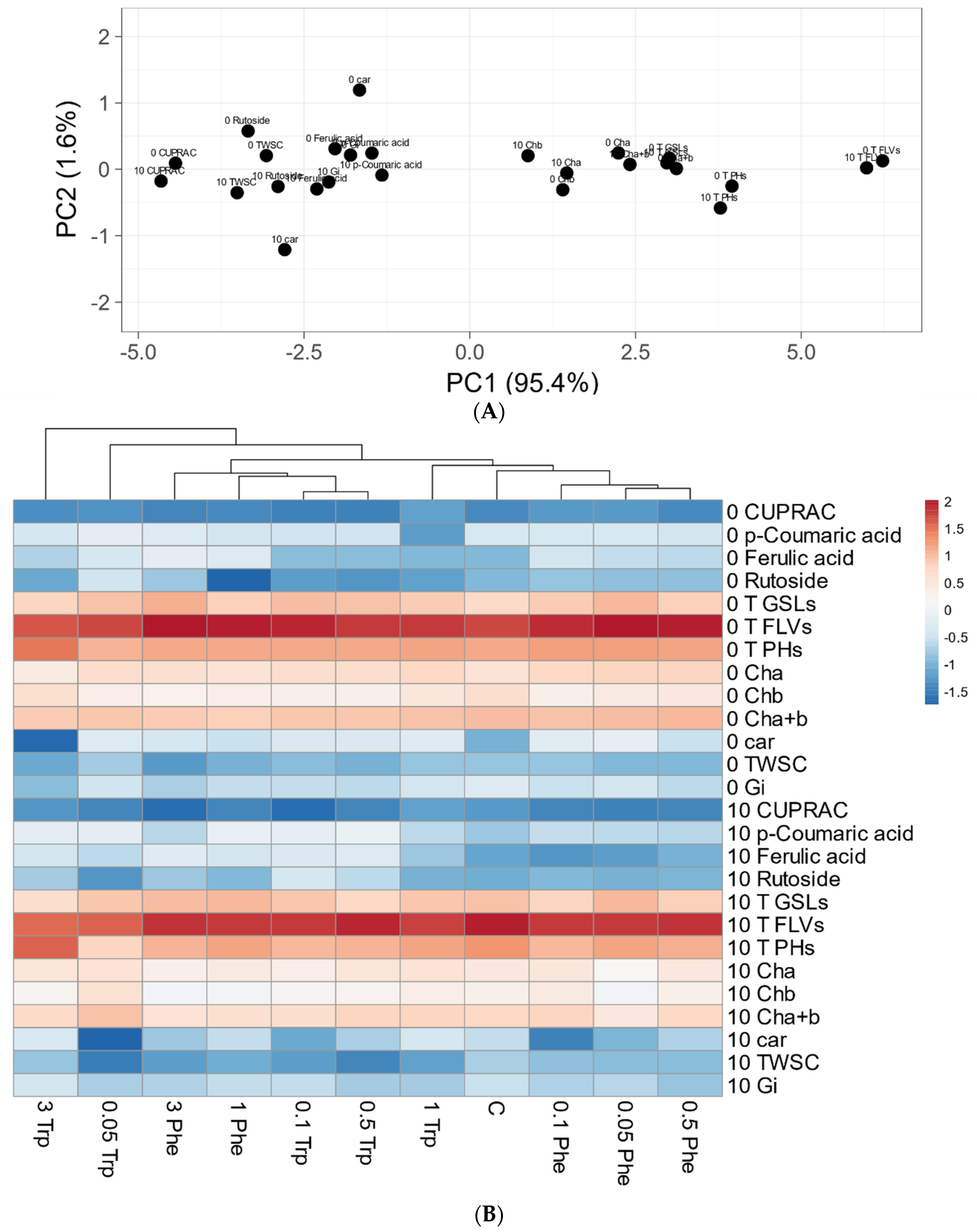
2.5. Principal Component Analysis (PCA)
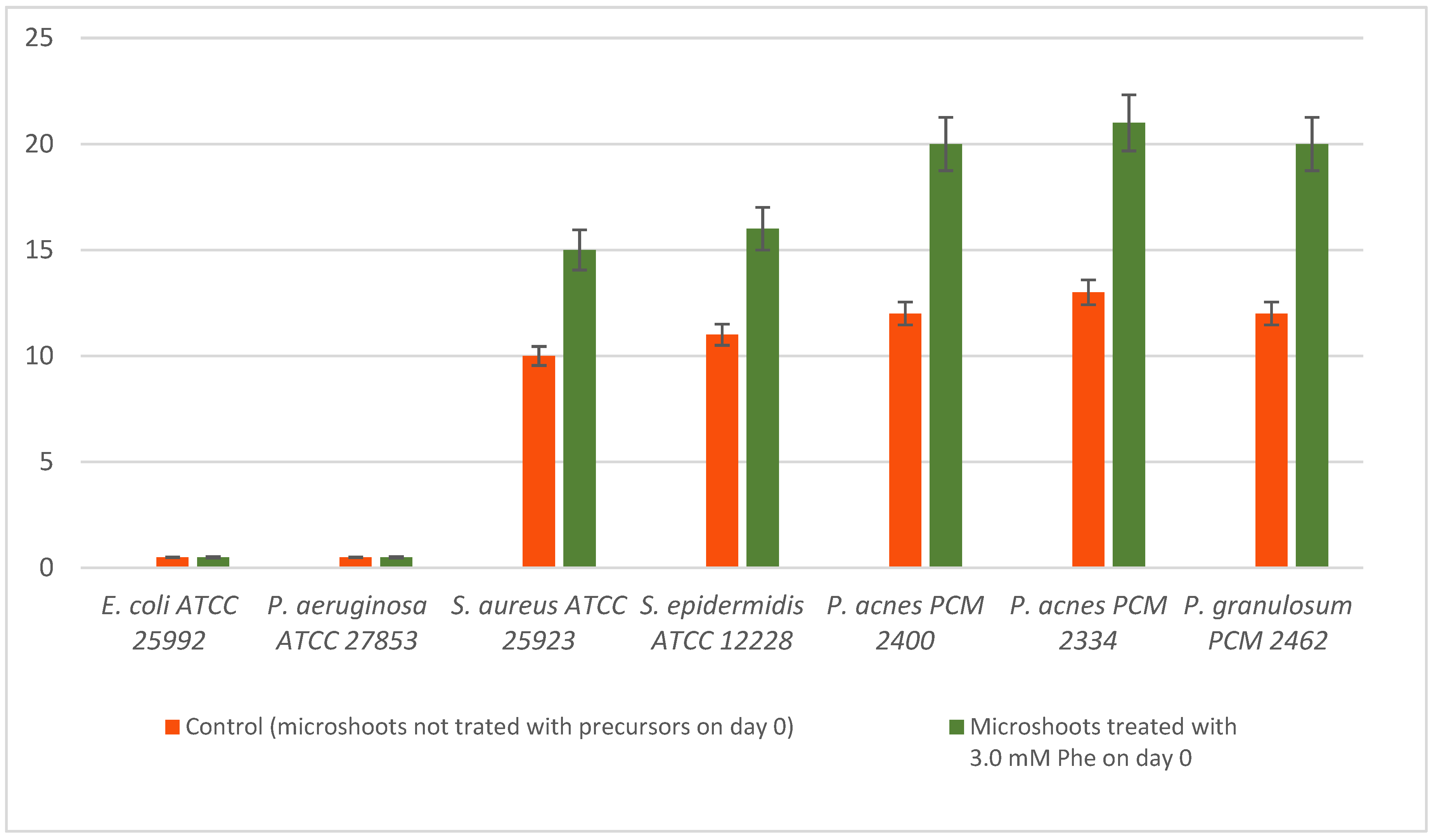
2.6. Antimicrobial Activity
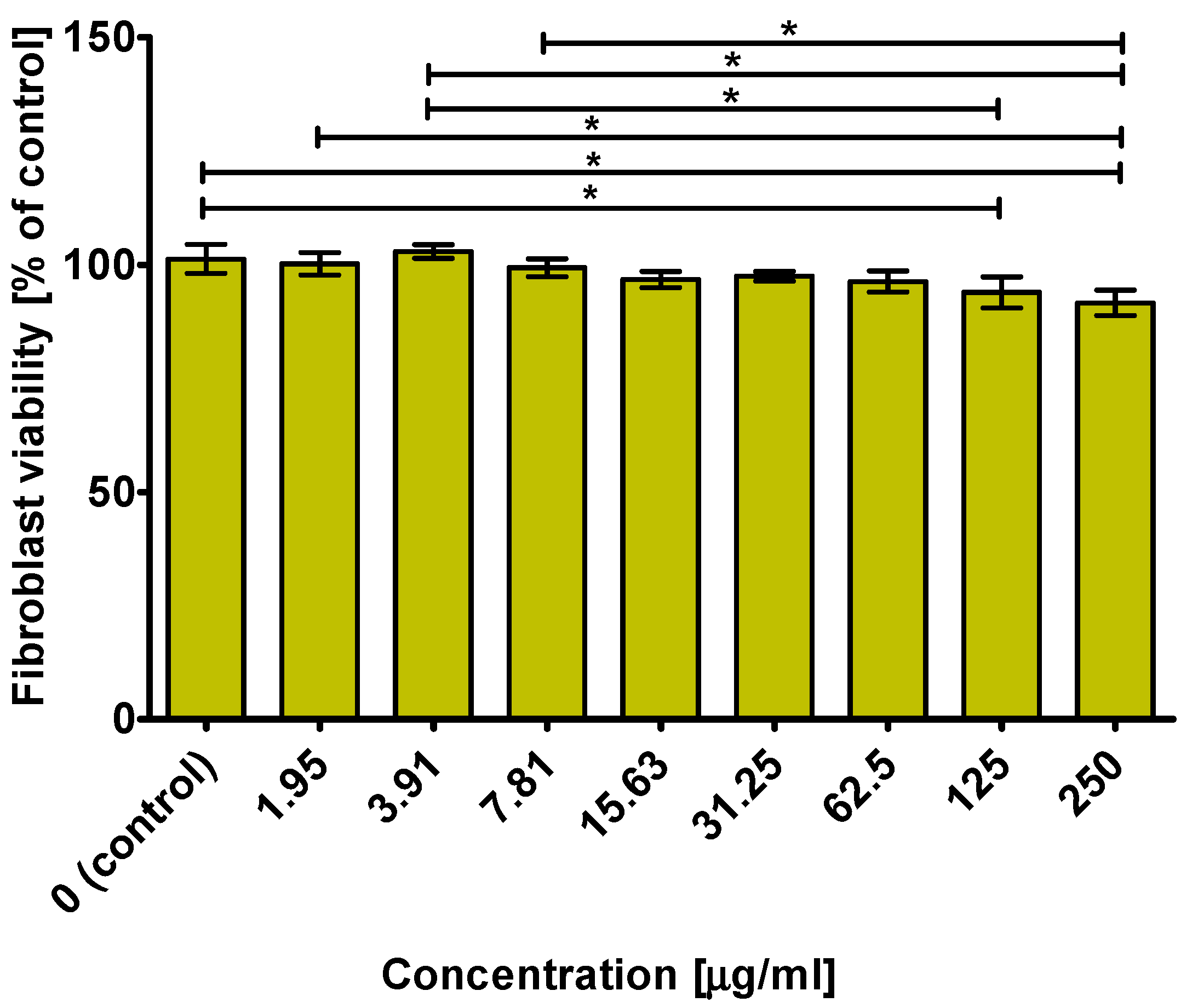
2.7. Cytotoxicity towards Normal Human Fibroblast Cells
3. Materials and Methods
3.1. Experimental In Vitro Cultures
3.2. Procedure for Precursor Feeding
3.3. Calculating the Growth Index
3.4. Biomass Extraction
3.5. Phytochemical Analyses of Metabolites
3.5.1. Determination of Total Soluble Saccharides
3.5.2. Analysis of Photosynthetic Pigments
3.5.3. Spectrophotometric Analysis of the Total GSL Pool
3.5.4. Total Flavonoid Assay
3.5.5. Total Phenolic Assay
3.5.6. Analysis of GSL Content with UHPLC-DAD-MS/MS
3.5.7. Analysis of Polyphenol Compounds Using HPLC-DAD
3.6. Antioxidant Activity Assays
3.6.1. CUPRAC Assay
3.6.2. FRAP Assay
3.6.3. DPPH Radical-Scavenging Activity Assay
3.7. In Vitro Antimicrobial Assays
3.8. Cytotoxicity Evaluation
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Klimek-Szczykutowicz, M.; Szopa, A.; Ekiert, H. Chemical composition, traditional and professional use in medicine, application in environmental protection, position in food and cosmetics industries, and biotechnological studies of Nasturtium officinale (watercress)—A review. Fitoterapia 2018, 129, 283–292. [Google Scholar] [CrossRef]
- De Lira, R.M.; Silva, Ê.F.D.F.E.; Da Silva, A.O.; De Medeiros, P.R.F.; Da Silva, G.F.; Soares, H.R.E. Watercress and chinese cabbage in a hydroponic system using groundwater. Rev. Caatinga 2019, 32, 1038–1047. [Google Scholar] [CrossRef]
- Lira, R.M.; Silva, Ê.F.F.; Silva, G.F.; Soares, H.R.; Willadino, L.G. Growth, water consumption and mineral composition of watercress under hydroponic system with brackish water. Hortic. Bras. 2018, 36, 13–19. [Google Scholar] [CrossRef]
- EFSA European Food Safety Authority (EFSA). Available online: http://www.efsa.europa.eu/ (accessed on 25 January 2020).
- Boligon, A.A.; Janovik, V.; Boligon, A.A.; Pivetta, C.R.; Pereira, R.P.; Da Rocha, J.B.T.; Athayde, M.L. HPLC analysis of polyphenolic compounds and antioxidant activity in Nasturtium officinale. Int. J. Food Prop. 2013, 16, 61–69. [Google Scholar] [CrossRef]
- Martínez-Sánchez, A.; Gil-Izquierdo, A.; Gil, M.I.; Ferreres, F. A comparative study of flavonoid compounds, vitamin C, and antioxidant properties of baby leaf Brassicaceae species. J. Agric. Food Chem. 2008, 56, 2330–2340. [Google Scholar] [CrossRef]
- Afsharypuor, S.; Salehi, M. Volatile constituents of leaves and stems of Nasturtium officinale R. Br. J. Essent. Oil Res. 2008, 20, 517–518. [Google Scholar] [CrossRef]
- Jeon, J.; Bong, S.J.; Park, J.S.; Park, Y.K.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. De novo transcriptome analysis and glucosinolate profiling in watercress (Nasturtium officinale R. Br.). BMC Genom. 2017, 18, 1–14. [Google Scholar] [CrossRef]
- Palaniswamy, U.R.; McAvoy, R.J. Watercress: A salad crop with chemopreventive potential. Horttechnology 2001, 11, 622–626. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.-M.; Stepanov, I.; Murphy, S.E.; Wang, R.; Allen, S.; Jensen, J.; Strayer, L.; Adams-Haduch, J.; Upadhyaya, P.; Le, C.; et al. Clinical trial of 2-phenethyl isothiocyanate as an inhibitor of metabolic activation of a tobacco-specific lung carcinogen in cigarette smokers. Cancer Prev. Res. 2016, 9, 396–405. [Google Scholar] [CrossRef] [Green Version]
- De Souza, D.A.; Costa, P.M.; Ribeiro, R.I.M.A.; Vidigal, P.V.T.; Pinto, F.C.H. Daily intake of watercress causes inhibition of experimental Ehrlich tumor growth. J. Bras. Patol. Med. Lab. 2016, 52, 393–399. [Google Scholar]
- Li, Q.; Zhan, M.; Chen, W.; Zhao, B.; Yang, K.; Yang, J.; Yi, J.; Huang, Q.; Mohan, M.; Hou, Z.; et al. Phenylethyl isothiocyanate reverses cisplatin resistance in biliary tract cancer cells via glutathionylation-dependent degradation of Mcl-1. Oncotarget 2016, 7, 10271–10282. [Google Scholar] [CrossRef] [Green Version]
- Bahramikia, S.; Yazdanparast, R. Antioxidant efficacy of Nasturtium officinale extracts using various in vitro assay systems. JAMS J. Acupunct. Meridian Stud. 2010, 3, 283–290. [Google Scholar] [CrossRef]
- Sadeghi, H.; Mostafazadeh, M.; Sadeghi, H.; Naderian, M.; Barmak, M.J.; Talebianpoor, M.S.; Mehraban, F. In vivo anti-inflammatory properties of aerial parts of Nasturtium officinale. Pharm. Biol. 2014, 52, 169–174. [Google Scholar] [CrossRef]
- Holst, B.; Williamson, G. A critical review of the bioavailability of glucosinolates and related compounds. Nat. Prod. Rep. 2004, 21, 425–447. [Google Scholar] [CrossRef]
- Karuppusamy, S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J. Med. Plants Res. 2009, 3, 1222–1239. [Google Scholar]
- Smetanska, I. Production of secondary metabolites using plant cell cultures. Adv. Biochem. Eng. Biotechnol. 2008, 111, 187–228. [Google Scholar]
- Steingroewer, J.; Bley, T.; Georgiev, V.; Ivanov, I.; Lenk, F.; Marchev, A.; Pavlov, A. Bioprocessing of differentiated plant in vitro systems. Eng. Life Sci. 2013, 13, 26–38. [Google Scholar] [CrossRef]
- Krol, A.; Kokotkiewicz, A.; Szopa, A.; Ekiert, H.; Luczkiewicz, M. Bioreactor-Grown Shoot Cultures for the Secondary Metabolite Production. In Plant Cell and Tissue Differentiation and Secondary Metabolites: Fundamentals and Applications; Ramawat, K.G., Ekiert, H.M., Goyal, S., Eds.; Springer Nature: Cham, Switzerland, 2021; ISBN 978-3-030-30184-2. [Google Scholar]
- Sankar-Thomas, Y.D.; Lieberei, R. Camptothecin accumulation in various organ cultures of Camptotheca acuminata Decne grown in different culture systems. Plant Cell Tissue Organ Cult. 2011, 106, 445–454. [Google Scholar] [CrossRef]
- Kokotkiewicz, A.; Bucinski, A.; Luczkiewicz, M. Xanthone, benzophenone and bioflavonoid accumulation in Cyclopia genistoides (L.) Vent. (honeybush) shoot cultures grown on membrane rafts and in a temporary immersion system. Plant Cell Tissue Organ Cult. 2015, 120, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Pujante, P.J.; Borja-Martínez, M.; Pedreño, M.Á.; Almagro, L. Biosynthesis and bioactivity of glucosinolates and their production in plant in vitro cultures. Planta 2017, 246, 19–32. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef]
- Klimek-Szczykutowicz, M.; Dziurka, M.; Blažević, I.; Đulović, A.; Granica, S.; Korona-Glowniak, I.; Ekiert, H.; Szopa, A. Phytochemical and biological activity studies on Nasturtium officinale (watercress) microshoot cultures grown in RITA® temporary immersion systems. Molecules 2020, 25, 5257. [Google Scholar] [CrossRef]
- Weremczuk-Jeżyna, I.; Lisiecki, P.; Gonciarz, W.; Kuźma, Ł.; Szemraj, M.; Chmiela, M.; Grzegorczyk-Karolak, I. Transformed shoots of Dracocephalum forrestii W.W. Smith from different bioreactor systems as a rich source of natural phenolic compounds. Molecules 2020, 25, 4533. [Google Scholar] [CrossRef]
- Klimek-Szczykutowicz, M.; Szopa, A.; Dziurka, M.; Komsta, Ł.; Tomczyk, M.; Ekiert, H. The influence of Nasturtium officinale R. Br. agar and agitated microshoot culture media on glucosinolate and phenolic acid production, and antioxidant activity. Biomolecules 2020, 10, 1216. [Google Scholar] [CrossRef]
- Szopa, A.; Klimek-Szczykutowicz, M.; Kokotkiewicz, A.; Maślanka, A.; Król, A.; Luczkiewicz, M.; Ekiert, H. Phytochemical and biotechnological studies on Schisandra chinensis cultivar Sadova No. 1—A high utility medicinal plant. Appl. Microbiol. Biotechnol. 2018, 102, 5105–5120. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, J.; Wang, X.D.; Zhao, B.; Wang, Y.C. Enhanced production of phenylethanoid glycosides by precursor feeding to cell culture of Cistanche deserticola. Process Biochem. 2005, 40, 3480–3484. [Google Scholar] [CrossRef]
- Chand Meena, M.; Kesh Meena, R.; Patni, V. Effect of elicitor on quercetin production in cell cultures of Citrullus colocynthis (Linn.) Schrad. Pharma Innov. 2014, 3, 18–23. [Google Scholar]
- Sajjalaguddam, R.R.; Paladugu, A. Phenylalanine enhances quercetin content in in vitro cultures of Abutilon indicum L. J. Appl. Pharm. Sci. 2015, 5, 80–84. [Google Scholar] [CrossRef] [Green Version]
- Wielanek, M.; Królicka, A.; Bergier, K.; Gajewska, E.; Skłodowska, M. Transformation of Nasturtium officinale, Barbarea verna and Arabis caucasica for hairy roots and glucosinolate-myrosinase system production. Biotechnol. Lett. 2009, 31, 917–921. [Google Scholar] [CrossRef]
- El-Hawary, S.S.; Abd El-Kader, E.M.; Rabeh, M.A.; Abdel Jaleel, G.A.; Arafat, M.A.; Schirmeister, T.; Abdelmohsen, U.R. Eliciting callus culture for production of hepatoprotective flavonoids and phenolics from Sequoia sempervirens (D. Don Endl). Nat. Prod. Res. 2020, 34, 3125–3129. [Google Scholar] [CrossRef]
- Koca, N.; Karaman, Ş. The effects of plant growth regulators and L-phenylalanine on phenolic compounds of sweet basil. Food Chem. 2015, 166, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Skrzypczak-Pietraszek, E.; Piska, K.; Pietraszek, J. Enhanced production of the pharmaceutically important polyphenolic compounds in Vitex agnus castus L. shoot cultures by precursor feeding strategy. Eng. Life Sci. 2018, 18, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Szopa, A.; Kubica, P.; Komsta, Ł.; Walkowicz-Bożek, A.; Ekiert, H. The effect of feeding culture media with biogenetic precursors on high production of depsides in agitated shoot cultures of black and red aronias. Plant Cell Tissue Organ Cult. 2020, 142, 379–399. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- O’Donnell, F.; Smyth, T.J.P.; Ramachandran, V.N.; Smyth, W.F. A study of the antimicrobial activity of selected synthetic and naturally occurring quinolines. Int. J. Antimicrob. Agents 2010, 35, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Andini, S.; Araya-Cloutier, C.; Lay, B.; Vreeke, G.; Hageman, J.; Vincken, J.P. QSAR-based physicochemical properties of isothiocyanate antimicrobials against gram-negative and gram-positive bacteria. LWT 2021, 144, 111222. [Google Scholar] [CrossRef]
- Klimek-Szczykutowicz, M.; Szopa, A.; Blicharska, E.; Dziurka, M.; Komsta, Ł.; Ekiert, H. Bioaccumulation of selected macro- and microelements and their impact on antioxidant properties and accumulation of glucosinolates and phenolic acids in in vitro cultures of Nasturtium officinale (watercress) microshoots. Food Chem. 2019, 300, 125184. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Grzegorczyk, I.; Wysokińska, H. Liquid shoot culture of Salvia officinalis L. for micropropagation and production of antioxidant compounds; effects of triacontanol. Acta Soc. Bot. Pol. 2008, 77, 99–104. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. A colorimetric method forthe determination of sugars. Nature 1951, 168, 167–168. [Google Scholar] [CrossRef]
- Bach, A.; Kapczyńska, A.; Dziurka, K.; Dziurka, M. Phenolic compounds and carbohydrates in relation to bulb formation in Lachenalia “Ronina” and “Rupert” in vitro cultures under different lighting environments. Sci. Hortic. 2015, 188, 23–29. [Google Scholar] [CrossRef]
- Czyczyło-Mysza, I.; Tyrka, M.; Marcińska, I.; Skrzypek, E.; Karbarz, M.; Dziurka, M.; Hura, T.; Dziurka, K.; Quarrie, S.A. Quantitative trait loci for leaf chlorophyll fluorescence parameters, chlorophyll and carotenoid contents in relation to biomass and yield in bread wheat and their chromosome deletion bin assignments. Mol. Breed. 2013, 32, 189–210. [Google Scholar] [CrossRef] [Green Version]
- Gallaher, C.M.; Gallaher, D.D.; Peterson, S. Development and validation of a spectrophotometric method for quantification of total glucosinolates in Cruciferous vegetables. J. Agric. Food Chem. 2012, 60, 1358–1362. [Google Scholar] [CrossRef] [PubMed]
- Ramos, R.T.M.; Bezerra, I.C.F.; Ferreira, M.R.A.; Soares, L.A.L. Spectrophotometric quantification of flavonoids in herbal material, crude extract, and fractions from leaves of Eugenia uniflora Linn. Pharmacogn. Res. 2017, 9, 253–260. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzym. 1999, 299, 152–178. [Google Scholar]
- Blažević, I.; Đulović, A.; Čikeš Čulić, V.; Burčul, F.; Ljubenkov, I.; Ruščić, M.; Generalić Mekinić, I. Bunias erucago L.: Glucosinolate profile and in vitro biological potential. Molecules 2019, 24, 741. [Google Scholar] [CrossRef] [Green Version]
- Grosser, K.; van Dam, N.M. A straightforward method for glucosinolate extraction and analysis with high-pressure liquid chromatography (HPLC). J. Vis. Exp. 2017, 2017, 55425. [Google Scholar] [CrossRef] [Green Version]
- Brown, P.D.; Tokuhisa, J.G.; Reichelt, M.; Gershenzon, J. Variation of glucosinolate accumulation among different organs and developmental stages of Arabidopsis thaliana. Phytochemistry 2003, 62, 471–481. [Google Scholar] [CrossRef]
- Wathelet, J.P.; Iori, R.; Leoni, O.; Quinsac, O.; Palmieri, S. Guidelines for glucosinolate analysis in green tissues used for biofumigation. Agroindustria 2004, 3, 257–266. [Google Scholar]
- Ellnain-Wojtaszek, M.; Zgórka, G. High-performance liquid chromatography and thin-layer chromatography of phenolic acids from Gingko biloba L. leaves collected within vegetative period. J. Liq. Chromatogr. Relat. Technol. 1999, 22, 1457–1471. [Google Scholar] [CrossRef]
- Sułkowska-Ziaja, K.; Maślanka, A.; Szewczyk, A.; Muszyńska, B. Physiologically active compounds in four species of genus Phellinus. Nat. Prod. Commun. 2017, 12, 363–366. [Google Scholar]
- Özyürek, M.; Güçlü, K.; Bektaşoğlu, B.; Apak, R. Spectrophotometric determination of ascorbic acid by the modified CUPRAC method with extractive separation of flavonoids–La(III) complexes. Anal. Chim. Acta 2007, 588, 88–95. [Google Scholar] [CrossRef]
- Biesaga-Kościelniak, J.; Dziurka, M.; Ostrowska, A.; Mirek, M.; Kościelniak, J.; Janeczko, A. Brassinosteroid improves content of antioxidants in seeds of selected Leguminous plants. Aust. J. Crop Sci. 2014, 8, 378–388. [Google Scholar]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measuer of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blios, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 26, 1199–1200. [Google Scholar] [CrossRef]
- Bauer, A.; Kirby, W.; Sherris, J.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing. Eighteenth International Supplement. In CLSI Document M7-MIC; CLSI: Wayne, UK, 2008. [Google Scholar]
- Pitucha, M.; Woś, M.; Miazga-Karska, M.; Klimek, K.; Mirosław, B.; Pachuta-Stec, A.; Gładysz, A.; Ginalska, G. Synthesis, antibacterial and antiproliferative potential of some new 1-pyridinecarbonyl-4-substituted thiosemicarbazide derivatives. Med. Chem. Res. 2016, 25, 1666–1677. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day of Supplementation | C | Precursor Concentrations (mM) | ||||
|---|---|---|---|---|---|---|
| 0.05 | 0.1 | 0.5 | 1.0 | 3.0 | ||
| Phe | ||||||
| 0 | 12.23 ab ± 1.12 | 11.46 ab ± 1.20 | 11.88 abcd ± 1.06 | 10.80 abcd ± 0.78 | 10.93 abcd ± 0.86 | 10.74 e ± 0.61 |
| 10 | 8.32 bcde ± 0.52 | 8.06 bcde ± 0.67 | 8.57 bcde ± 0.44 | 6.92 cde ± 0.06 | 10.63 abcd ± 0.89 | 11.48 abc ± 0.85 |
| Trp | ||||||
| 0 | 12.23 ab ± 1.12 | 11.92 ab ± 1.00 | 12.46 ab ± 0.81 | 11.68 ab ± 0.91 | 12.10 ab ± 1.01 | 5.47 e ± 0.61 |
| 10 | 8.32 bcde ± 0.52 | 7.71 bcde ± 0.55 | 12.35 ab ± 1.12 | 8.45 bcde ± 0.32 | 6.28 de ± 0.20 | 13.66 a ± 0.77 |
| Day of Supplementation | C | Precursor Concentration (mM) | ||||
|---|---|---|---|---|---|---|
| 0.05 | 0.1 | 0.5 | 1.0 | 3.0 | ||
| Phe | ||||||
| 0 | 4.65 c ± 0.45 | 4.55 c ± 0.22 | 6.84 a ± 1.05 | 5.82 ab ± 0.23 | 4.88 c ± 0.05 | 4.34 cd ± 0.01 |
| 10 | 5.57 abc ± 0.06 | 5.12 c ± 0.40 | 6.19 ab ± 0.01 | 6.01 ab ± 0.03 | 4.40 cd ± 0.10 | 4.60 c ± 0.20 |
| Trp | ||||||
| 0 | 4.65 c ± 0.45 | 6.92 a ± 1.07 | 6.65 a ± 0.44 | 5.49 abc ± 0.02 | 5.60 ab ± 1.04 | 3.49 d ± 0.35 |
| 10 | 5.57 abc ± 0.06 | 1.58 e ± 0.22 | 3.91 cd ± 0.12 | 2.44 e ± 0.14 | 2.65 e ± 0.11 | 6.41 a ± 0.33 |
| Photosynthetic Pigments | Day of Supplementation | C | Precursor Concentration (mM) | ||||
|---|---|---|---|---|---|---|---|
| 0.05 | 0.1 | 0.5 | 1.0 | 3.0 | |||
| Chlorophyll a | Phe | ||||||
| 0 | 64.36 fgh ± 21.92 | 116.61 ab ± 4.73 | 133.32 a ± 24.93 | 132.57 a ± 41.47 | 89.82 cdf ± 17.03 | 118.17 ab ± 2.85 | |
| 10 | 50.93 gh ± 6.52 | 35.49 h ± 5.05 | 80.14 dfg ± 7.58 | 72.42 fgh ± 5.46 | 64.82 fgh ± 1.39 | 64.24 fgh ± 9.04 | |
| Trp | |||||||
| 0 | 64.36 fgh ± 21.92 | 119.32 ab ± 11.81 | 114.48 ab ± 16.21 | 118.34 ab ± 15.81 | 140.18 a ± 20.45 | 88.09 cdf ± 19.31 | |
| 10 | 50.93 gh ± 6.52 | 98.58 bcd ± 5.30 | 59.63 fgh ± 3.67 | 88.44 cdf ± 2.53 | 98.18 bcd ± 1.14 | 109.01 abc ± 12.87 | |
| Chlorophyll b | Phe | ||||||
| 0 | 77.03 bcd ± 6.12 | 59.00 bcde ± 4.73 | 58.02 bcde ± 10.02 | 74.38 bcd ± 6.60 | 56.64 bcde ± 18.33 | 60.84 bcde ± 4.40 | |
| 10 | 38.49 de ± 5.21 | 29.15 ± 4.07 e | 71.15 bcd ± 8.23 | 53.75 cde ± 1.39 | 33.36 de ± 0.73 | 42.63 cde ± 6.36 | |
| Trp | |||||||
| 0 | 77.03 bcd ± 6.12 | 61.19 bcde ± 6.68 | 56.98 bcde ± 6.76 | 58.82 bcde ± 11.33 | 82.39 bc ± 2.34 | 148.24 a ± 5.55 | |
| 10 | 38.49 de ± 5.21 | 100.71 b ± 17.93 | 50.18 cde ± 3.50 | 53.29 cde ± 3.99 | 60.84 bcde ± 0.98 | 60.21 bcd ± 7.09 | |
| Chlorophyll a + b | Phe | ||||||
| 0 | 141.39 cdef ± 8.31 | 175.61 abcde ± 9.45 | 191.28 abcd ± 34.87 | 207.01 abc ± 48.15 | 146.46 cdef ± 35.36 | 179.01 abcde ± 1.55 | |
| 10 | 89.42 ef ± 11.73 | 64.70 f ± 9.04 | 151.24 bcde ± 0.57 | 126.18 def ± 6.84 | 98.23 ef ± 0.57 | 106.88 def ± 15.40 | |
| Trp | |||||||
| 0 | 141.39 cdef ± 8.31 | 180.51 abcd ± 18.50 | 171.46 abcde ± 22.98 | 177.17 abcde ± 27.13 | 222.62 ab ± 43.84 | 236.39 a ± 74.88 | |
| 10 | 89.42 ef ± 11.73 | 199.29 abc ± 12.63 | 109.70 def ± 0.16 | 141.67 bcde ± 6.60 | 159.02 bcde ± 2.12 | 169.16 abcde ± 19.88 | |
| Carotenoids | Phe | ||||||
| 0 | 3.23 de ± 0.01 | 20.80 ab ± 0.24 | 21.72 a ± 0.90 | 12.27 c ± 1.71 | 11.75 c ± 0.33 | 19.01 ab ± 2.44 | |
| 10 | 7.37 d ± 0.98 | 4.32 de ± 0.73 | 1.96 e ± 0.27 | 9.22 cd ± 1.75 | 10.72 cd ± 0.65 | 9.33 cd ± 1.14 | |
| Trp | |||||||
| 0 | 3.23 de ± 0.01 | 17.75 ab ± 1.47 | 18.21 ab ± 3.26 | 18.67 ab ± 1.79 | 17.80 ab ± 3.01 | nd | |
| 10 | 7.37 d ± 0.98 | nd | 4.55 de ± 0.20 | 9.56 cd ± 1.14 | 12.62 c ± 0.08 | 16.94 b ± 2.28 | |
| Method | Day of Supplementation | C | Precursor Concentration (mM) | ||||
|---|---|---|---|---|---|---|---|
| 0.05 | 0.1 | 0.5 | 1.0 | 3.0 | |||
| Total GSLs content | Phe | ||||||
| 0 | 81.58 c ± 2.18 | 181.10 abc ± 11.50 | 163.51 abc ± 20.78 | 141.06 bc ± 10.65 | 147.48 bc ± 20.68 | 269.20 a ± 15.78 | |
| 10 | 125.04 bc ± 5.05 | 180.07 abc ± 15.73 | 142.89 bc ± 17.63 | 141.40 bc ± 15.66 | 196.73 ab ± 20.60 | 216.82 ab ± 18.87 | |
| Trp | |||||||
| 0 | 81.58 c ± 2.18 | 200.44 ab ± 12.04 | 194.89 ab ± 23.76 | 187.84 ± 27.11 | 184.27 ab ± 28.43 | 196.26 ab ± 12.56 | |
| 10 | 125.04 bc ± 5.05 | 190.72 ab ± 20.56 | 168.46 abc ± 28.34 | 140.09 abc ± 20.40 | 200.97 ab ± 5.17 | 161.70 abc ± 10.91 | |
| Total flavono-ids content | Phe | ||||||
| 0 | 565.16 g ± 14.32 | 1124.60 bcd ± 102.47 | 1211.09 bc ± 24.31 | 1188.98 bcd ± 39.50 | 1118.99 bcd ± 51.65 | 1364.38 a ± 80.14 | |
| 10 | 863.71 fe ± 49.96 | 808.38 fe ± 45.35 | 1043.81 cdf ± 5.68 | 944.88 dfe ± 9.07 | 878.70 fe ± 21.79 | 1016.75 cdf ± 23.76 | |
| Trp | |||||||
| 0 | 565.16 g ± 14.32 | 965.82 dfe ± 11.02 | 1032.24 cdf ± 91.49 | 958.72 dfe ± 16.55 | 1324.14 ab ± 123.67 | 1241.89 abc ± 74.62 | |
| 10 | 863.71 fe ± 49.96 | 739.22 f ± 55.75 | 825.89 fe ± 33.91 | 1131.38 bcd ± 120.72 | 1169.84 bcd ± 79.37 | 964.93 dfe ± 142.40 | |
| Total poly-phenols content | Phe | ||||||
| 0 | 189.61 i ± 25.82 | 251.37 defg ± 15.50 | 325.55 c ± 7.99 | 266.47 def ± 9.46 | 253.55 defg ± 9.90 | 282.68 d ± 7.75 | |
| 10 | 248.02 efg ± 4.55 | 235.10 fgh ± 8.99 | 228.29 gh ± 9.34 | 237.24 fg ± 0.05 | 274.53 de ± 3.98 | 244.28 efg ± 9.20 | |
| Trp | |||||||
| 0 | 189.61 i ± 25.82 | 258.81 dfg ± 5.05 | 258.59 dfg ± 10.01 | 271.26 de ± 15.39 | 349.34 c ± 15.56 | 873.11 b ± 40.89 | |
| 10 | 248.02 efg ± 4.55 | 151.43 j ± 14.05 | 201.89 hi ± 7.90 | 243.18 efg ± 10.06 | 335.12 c ± 7.94 | 1062.76 a ± 28.77 | |
| Subgroups of GSLs | No. * | GSLs (Trivial Name) | tR (min) | [M + Na]+ | C | Precursor Treatments | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 3.0 mM Phe, Day 0 | 3.0 mM Trp, Day 0 | |||||||||
| GSLs mg/100 g DW ± SD | Productivity | GSLs mg/100 g DW ± SD | Productivity | GSLs mg/100 g DW ± SD | Productivity | |||||
| Methionine derived | 1 | 7-(Methylsulfinyl)heptyl GSL | 6.55 | 422 | tr | nd | tr | nd | nd | nd |
| 2 | 8-(Methylsulfinyl)octyl GSL (Glucohirsutin) | 7.58 | 436 | tr | nd | tr | nd | nd | nd | |
| Phenylalanine derived | 3 | 2-Phenylethyl GSL (Gluconasturtiin) | 8.20 | 366 | 15.65 ± 1.49 | 35.77 | 36.71 ± 3.32 | 97.59 | 8.83 ± 0.89 | 13.14 |
| Tryptophan derived | 4 | 4-Hydroxyindol-3-ylmethyl GSL (Hydroxyglucobrassicin) | 5.85 | 407 | nd | nd | tr | nd | tr | nd |
| 5 | Indol-3-ylmethyl GSL (Glucobrassicin) | 7.64 | 391 | tr | nd | 6.78 ± 0.04 | 18.03 | 3.78 ± 0.24 | 5.63 | |
| 6 | 4-Methoxyindol-3-ylmethyl GSL (4-Methoxyglucobrassicin) | 8.35 | 421 | 12.79 ± 1.03 | 29.23 | 149.99 ± 17.44 | 398.77 | 76.12 ± 3.12 | 113.31 | |
| Polyphenol Compound | Day of Supplementation | C | Precursor Concentrations (mM) | ||||
|---|---|---|---|---|---|---|---|
| 0.05 | 0.1 | 0.5 | 1.0 | 3.0 | |||
| p-Coumaric acid | Phe | ||||||
| 0 | 10.99 c ± 2.07 | 13.54 c ± 0.82 | 16.27 bc ± 1.32 | 13.68 c ± 2.58 | 13.71 c ± 3.32 | 23.38 ab ± 3.55 | |
| 10 | 4.90 e ± 0.35 | 8.74 d ± 0.02 | 11.39 c ± 1.01 | 10.20 c ± 1.06 | 25.38 a ± 2.11 | 12.19 c ± 1.34 | |
| Trp | |||||||
| 0 | 10.99 c ± 2.07 | 22.57 b ± 3.35 | 13.79 c ± 0.09 | 14.05 c ± 1.22 | 2.51 e ± 0.20 | 15.09 bc ± 0.33 | |
| 10 | 4.90 e ± 0.35 | 21.54 b ± 0.42 | 24.65 a ± 2.64 | 29.11 a ± 3.70 | 8.97d ± 0.92 | 24.97 a ± 5.87 | |
| Ferulic acid | Phe | ||||||
| 0 | 3.66 d ± 0.70 | 9.58 cd ± 0.27 | 13.64 c ± 0.82 | 10.84 cd ± 2.44 | 18.20 b ± 3.20 | 27.76 a ± 2.12 | |
| 10 | 2.62 e ± 0.35 | 2.90 e ± 0.26 | 2.87 e ± 0.11 | 4.98 d ± 0.07 | 13.04 c ± 0.62 | 24.30 a ± 0.79 | |
| Trp | |||||||
| 0 | 3.66 d ± 0.70 | 13.70 c ± 2.07 | 6.45 d ± 0.11 | 6.34 d ± 0.59 | 4.35d ± 0.08 | 8.47 cd ± 0.88 | |
| 10 | 2.62 e ± 0.35 | 9.75 cd ± 0.30 | 18.12 b ± 1.43 | 19.64 b ± 1.91 | 6.07 d ± 0.54 | 13.81 c ± 1.98 | |
| Rutoside | Phe | ||||||
| 0 | 3.82 d ± 0.60 | 5.18 cd ± 0.36 | 6.71 b ± 0.90 | 6.57 c ± 0.36 | 1.26 e ± 0.32 | 9.50 b ± 1.14 | |
| 10 | 2.94 d ± 0.52 | 4.02 d ± 0.02 | 5.23 cd ± 0.06 | 5.25 cd ± 0.13 | 5.30 cd ± 0.09 | 9.94 b ± 0.89 | |
| Trp | |||||||
| 0 | 3.82 d ± 0.60 | 11.66 b ± 1.44 | 3.89 d ± 0.07 | 3.34 d ± 0.59 | 2.81 d ± 0.04 | 3.51 d ± 0.67 | |
| 10 | 2.94 d ± 0.52 | 2.48 d ± 0.12 | 16.03 a ± 1.28 | 11.64 b ± 1.23 | 3.89 d ± 0.21 | 7.55 c ± 0.66 | |
| Assay | Day of Supplementation | C | Precursor Concentrations (mM) | ||||
|---|---|---|---|---|---|---|---|
| 0.05 | 0.1 | 0.5 | 1.0 | 3.0 | |||
| CUPRAC | Phe | ||||||
| 0 | 1.54 j ± 0.06 | 2.61 b ± 0.14 | 3.05 a ± 0.18 | 2.51 bc ± 0.02 | 2.32 defg ± 0.07 | 3.02 a ± 0.01 | |
| 10 | 2.15 gh ± 0.01 | 1.71 j ± 0.04 | 2.18 fgh ± 0.01 | 2.27 defg ± 0.02 | 2.22 efgh ± 0.11 | 2.14 gh ± 0.03 | |
| Trp | |||||||
| 0 | 1.54 j ± 0.06 | 2.39 cde ± 0.02 | 2.34 def ± 0.01 | 2.37 de ± 0.01 | 2.70 b ± 0.09 | 2.09 h ± 0.02 | |
| 10 | 2.15 gh ± 0.01 | 1.90 i ± 0.16 | 1.70 j ± 0.07 | 2.45 cd ± 0.04 | 2.72 b ± 0.06 | 2.34 def ± 0.03 | |
| DPPH | Phe | ||||||
| 0 | 0.47j ± 0.02 | 0.66 dfgh ± 0.04 | 0.86 ab ± 0.05 | 0.77 bcd ± 0.05 | 0.77 bcd ± 0.02 | 0.85 ab ± 0.03 | |
| 10 | 0.69 dfg ± 0.09 | 0.73 bcd ± 0.06 | 0.72 cdf ± 0.07 | 0.57 hij ± 0.02 | 0.82 abc ± 0.01 | 0.64 efghi ± 0.06 | |
| Trp | |||||||
| 0 | 0.47j ± 0.02 | 0.60 ghi ± 0.04 | 0.54 ij ± 0.02 | 0.64 efghi ± 0.01 | 0.62 fghi ± 0.09 | 0.69 dfg ± 0.09 | |
| 10 | 0.69 dfg ± 0.09 | 0.77 bcd ± 0.06 | 0.54 ij ± 0.03 | 0.90 a ± 0.03 | 0.62 fghi ± 0.01 | 0.61 fghi ± 0.03 | |
| FRAP | Phe | ||||||
| 0 | 0.24 i ± 0.01 | 0.61 bc ± 0.02 | 0.94 a ± 0.26 | 0.55 cde ± 0.01 | 0.57 cd ± 0.01 | 0.73 b ± 0.03 | |
| 10 | 0.40 fgh ± 0.02 | 0.32 hi ± 0.05 | 0.40 gh ± 0.04 | 0.39 gh ± 0.01 | 0.52 cdfg ± 0.01 | 0.31 hi ± 0.11 | |
| Trp | |||||||
| 0 | 0.24 i ± 0.01 | 0.52 cdfg ± 0.01 | 0.49 cdfg ± 0.01 | 0.52 cdfg ± 0.01 | 0.53 cdf ± 0.02 | 0.42 fgh ± 0.01 | |
| 10 | 0.40 fgh ± 0.02 | 0.33 hi ± 0.02 | 0.34 hi ± 0.02 | 0.43 fgh ± 0.01 | 0.49 cdfg ± 0.03 | 0.44 dfgh ± 0.01 | |
| N. officinale Extract | S. aureus ATCC 25923 | S. epidermidis ATCC 12228 | P. acnes PCM 2400 | P. acnes PCM 2334 | P. granulosum PCM 2462 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| MIC | MIC | MIC | MIC | MIC | ||||||
| C | 2000 | >8 | 2000 | >8 | 1000 | >8 | 1000 | >8 | 500 | 16 |
| Phe | 1000 | >8 | 1000 | 8 | 500 | 8 | 250 | 8 | 500 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klimek-Szczykutowicz, M.; Dziurka, M.; Blažević, I.; Đulović, A.; Miazga-Karska, M.; Klimek, K.; Ekiert, H.; Szopa, A. Precursor-Boosted Production of Metabolites in Nasturtium officinale Microshoots Grown in Plantform Bioreactors, and Antioxidant and Antimicrobial Activities of Biomass Extracts. Molecules 2021, 26, 4660. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26154660
Klimek-Szczykutowicz M, Dziurka M, Blažević I, Đulović A, Miazga-Karska M, Klimek K, Ekiert H, Szopa A. Precursor-Boosted Production of Metabolites in Nasturtium officinale Microshoots Grown in Plantform Bioreactors, and Antioxidant and Antimicrobial Activities of Biomass Extracts. Molecules. 2021; 26(15):4660. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26154660
Chicago/Turabian StyleKlimek-Szczykutowicz, Marta, Michał Dziurka, Ivica Blažević, Azra Đulović, Małgorzata Miazga-Karska, Katarzyna Klimek, Halina Ekiert, and Agnieszka Szopa. 2021. "Precursor-Boosted Production of Metabolites in Nasturtium officinale Microshoots Grown in Plantform Bioreactors, and Antioxidant and Antimicrobial Activities of Biomass Extracts" Molecules 26, no. 15: 4660. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26154660