
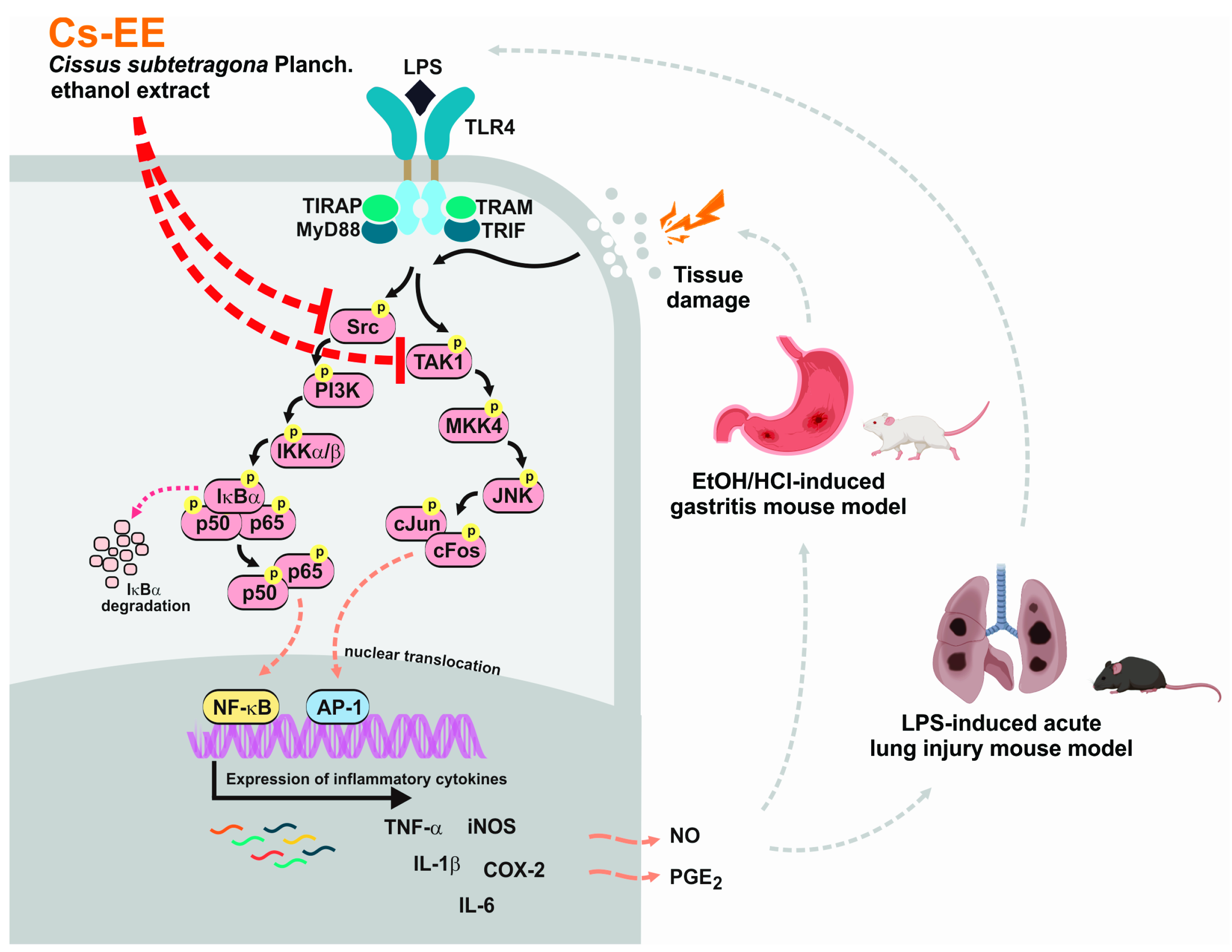
Cissus subtetragona Planch. Ameliorates Inflammatory Responses in LPS-induced Macrophages, HCl/EtOH-induced Gastritis, and LPS-induced Lung Injury via Attenuation of Src and TAK1

 , , ,
, , ,  , ,
, , 
Abstract
:1. Introduction
2. Results
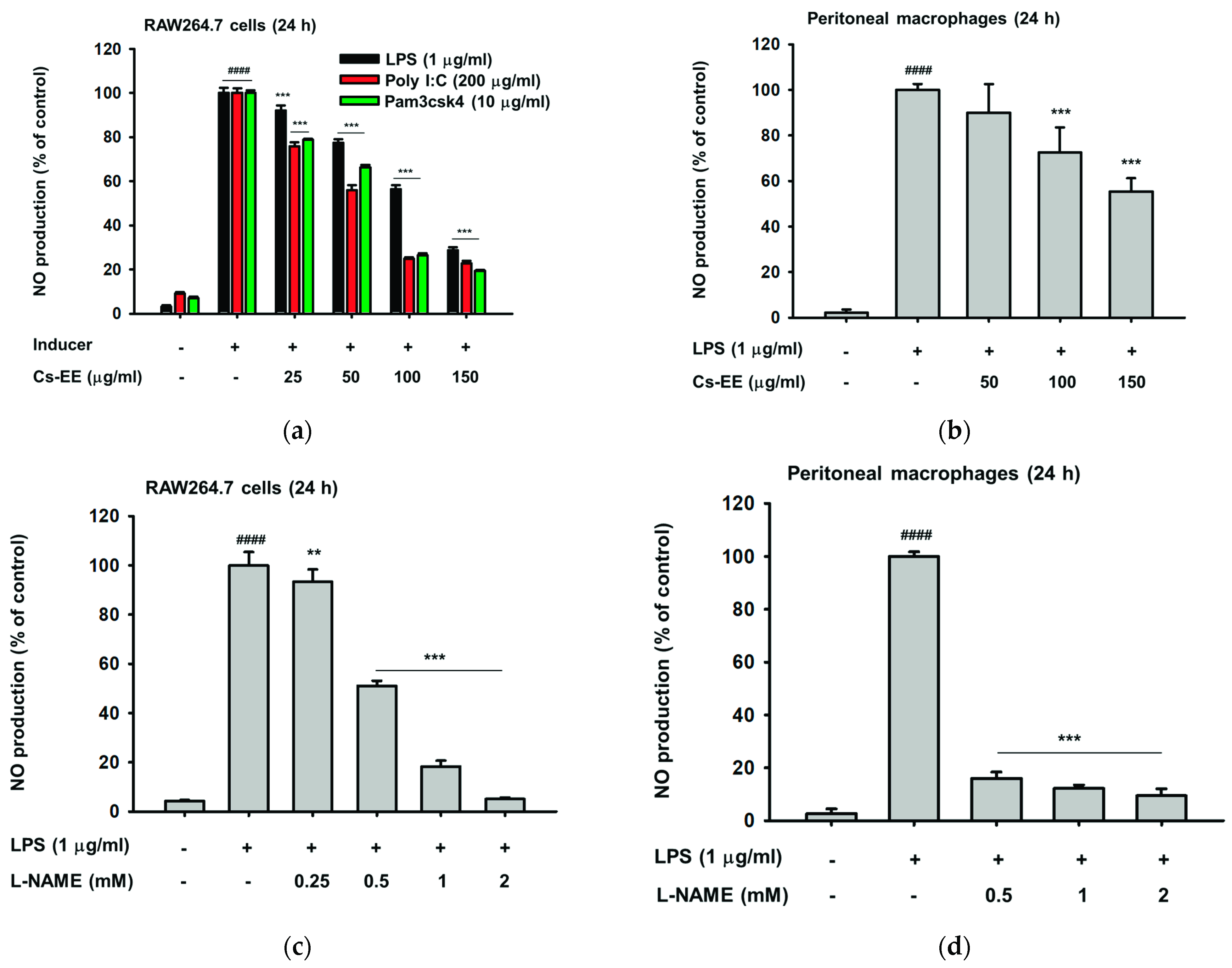
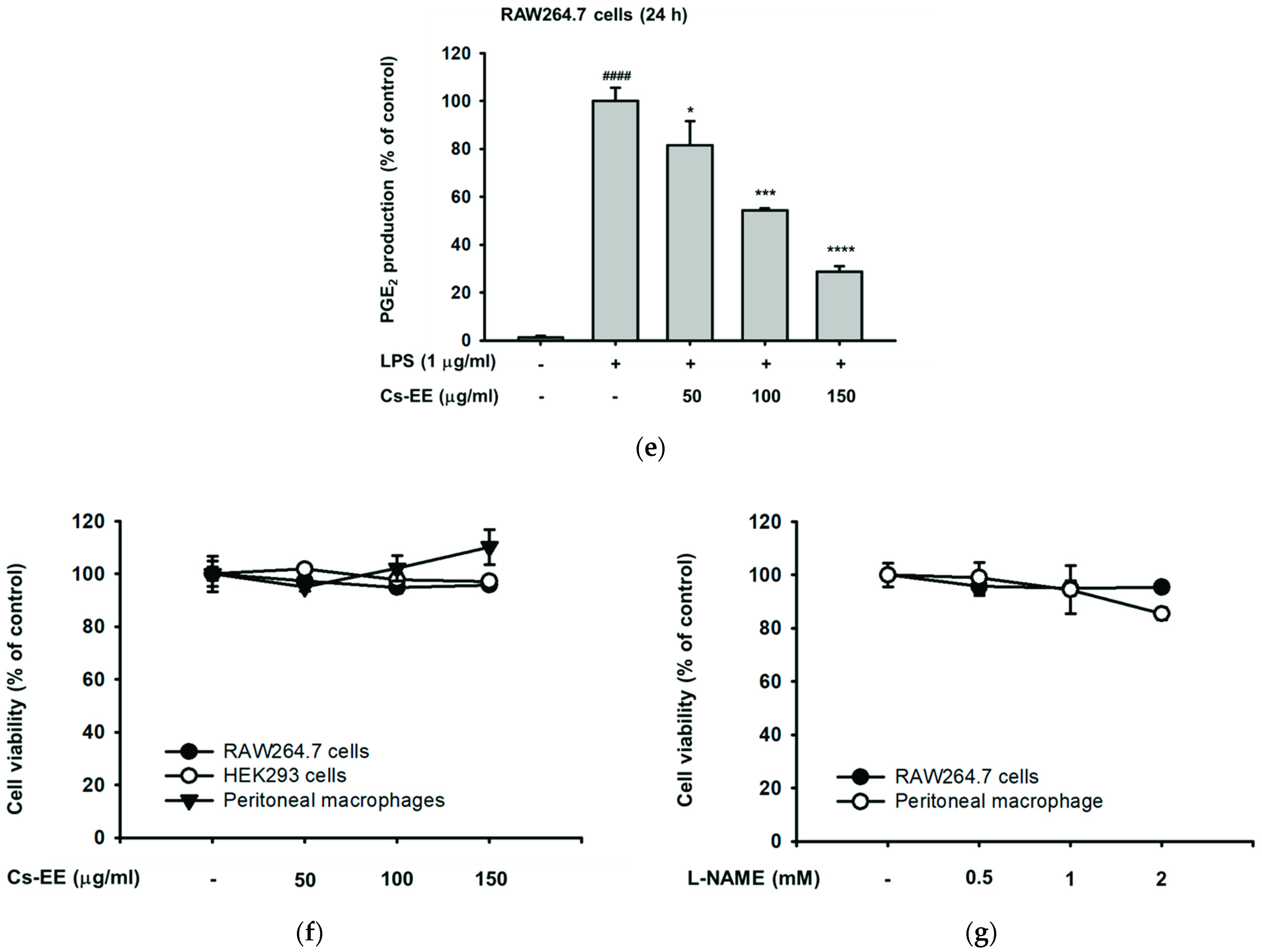
2.1. Cs-EE Affects NO and PGE2 Production without Showing Any Cytotoxicity
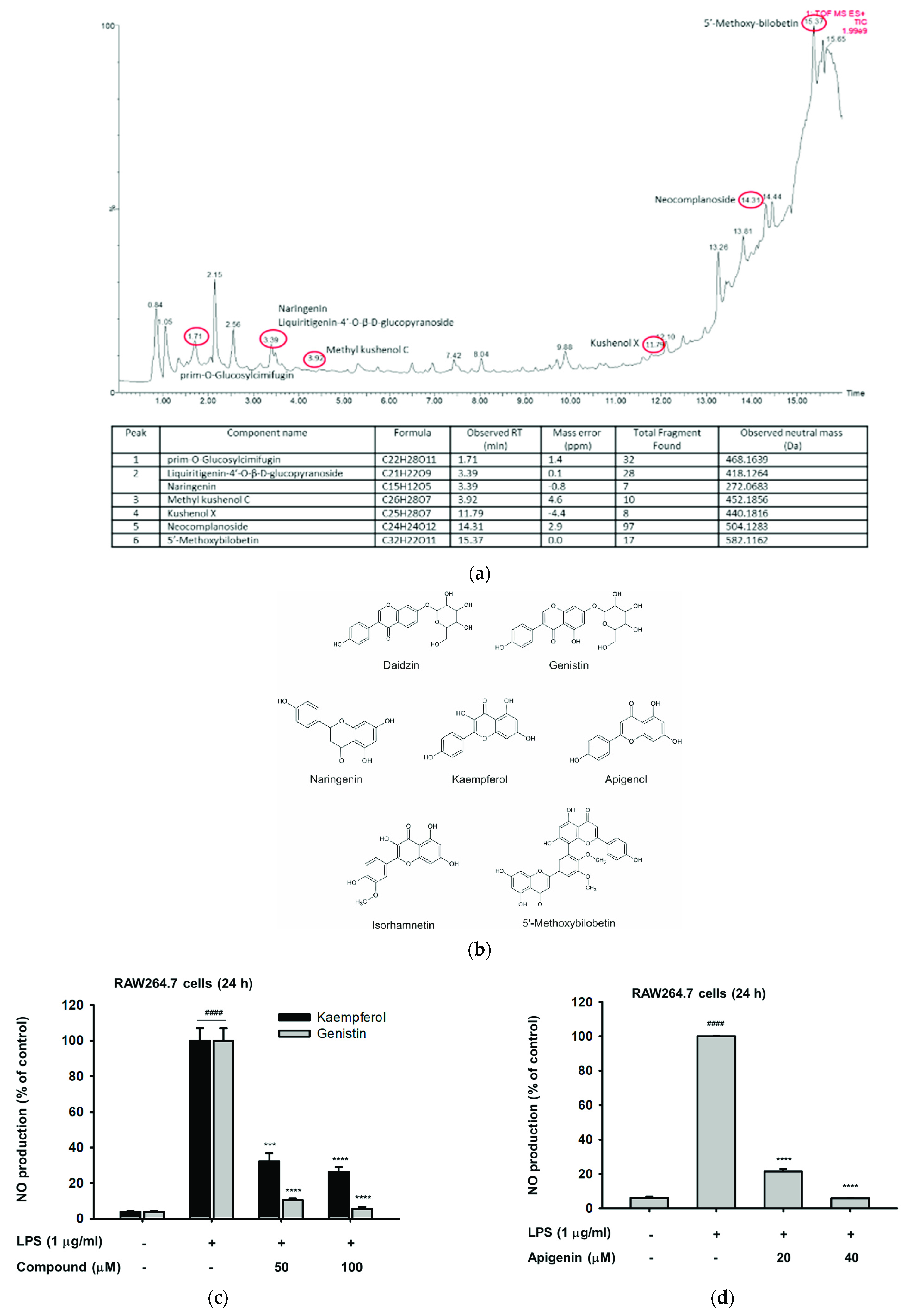
2.2. The Phytochemical Constituents of Cs-EE
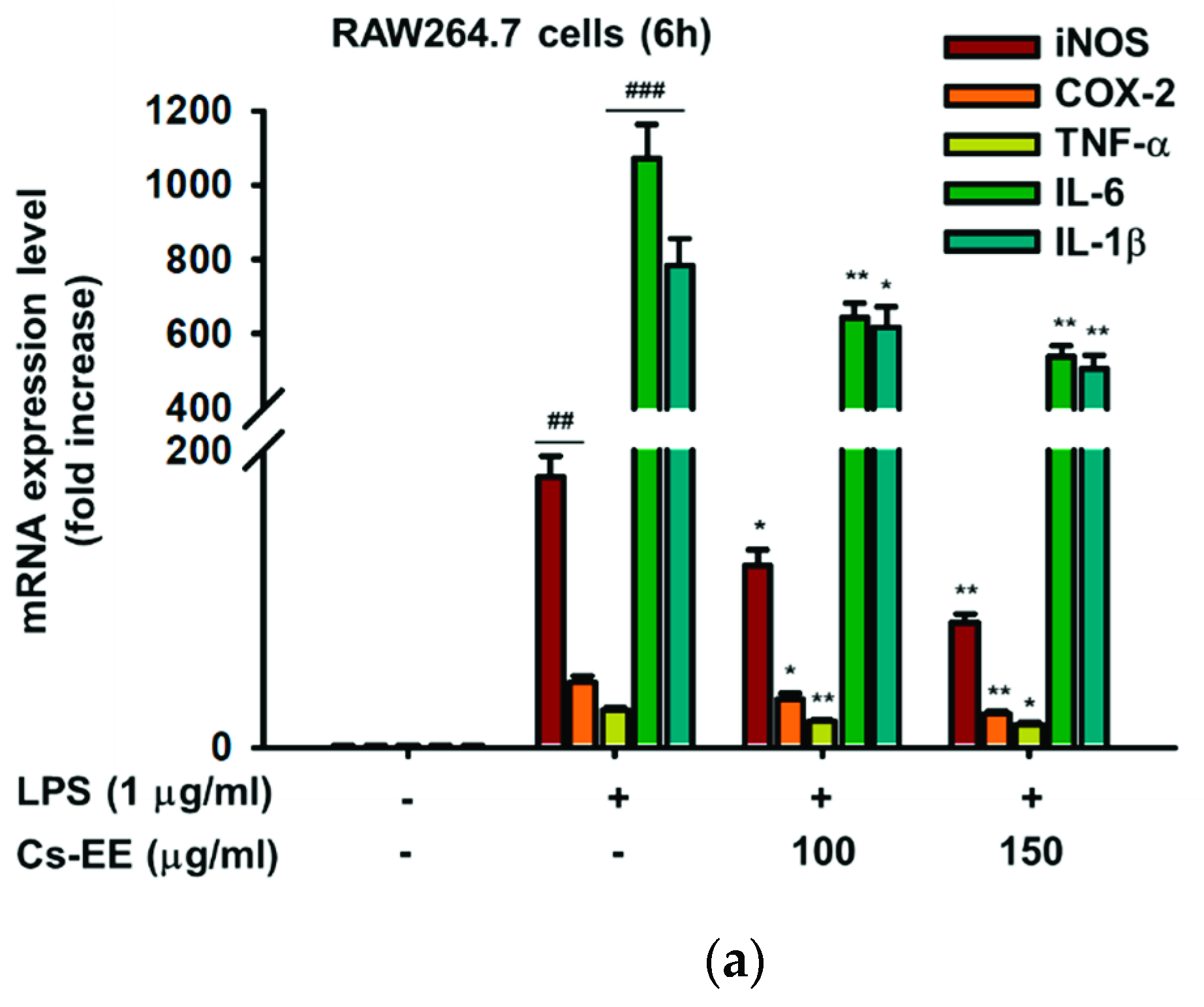
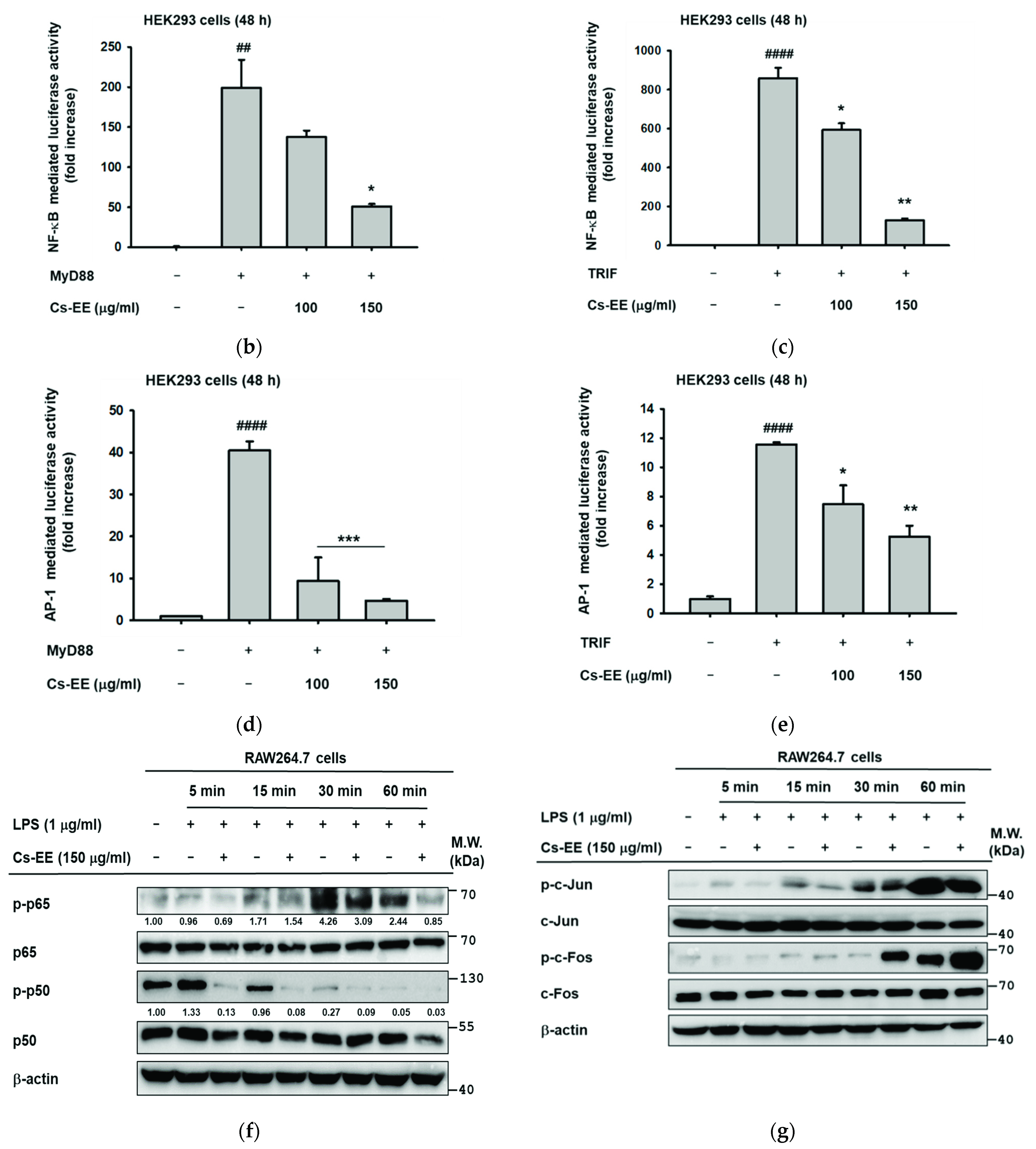
2.3. Effects of Cs-EE on the Expression of Inflammatory Genes and Transcriptional Activation of NF-κB and AP-1
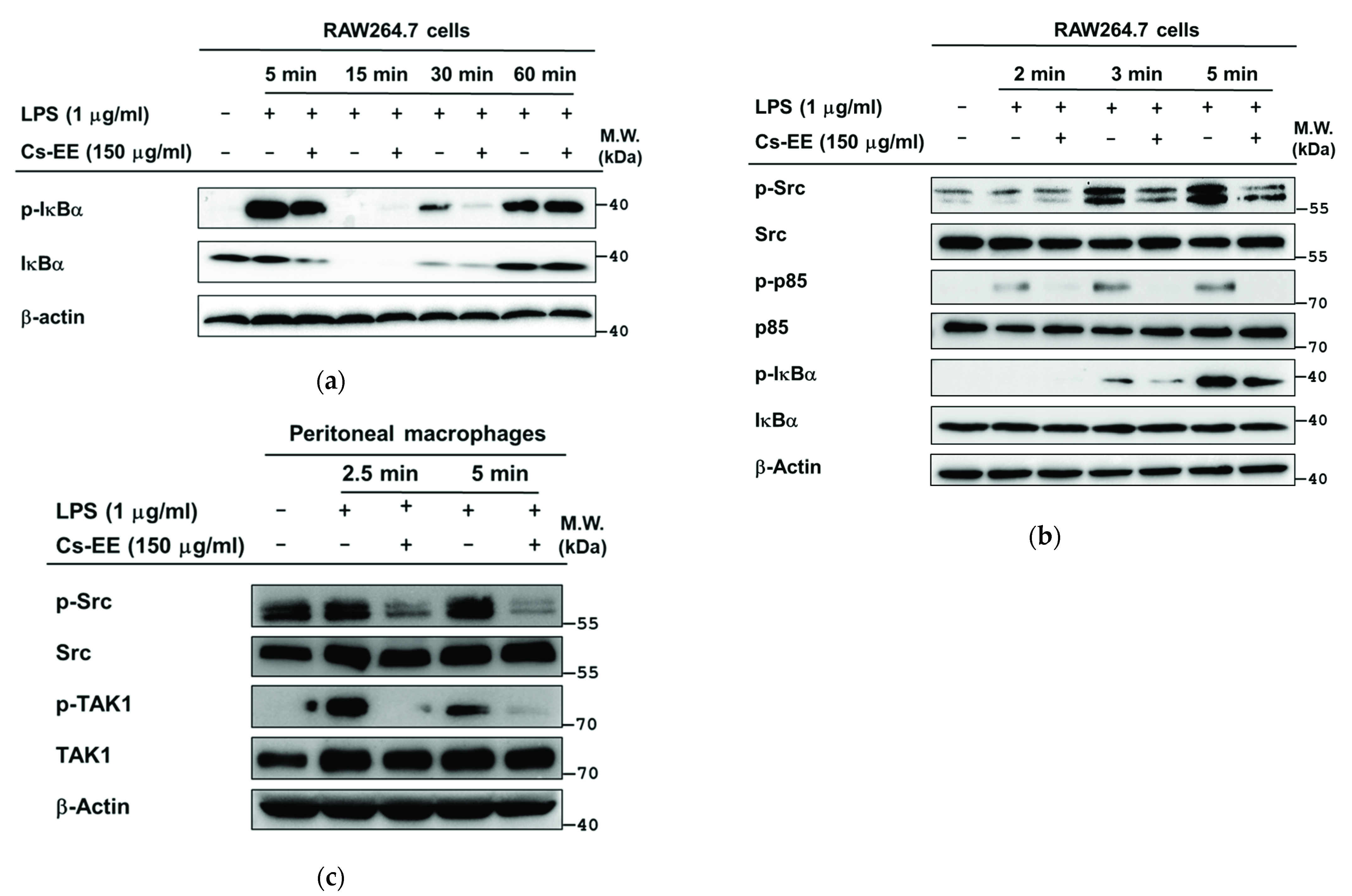
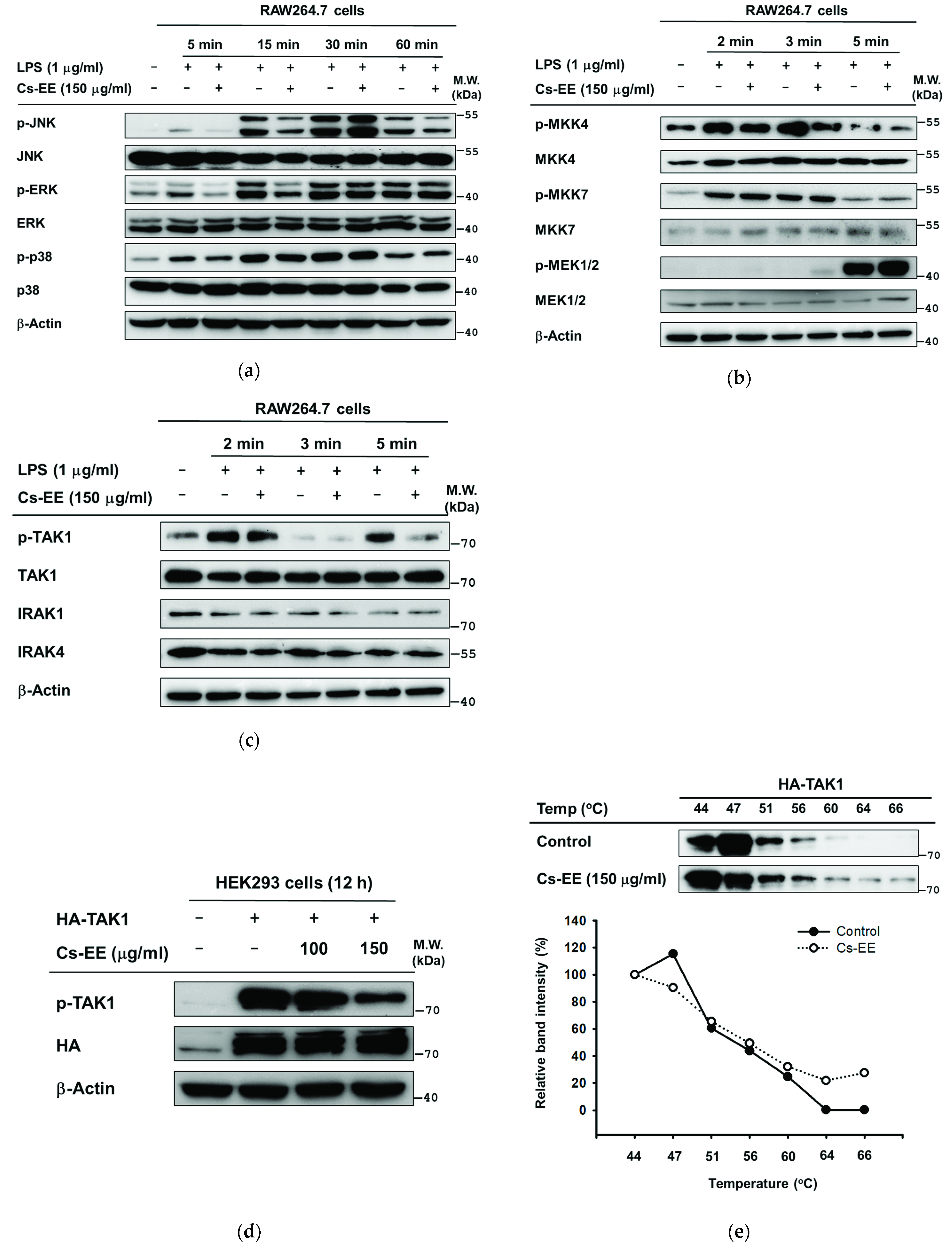
2.4. Effects of Cs-EE on the Signaling Events Upstream of NF-κB Activation
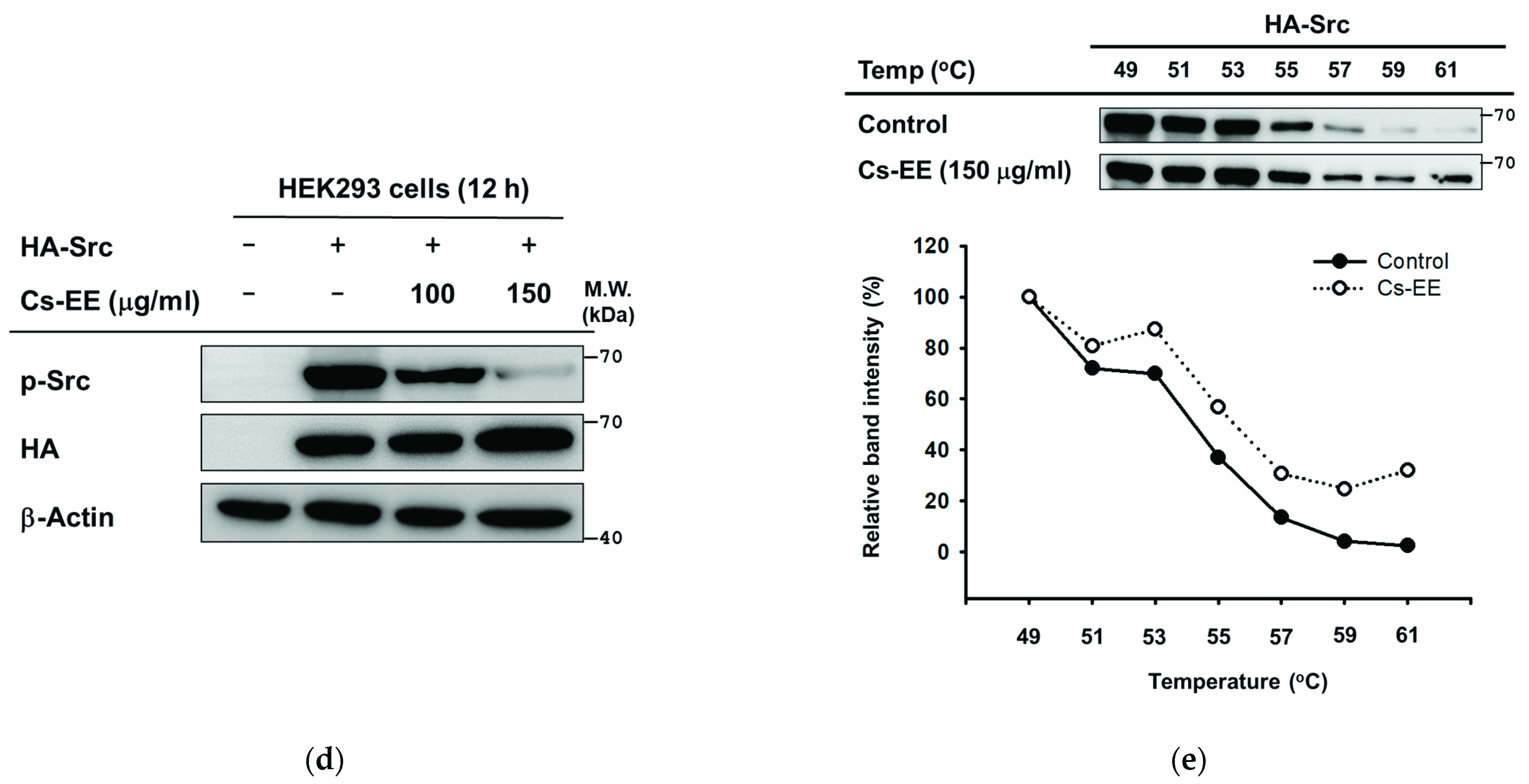
2.5. Effects of Cs-EE on the Signaling Events Upstream of AP-1 Activation
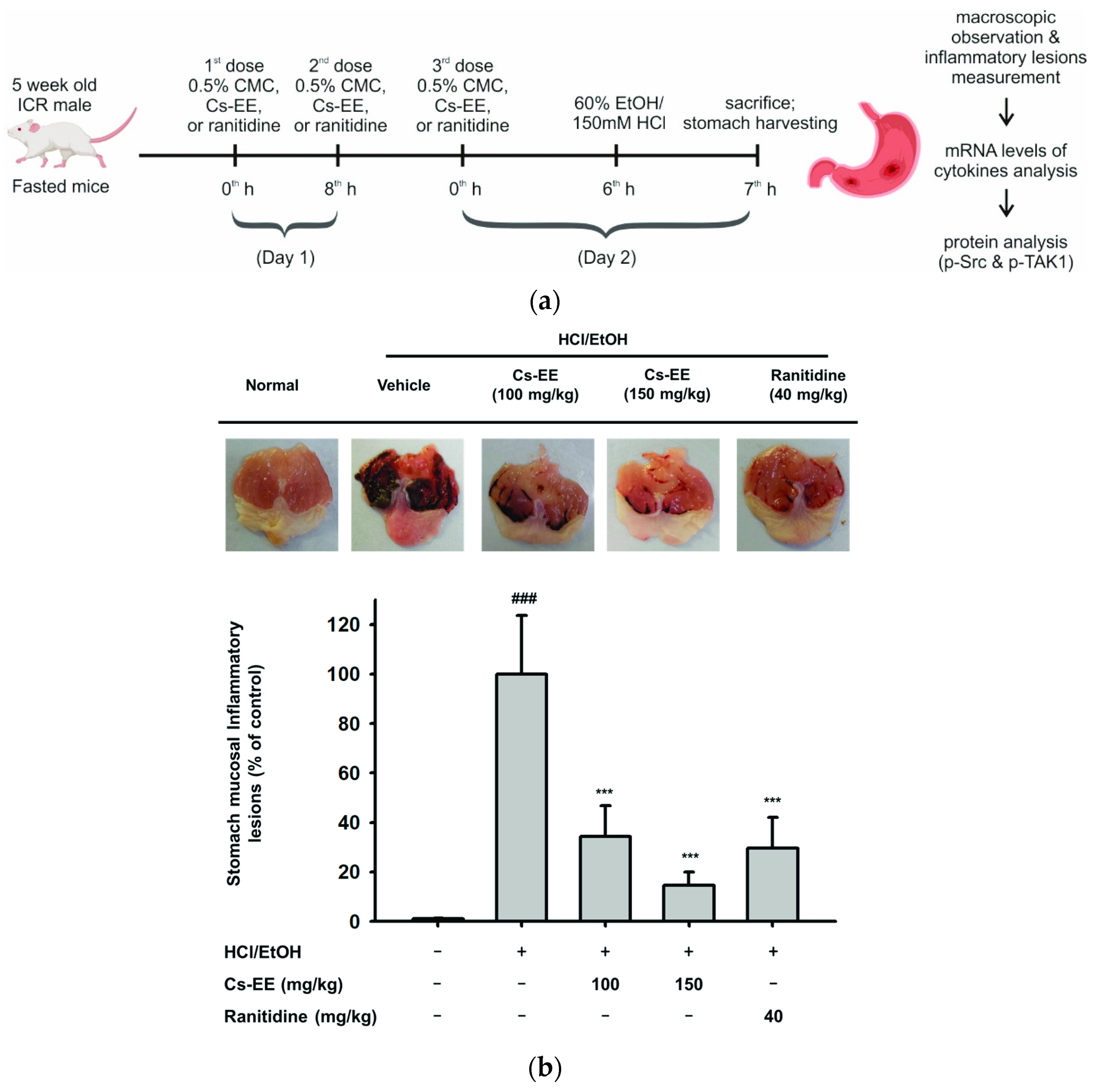
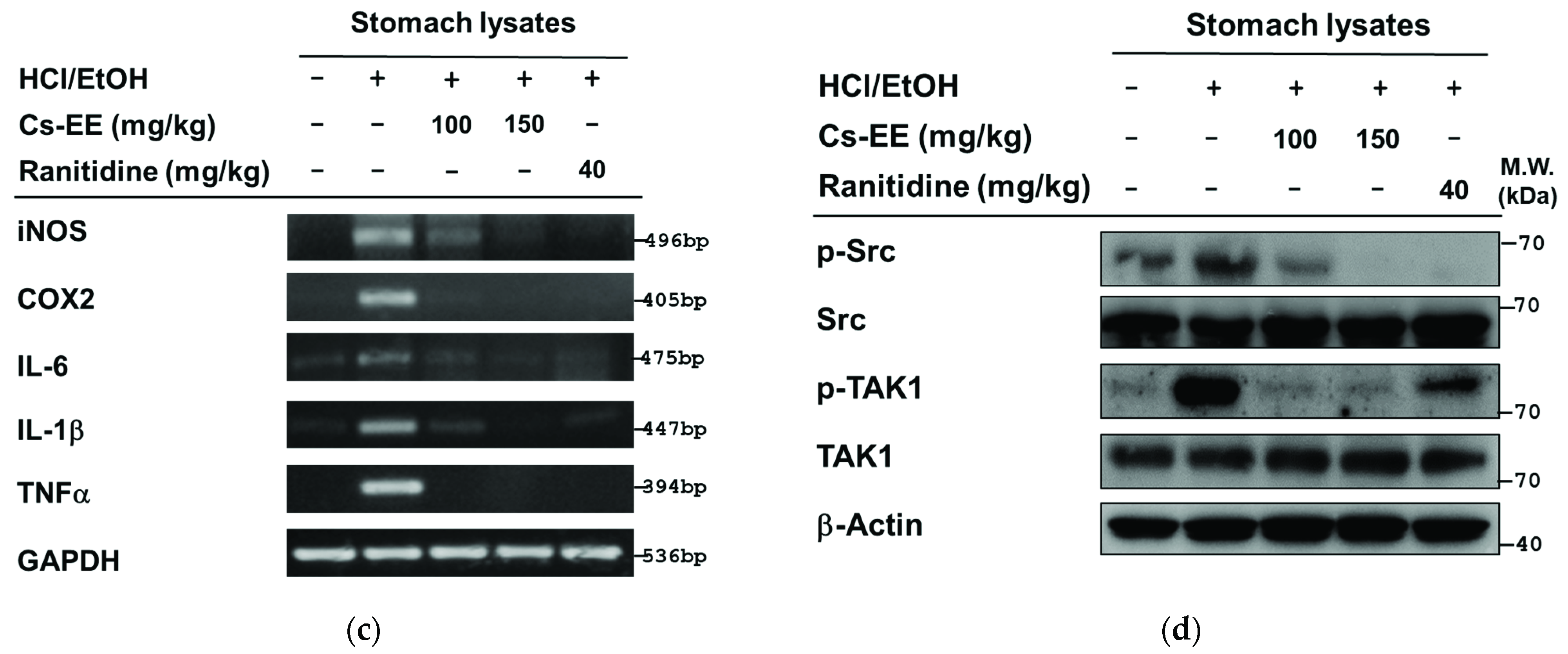
2.6. Effects of Cs-EE on HCl/EtOH-Induced Acute Gastritis Models
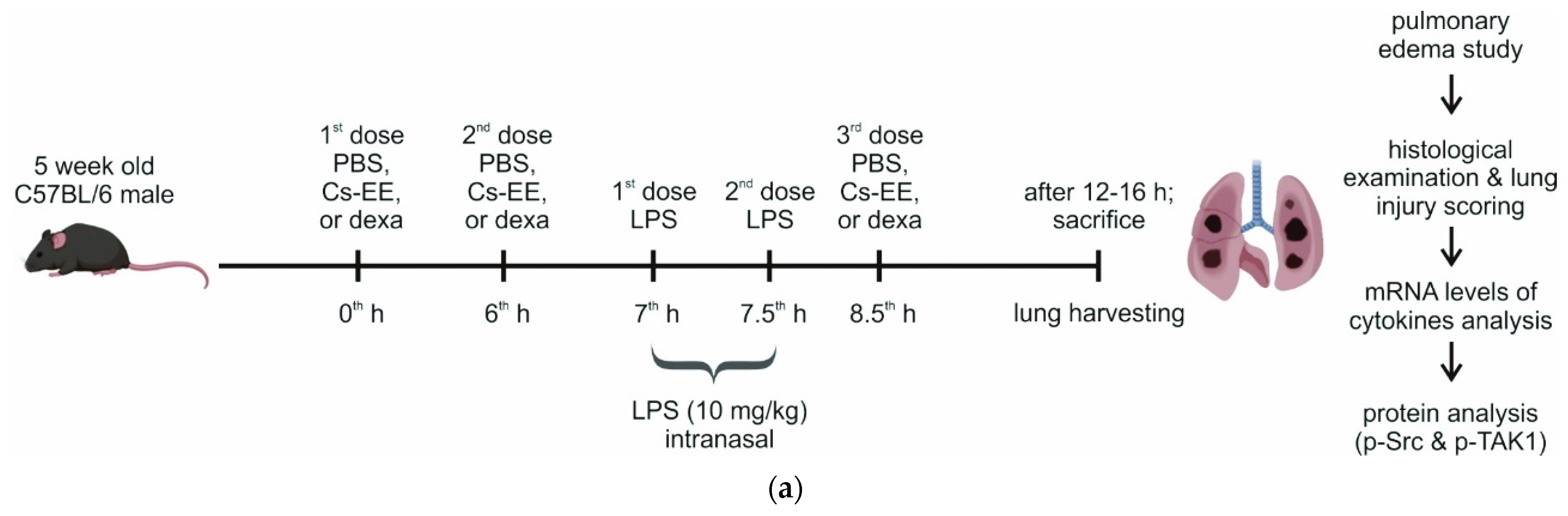
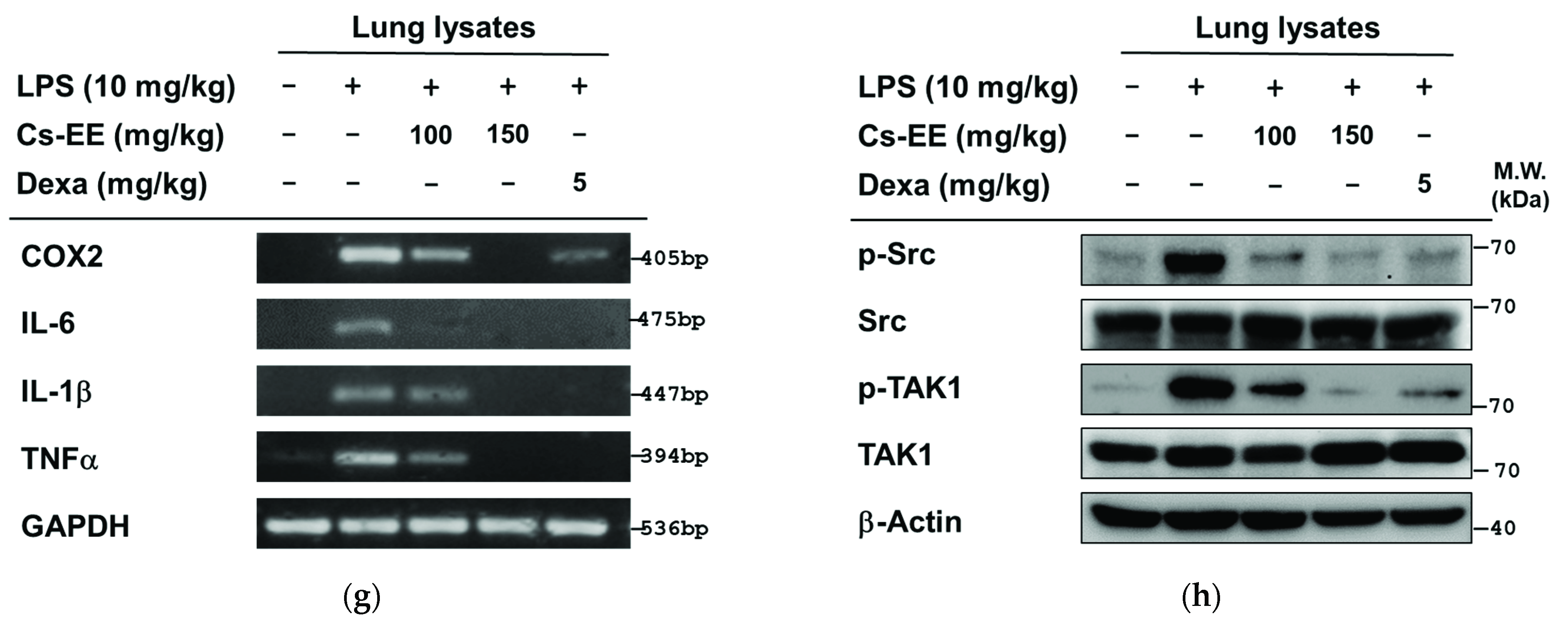
2.7. Effects of Cs-EE on LPS-Induced Acute Lung Injury Models
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Plant Collection, Specimen Information, and Cs-EE Preparation
4.3. Cell Culture and Treatment
4.4. Animals
4.5. Preparation of Peritoneal Macrophages
4.6. Determination of NO and PGE2 Production
4.7. Cell Viability Assay
4.8. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
4.9. mRNA Analysis by Quantitative Real-Time (q) Polymerase Chain Reaction (PCR) and Semiquantitative Reverse Transcriptase (RT)-PCR
4.10. Plasmid Transfection and Luciferase Reporter Gene Assay
4.11. Cell Lysate Extraction and Immunoblotting Analysis
4.12. Cellular Thermal Shift Assay (CETSA)
4.13. HCl/EtOH-Induced Acute Gastritis
4.14. LPS-Induced Acute Lung Injury
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| TAK1 | Transforming growth factor-β-activated kinase 1 |
| NF-κB | Nuclear factor-κB |
| AP-1 | Activator protein-1 |
| MAPKs | Mitogen-activated protein kinases |
| JNK | c-Jun N-terminal kinase |
| PI3K | Phosphoinositide 3-kinase |
| PGE2 | Prostaglandin E2 |
| COX-2 | Cyclooxygenase-2 |
| IL-1β | Interleukin-1β |
| IL-6 | Interleukin 6 |
| TNF-α | Tumor necrosis factor-alpha |
| MKK | Mitogen-activated protein kinase |
| iNOS | Inducible nitric oxide synthase |
References
- Janssens, S.; Beyaert, R. Role of Toll-like receptors in pathogen recognition. Clin. Microbiol. Rev. 2003, 16, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [Green Version]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- Arthur, J.S.C.; Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Qomaladewi, N.P.; Kim, M.Y.; Cho, J.Y. Autophagy and its regulation by ginseng components. J. Ginseng Res. 2019, 43, 349–353. [Google Scholar] [CrossRef]
- Yi, Y.-S.; Cho, J.Y.; Kim, D. Cerbera manghas methanol extract exerts anti-inflammatory activity by targeting c-Jun N-terminal kinase in the AP-1 pathway. J. Ethnopharmacol. 2016, 193, 387–396. [Google Scholar] [CrossRef]
- Chen, S.; Guo, J.; Feng, C.; Ke, Z.; Chen, L.; Pan, Y. The preoperative platelet-lymphocyte ratio versus neutrophil-lymphocyte ratio: Which is better as a prognostic factor in oral squamous cell carcinoma? Ther. Adv. Med. Oncol. 2016, 8, 160–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, M.; Singh, M.; Silakari, O. Inhibitors of switch kinase ’spleen tyrosine kinase’ in inflammation and immune-mediated disorders: A review. Eur. J. Med. Chem. 2013, 67, 434–446. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, A.; Kim, J.H.; Kim, J.H.; Yi, Y.S.; Cho, J.Y. Functional role of ginseng-derived compounds in cancer. J. Ginseng Res. 2018, 42, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.; Kim, M.-Y.; Cho, J.Y. Anti-inflammatory activities of Canarium subulatum Guillaumin methanol extract operate by targeting Src and Syk in the NF-κB pathway. J. Ethnopharmacol. 2019, 238, 111848. [Google Scholar] [CrossRef]
- Kim, J.K.; Choi, E.; Hong, Y.H.; Kim, H.; Jang, Y.J.; Lee, J.S.; Choung, E.S.; Woo, B.Y.; Hong, Y.D.; Lee, S.; et al. Syk/NF-κB-targeted anti-inflammatory activity of Melicope accedens (Blume) T.G. Hartley methanol extract. J. Ethnopharmacol. 2021, 271, 113887. [Google Scholar] [CrossRef]
- Huai, H.-Y.; Pei, S.-J. Plants used medicinally by folk healers of the Lahu people from the autonomous county of Jinping Miao, Yao, and Dai in Southwest China. Econ. Bot. 2004, 58, S265–S273. [Google Scholar] [CrossRef]
- Whitney, C.; Sin, M.; Hong, G.L.; Van, C.V.; Barber, K.S.; Tran, L.T.N. Conservation and ethnobotanical knowledge of a Hmong community in Long Lan, Luang Prabang, Lao People’s Democratic Republic. Ethnob. Res. Appl. 2014, 12, 643–658. [Google Scholar] [CrossRef] [Green Version]
- Stohs, S.J.; Ray, S.D. A review and evaluation of the efficacy and safety of Cissus quadrangularis extracts. Phytother. Res. 2013, 27, 1107–1114. [Google Scholar] [CrossRef]
- Fernandes, G.; Banu, J. Medicinal properties of plants from the genus Cissus: A review. J. Med. Plants Res. 2012, 6, 3080–3086. [Google Scholar]
- Wu, S.; Rajeshkumar, S.; Madasamy, M.; Mahendran, V. Green synthesis of copper nanoparticles using Cissus vitiginea and its antioxidant and antibacterial activity against urinary tract infection pathogens. Artif. Cells Nanomed. Biotechnol. 2020, 48, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Dutta, T.; Paul, A.; Majumder, M.; Sultan, R.A.; Emran, T.B. Pharmacological evidence for the use of Cissus assamica as a medicinal plant in the management of pain and pyrexia. Biochem. Biophys. Rep. 2019, 21, 100715. [Google Scholar] [CrossRef]
- Srisook, K.; Palachot, M.; Mongkol, N.; Srisook, E.; Sarapusit, S. Anti-inflammatory effect of ethyl acetate extract from Cissus quadrangularis Linn may be involved with induction of heme oxygenase-1 and suppression of NF-κB activation. J. Ethnopharmacol. 2011, 133, 1008–1014. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Helming, L.; Gordon, S. Alternative activation of macrophages: An immunologic functional perspective. Annu. Rev. Immunol. 2009, 27, 451–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef]
- Lee, Y.G.; Chain, B.M.; Cho, J.Y. Distinct role of spleen tyrosine kinase in the early phosphorylation of inhibitor of kappaB alpha via activation of the phosphoinositide-3-kinase and Akt pathways. Int. J. Biochem. Cell Biol. 2009, 41, 811–821. [Google Scholar] [CrossRef]
- Hong, Y.H.; Kim, J.H.; Cho, J.Y. Ranunculus bulumei methanol extract exerts anti-inflammatory activity by targeting Src/Syk in NF-κB signaling. Biomolecules 2020, 10, 546. [Google Scholar] [CrossRef] [Green Version]
- Li, C.-Y.; Zhou, Q.; Yang, L.-C.; Chen, Y.-H.; Hou, J.-W.; Guo, K.; Wang, Y.-P.; Li, Y.-G. Dual-specificity phosphatase 14 protects the heart from aortic banding-induced cardiac hypertrophy and dysfunction through inactivation of TAK1-P38MAPK/-JNK1/2 signaling pathway. Basic Res. Cardiol. 2016, 111, 19. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.K.; Park, J.G.; Hong, Y.H.; Aziz, N.; Park, S.H.; Kim, S.; Kim, E.; Cho, J.Y. Anti-inflammatory effects of Licania macrocarpa Cuatrec methanol extract target Src- and TAK1-mediated pathways. Evid. Based Complement. Alternat. Med. 2019, 2019, 4873870. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.G.; Kim, M.Y.; Cho, J.Y. Alisma canaliculatum ethanol extract suppresses inflammatory responses in LPS-stimulated macrophages, HCl/EtOH-induced gastritis, and DSS-triggered colitis by targeting Src/Syk and TAK1 activities. J. Ethnopharmacol. 2018, 219, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Kang, Y.-G.; Kim, Y.-J.; Lee, T.R.; Yoo, B.C.; Jo, M.; Kim, J.H.; Kim, J.-H.; Kim, D.; Cho, J.Y. Dehydroabietic acid suppresses inflammatory response via suppression of Src-, Syk-, and TAK1-mediated pathways. Int. J. Mol. Sci. 2019, 20, 1593. [Google Scholar] [CrossRef] [Green Version]
- Cooke, A.R. Drugs and gastric damage. Drugs 1976, 11, 36–44. [Google Scholar] [CrossRef]
- Kuzmich, N.N.; Sivak, K.V.; Chubarev, V.N.; Porozov, Y.B.; Savateeva-Lyubimova, T.N.; Peri, F. TLR4 signaling pathway modulators as potential therapeutics in inflammation and sepsis. Vaccines 2017, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Matute-Bello, G.; Downey, G.; Moore, B.B.; Groshong, S.D.; Matthay, M.A.; Slutsky, A.S.; Kuebler, W.M. An official American Thoracic Society workshop report: Features and measurements of experimental acute lung injury in animals. Am. J. Respir. Cell. Mol. Biol. 2011, 44, 725–738. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.P.; Son, K.H.; Chang, H.W.; Kang, S.S. Anti-inflammatory plant flavonoids and cellular action mechanisms. J. Pharmacol. Sci. 2004, 96, 229–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.W.; Lee, H.H.; Han, M.H.; Kim, G.Y.; Kim, W.J.; Choi, Y.H. Anti-inflammatory effects of genistein via suppression of the toll-like receptor 4-mediated signaling pathway in lipopolysaccharide-stimulated BV2 microglia. Chem. Biol. Interact. 2014, 212, 30–39. [Google Scholar] [CrossRef]
- Kim, S.H.; Park, J.G.; Sung, G.H.; Yang, S.; Yang, W.S.; Kim, E.; Kim, J.H.; Ha, V.T.; Kim, H.G.; Yi, Y.S.; et al. Kaempferol, a dietary flavonoid, ameliorates acute inflammatory and nociceptive symptoms in gastritis, pancreatitis, and abdominal pain. Mol. Nutr. Food Res. 2015, 59, 1400–1405. [Google Scholar] [CrossRef] [PubMed]
- Hämäläinen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: Genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-kappaB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-kappaB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediat. Inflamm. 2007, 2007, 45673. [Google Scholar]
- Lee, J.H.; Zhou, H.Y.; Cho, S.Y.; Kim, Y.S.; Lee, Y.S.; Jeong, C.S. Anti-inflammatory mechanisms of apigenin: Inhibition of cyclooxygenase-2 expression, adhesion of monocytes to human umbilical vein endothelial cells, and expression of cellular adhesion molecules. Arch. Pharm. Res. 2007, 30, 1318–1327. [Google Scholar] [CrossRef] [PubMed]
- Lorz, L.R.; Kim, D.; Kim, M.Y.; Cho, J.Y. Panax ginseng-derived fraction BIOGF1K reduces atopic dermatitis responses via suppression of mitogen-activated protein kinase signaling pathway. J. Ginseng Res. 2020, 44, 453–460. [Google Scholar] [CrossRef]
- Alm, A.S.; Li, K.; Chen, H.; Wang, D.; Andersson, R.; Wang, X. Variation of lipopolysaccharide-induced acute lung injury in eight strains of mice. Respir. Physiol. Neurobiol. 2010, 171, 157–164. [Google Scholar] [CrossRef]
- An, X.; Sun, X.; Hou, Y.; Yang, X.; Chen, H.; Zhang, P.; Wu, J. Protective effect of oxytocin on LPS-induced acute lung injury in mice. Sci. Rep. 2019, 9, 2836. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Fu, Z. Role of tilianin against acute lung injury in in vitro LPS-induced alveolar macrophage cells and an in vivo C57BL/6 Mice Model. J. Environ. Pathol. Toxicol. Oncol. 2020, 39, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Rho, T.; Jeong, H.W.; Hong, Y.D.; Yoon, K.; Cho, J.Y.; Yoon, K.D. Identification of a novel triterpene saponin from Panax ginseng seeds, pseudoginsenoside RT8, and its antiinflammatory activity. J. Ginseng Res. 2020, 44, 145–153. [Google Scholar] [CrossRef]
- Misko, T.P.; Schilling, R.J.; Salvemini, D.; Moore, W.M.; Currie, M.G. A Fluorometric assay for the measurement of nitrite in biological samples. Anal. Biochem. 1993, 214, 11–16. [Google Scholar] [CrossRef]
- Gerlier, D.; Thomasset, N. Use of MTT colorimetric assay to measure cell activation. J. Immunol. Methods 1986, 94, 57–63. [Google Scholar] [CrossRef]
- Choi, E.; Kim, E.; Kim, J.H.; Yoon, K.; Kim, S.; Lee, J.; Cho, J.Y. AKT1-targeted proapoptotic activity of compound K in human breast cancer cells. J. Ginseng Res. 2019, 43, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.; Son, Y.J.; Cho, J.Y. Thymoquinone suppresses IRF-3-mediated expression of type I interferons via suppression of TBK1. Int. J. Mol. Sci. 2018, 19, 1355. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.O.; Choi, E.; Shin, K.K.; Hong, Y.H.; Kim, H.G.; Jeong, D.; Hossain, M.A.; Kim, H.S.; Yi, Y.S.; Kim, D.; et al. Compound K, a ginsenoside metabolite, plays an antiinflammatory role in macrophages by targeting the AKT1-mediated signaling pathway. J. Ginseng Res. 2019, 43, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kim, M.Y.; Cho, J.Y. Euodia pasteuriana methanol extract exerts anti-inflammatory effects by targeting TAK1 in the AP-1 signaling pathway. Molecules 2020, 25, 5760. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lee, J.; Rhee, M.H.; Yu, T.; Baek, K.S.; Sung, N.Y.; Kim, Y.; Yoon, K.; Kim, J.H.; Kwak, Y.S.; et al. Molecular mechanism of protopanaxadiol saponin fraction-mediated anti-inflammatory actions. J. Ginseng Res. 2015, 39, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.O.; Kim, J.H.; Kim, S.; Kim, M.Y.; Hong, Y.H.; Kim, H.G.; Cho, J.Y. Gastroprotective effects of the nonsaponin fraction of Korean Red Ginseng through cyclooxygenase-1 upregulation. J. Ginseng Res. 2020, 44, 655–663. [Google Scholar] [CrossRef]
- Yu, T.; Rhee, M.H.; Lee, J.; Kim, S.H.; Yang, Y.; Kim, H.G.; Kim, Y.; Kim, C.; Kwak, Y.S.; Kim, J.H.; et al. Ginsenoside Rc from Korean Red Ginseng (Panax ginseng C.A. Meyer) attenuates inflammatory symptoms of gastritis, hepatitis and arthritis. Am. J. Chin. Med. 2016, 44, 595–615. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCR Type | Genes Name | Sequence (5′-3′) | |
|---|---|---|---|
| Semiquantitative RT-PCR | GAPDH | Forward | GAAGGTCGGTGTGAACGGAT |
| Reverse | AGTGATGGCATGGACTGTGG | ||
| iNOS | Forward | CAAGAGAACGGAGAACGGAGA | |
| Reverse | GATGGACCCCAAGCAAGACT | ||
| COX-2 | Forward | TGAGTACCGCAAACGCTTCT | |
| Reverse | TGGGAGGCACTTGCATTGAT | ||
| TNF-α | Forward | TTGACCTCAGCGCTGAGTTG | |
| Reverse | CCTGTAGCCCACGTCGTAGC | ||
| IL-6 | Forward | GCCTTCTTGGGACTGATGCT | |
| Reverse | TGGAAATTGGGGTAGGAAGGAC | ||
| IL-1β | Forward | CAGGATGAGGACATGAGCACC | |
| Reverse | CTCTGCAGACTCAAACTCCAC | ||
| qPCR | GAPDH | Forward | GAAGGTCGGTGTGAACGGAT |
| Reverse | AGTGATGGCATGGACTGTGG | ||
| iNOS | Forward | CAAGAGAACGGAGAACGGAGA | |
| Reverse | GATGGACCCCAAGCAAGACT | ||
| COX-2 | Forward | TGAGTACCGCAAACGCTTCT | |
| Reverse | TGGGAGGCACTTGCATTGAT | ||
| TNF-α | Forward | TTGACCTCAGCGCTGAGTTG | |
| Reverse | CCTGTAGCCCACGTCGTAGC | ||
| IL-6 | Forward | GCCTTCTTGGGACTGATGCT | |
| Reverse | TGGAAATTGGGGTAGGAAGGAC | ||
| IL-1β | Forward | CAGGATGAGGACATGAGCACC | |
| Reverse | CTCTGCAGACTCAAACTCCAC | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahmawati, L.; Aziz, N.; Oh, J.; Hong, Y.H.; Woo, B.Y.; Hong, Y.D.; Manilack, P.; Souladeth, P.; Jung, J.H.; Lee, W.S.; et al. Cissus subtetragona Planch. Ameliorates Inflammatory Responses in LPS-induced Macrophages, HCl/EtOH-induced Gastritis, and LPS-induced Lung Injury via Attenuation of Src and TAK1. Molecules 2021, 26, 6073. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26196073
Rahmawati L, Aziz N, Oh J, Hong YH, Woo BY, Hong YD, Manilack P, Souladeth P, Jung JH, Lee WS, et al. Cissus subtetragona Planch. Ameliorates Inflammatory Responses in LPS-induced Macrophages, HCl/EtOH-induced Gastritis, and LPS-induced Lung Injury via Attenuation of Src and TAK1. Molecules. 2021; 26(19):6073. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26196073
Chicago/Turabian StyleRahmawati, Laily, Nur Aziz, Jieun Oh, Yo Han Hong, Byoung Young Woo, Yong Deog Hong, Philaxay Manilack, Phetlasy Souladeth, Ji Hwa Jung, Woo Shin Lee, and et al. 2021. "Cissus subtetragona Planch. Ameliorates Inflammatory Responses in LPS-induced Macrophages, HCl/EtOH-induced Gastritis, and LPS-induced Lung Injury via Attenuation of Src and TAK1" Molecules 26, no. 19: 6073. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26196073