
Effective In Vitro Control of Two Phytopathogens of Agricultural Interest Using Cell-Free Extracts of Pseudomonas fluorescens and Chitosan

 ,
,
Abstract
:1. Introduction
2. Results
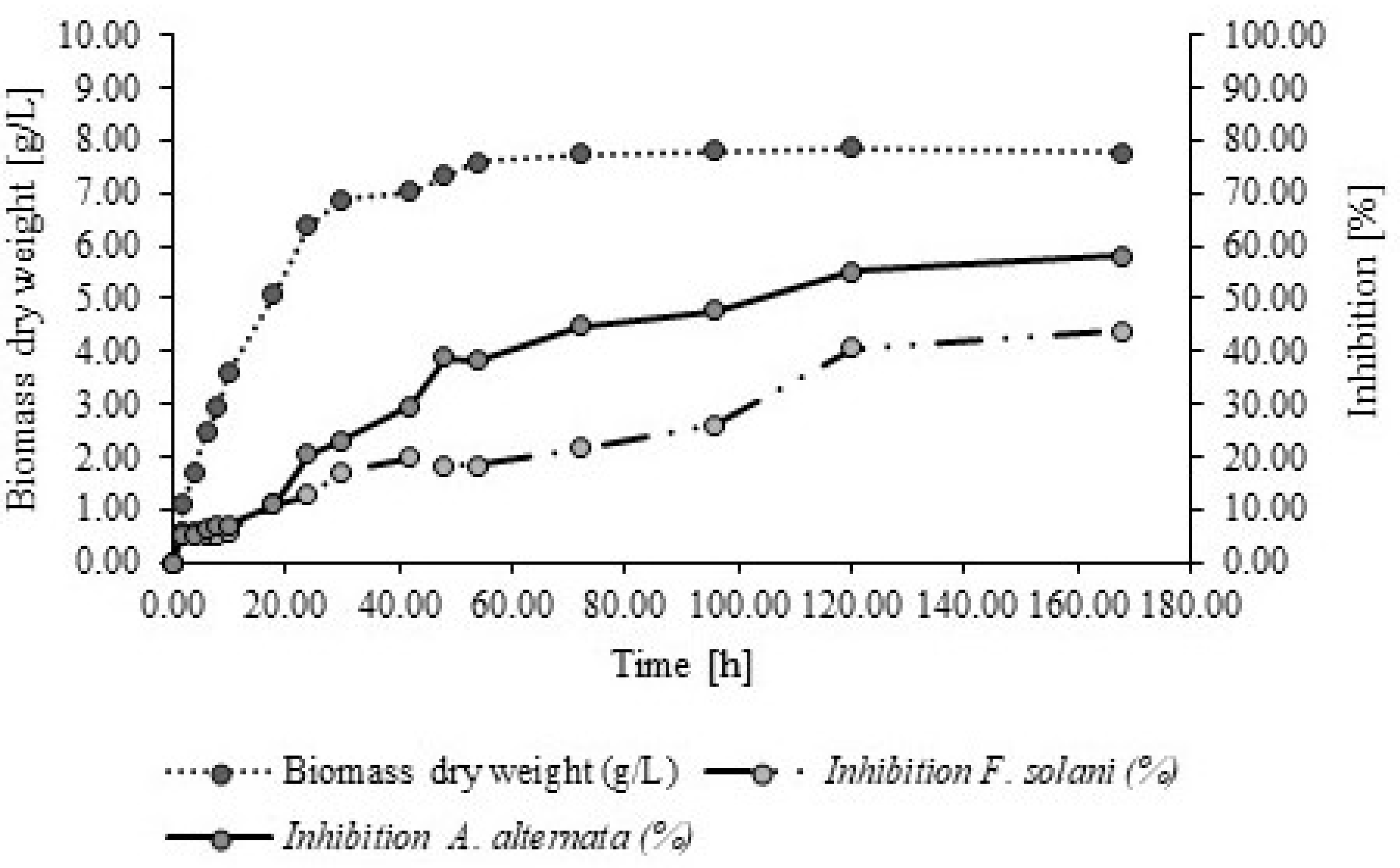
2.1. Culture Kinetics of P. fluorescens and the Cell-Free Extracts of Antifungal Activity of P. fluorescens
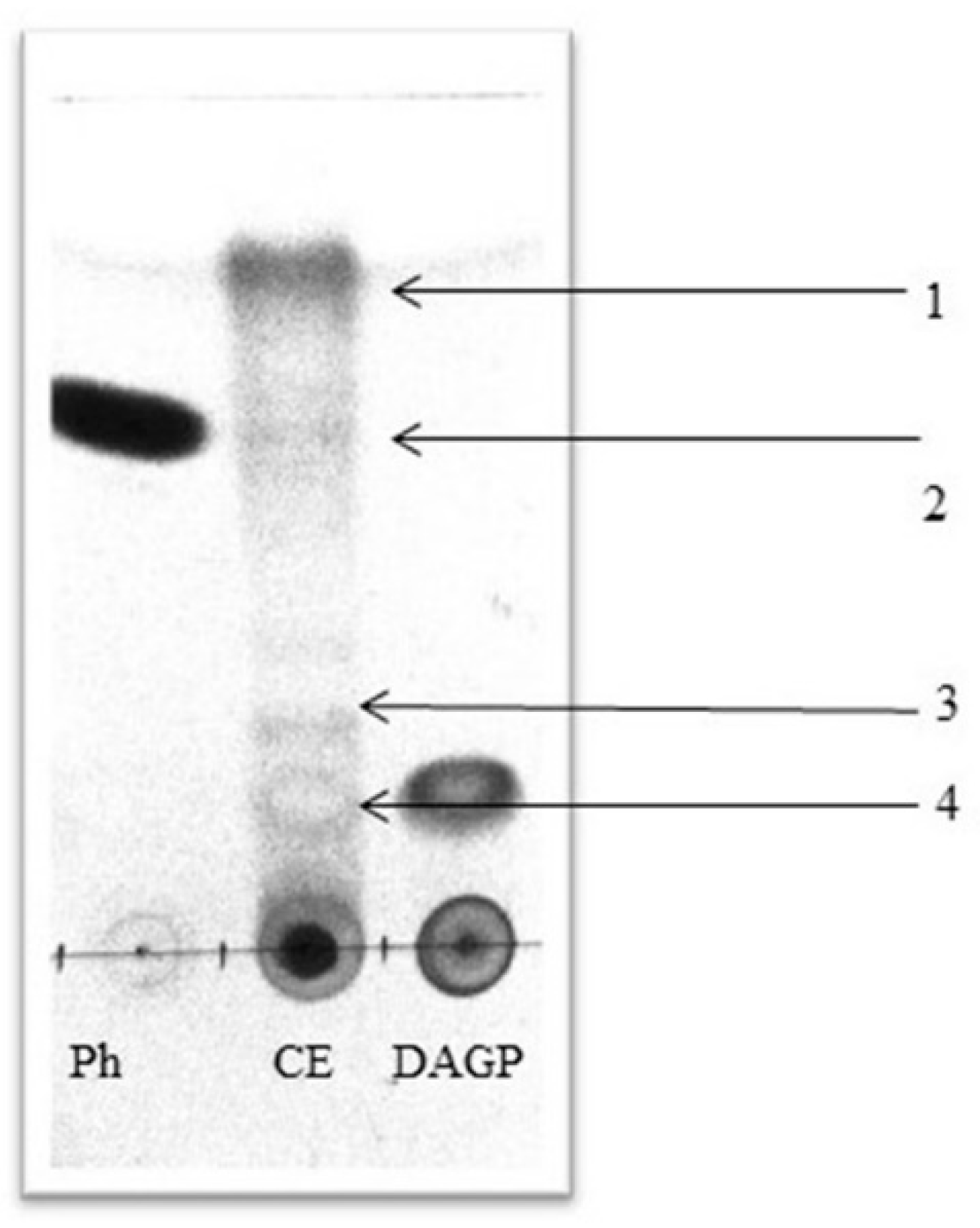
2.2. Identification of Antifungal Metabolites in Cell-Free Extracts of P. fluorescens
2.3. Combination of Chitosan and Cell-Free Extract of P. fluorescens and Its Antifungal Activity
2.4. Effect of the Combination of Chitosan and Cell-Free Extract of P. fluorescens on the Morphology of Mycelia and Conidia of A. alternata and F. solani
3. Discussion
4. Materials and Methods
4.1. Biological Material
4.2. Growth Kinetics of P. fluorescens
4.3. Obtention of Cell-Free Extracts of P. fluorescens and Evaluation of its Antifungal Activity
4.4. Identification of Antifungal Metabolites in Cell-Free Extracts of P. fluorescens
4.5. Preparation of Chitosan and Evaluation of its Antifungal Activity
4.6. Preparation of the Combination of Chitosan and Cell-Free Extract of P. fluorescens and the Evaluation of the Antifungal Activity
4.7. Effect of the Combination of Chitosan and Cell-Free Extract of P. fluorescens on the Morphology of Mycelia and Conidia
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Agrios, G.N. How patogens attack plants. In Plant Pathology; Agrios, G.N., Ed.; Academic Press: New York, NY, USA, 2005; pp. 175–205. [Google Scholar] [CrossRef]
- Jain, A.; Sarsaiya, S.; Wu, Q.; Lu, Y.; Shi, J. A review of plant leaf fungal diseases and its environment speciation. J. Bioeng. 2019, 10, 409–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, M.Y.; Cha, B.; Kim, J.C. Recent trends in studies on botanical fungicides in agriculture. J. Plant. Pathol 2013, 29, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrios, G.N. Plant pathogens and disease: General introduction. In Encyclopedia of Microbiology; Thomas, S., Ed.; Academic Press: New York, NY, USA, 2009; pp. 613–646. [Google Scholar] [CrossRef]
- Doehlemann, G.; Ökmen, B.; Zhu, W.; Sharon, A. Plant pathogenic fungi. Microbiol. Spectr. 2017, 5, 703–726. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Hsiang, T.; Bhadauria, V.; Chen, X.L.; Li, G. Plant Fungal Pathogenesis. Biomed. Res. Int. 2017. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Foster, G.D. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant. Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Shuping, D.S.; Eloff, J. The use of plants to protect plants and food against fungal pathogens: A review. Afr. J. Tradit. Complementary Altern. Med. 2014, 14, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M. The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study. J. Biol. Chem. 2014, 7, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Brent, K. Historical perspectives of fungicide resistance. In Fungicide Resistance in Crop Protection: Risk and Management; Thind, T.S., Ed.; CABI Publising: Oxfordshire, UK, 2012; pp. 3–18. [Google Scholar] [CrossRef]
- Kretschmer, M. Emergence of multi-drug resistance in fungal pathogens: A potential threat to fungicide performance in agriculture. In Fungicide Resistance in Crop Protection: Risk and Management; Thind, T.S., Ed.; CABI Publising: Oxfordshire, UK, 2012; pp. 251–267. [Google Scholar] [CrossRef]
- Leadbeater, A. The Role of the Fungicide Resistance Action Committee in Managing Resistance. In Fungicide Resistance in Crop Protection: Risk and Management; Thind, T.S., Ed.; CABI Publising: Oxfordshire, UK, 2012; pp. 103–121. [Google Scholar] [CrossRef]
- Copping, L.G.; Menn, J.J. Biopesticides: A review of their action, applications and efficacy. Pest. Manag. Sci. 2002, 56, 651–676. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant. Sci 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Salma, M.; Jogen, C.K. A review on the use of biopesticides in insect pest management. Int. J. Adv. Sci. Technol. 2011, 1, 169–178. [Google Scholar] [CrossRef]
- Panpatte, D.G.; Jhala, Y.K.; Shelat, H.N.; Vyas, R.V. Pseudomonas fluorescens: A promising biocontrol agent and pgpr for sustainable agriculture. In Microbial Inoculants in Sustainable Agricultural Productivity Vol. 1: Research Perspectives; Singh, D.P., Singh, H.B., Prabha, R., Eds.; Springer: New York, NY, USA, 2016; pp. 257–270. [Google Scholar] [CrossRef]
- Channarayappa, C.; Biradar, D.P. Plant growth promoting bacteria. In Rhizosphere Engineering. Soil Basics, Management, and Rhizosphere; Taylor and Francis Group, Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 701–729. [Google Scholar] [CrossRef]
- Jani, J.; Parvez, N.; Mehta, D. Metabolites of pseudomonads: A new avenue of plant health management. new horizons. In Insect Science: Towards Sustainable Pest Management; Chakravarthy, A.K., Ed.; Springer: New York, NY, USA, 2015; pp. 61–69. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by Fluorescent Pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef]
- Rodríguez-Romero, V.M.; Villanueva-Arce, R.; Trejo-Raya, A.B.; Bautista-Baños, S. Quitosano y extractos de Pseudomonas fluorescens para el control de Alternaria alternata en jitomate (Solanum lycopersicum). Rev. Mex. Fitopatol. 2019, 37, 202–219. [Google Scholar] [CrossRef]
- Vagelas, I.; Kalorizou, H.; Papachatzis, A.; Botu, M. Bioactivity of olive oil mill wastewater against plant pathogens and post-harvest diseases. Biotechnol. Biotechnol. Equip. 2009, 23, 1217–1229. [Google Scholar] [CrossRef]
- Xu, X.M.; Jeffries, P.; Pautasso, M.; Jeger, M.I. Combined use of biocontrol agents to manage plant diseases in theory and practice. Phytopathology 2011, 101, 1024–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylla, J.; Beatrix, A.; Kruger, E.; Wohanka, W. Control of Botrytis cinerea in strawberries by biological control agents applied as single or combined treatments. Eur. J. Plant. Pathol. 2015, 143, 461–471. [Google Scholar] [CrossRef]
- Romanazzi, G.; Feliziani, E.; Sivakumar, D. Chitosan, a biopolymer with triple action on post-harvest decay of fruit and vegetables: Eliciting, antimicrobial and film-forming properties. Front. Microbiol. 2018, 9, 2475. [Google Scholar] [CrossRef] [PubMed]
- Badawy, M.E.I.; Rabea, E.I. Potential of the biopolymer chitosan with different molecular weights to control post-harvest gray mold of tomato fruit. Postharvest Biol. Technol. 2009, 51, 110–117. [Google Scholar] [CrossRef]
- Sharma, S.; Barman, K.; Siddiqui, M.W. Chitosan: Properties and roles in post-harvest quality preservation of horticultural crops. In Eco-Friendly Technology for Postharvest Produce Quality; Siddiqui, M.W., Ed.; Academic Press: Cambgridge, MA, USA, 2016; pp. 269–295. [Google Scholar] [CrossRef]
- Abbas, E.; Jochen, S.; Birgit, H.; Frank, W.; Ralf, T.V. Assessment of the antifungal activity of selected biocontrol agents and their secondary metabolites against Fusarium graminearum. Eur. J. Plant. Pathol. 2018, 150, 91–103. [Google Scholar] [CrossRef]
- Omnia, M.; Rania, A.A.H.; Mona, H.B.; Hussien, E.M. Antifungal activity of pseudomonas fluorescens metabolites against some phytopathogenic fungi. Middle East. J. Appl. Sci. 2020, 10, 158–168. [Google Scholar] [CrossRef]
- Yarzabal, L.A.; Chica, J.E. Role of rhizobacterial secondary metabolites in crop protection against agricultural pests and diseases. In Microbial Secondary Metabolites Biochemistry and Applications; Kumar, V., Pandey, A., Eds.; Elsiever: Amsterdam, The Netherlands, 2018; pp. 31–53. [Google Scholar]
- Fallahzadeh, V.; Ahmadzadeh, M.; Sharifi, R. Growth and pyoverdine production kinetics of Pseudomonas aeruginosa 7NSK2 in an experimental fermentor. J. Agric. Sci. Technol. 2010, 6, 107–115. [Google Scholar]
- Beyenala, H.; Nee, S.; Zbigniew, L. The double substrate growth kinetics of Pseudomonas aeruginosa. Enzyme Microb. Technol. 2003, 1, 92–98. [Google Scholar] [CrossRef]
- Boutheina, M.T.; Abdallah Rania, A.B.; Nawaim, A.; Mejda, D.R. Antifungal potential of extracellular metabolites from penicillium spp. and aspergillus spp. naturally associated to potato against fusarium species causing tuber dry rot. J. Microb. Biochem. Technol. 2017, 9, 181–190. [Google Scholar] [CrossRef]
- Sikander, A.; Syeda, T.A.; Syeda, F.N.; Muneeba, S. Strategies and kinetics of industrial fermentation for the mass production of various primary and secondary metabolites from microbes. Eur. J. Pharm. Med. Res. 2018, 5, 595–606. [Google Scholar]
- Manikandan, R.; Saravanakumar, D.; Rajendran, L.; Raguchander, T.; Samiyappan, R. Standardization of liquid formulation of Pseudomonas fluorescens Pf1 for its efficacy against Fusarium wilt of tomato. Biol. Control. 2010, 54, 83–89. [Google Scholar] [CrossRef]
- Brown, A.G.; Luke, R.K.J. Siderophore production and utilization b milk spoilage pseudomonas species. J. Dairy Sci. 2010, 93, 1355–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buono, N.I.; Ulla, E.L. Efectos de La Inoculación Con Bacterias Solubilizadoras de Fosfato En Tabaco (Nicotiana tabacum L.) Pimiento (Capsicum annuum L.) En Condiciones Controladas. Rev. Agron. Noroeste Arg. 2014, 34, 130–131. [Google Scholar]
- Chin-A-Woeng, T.F.; Thomas-Oates, J.E.; Lugtenberg, B.J.; Bloemberg, G.V. Introduction of the phzh gene of pseudomonas chlororaphis pcl1391 extends the range of biocontrol abilit of phenazine-1-carboxlic acid-producing pseudomonas spp. strains. Mol. Plant-Microbe Interact. 2001, 14, 1006–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwivedi, D.; Johri, B.N. Antifungals from Fluorescent Pseudomonads: Biosnthesis and Regulation. Curr. Sci. 2003, 85, 1693–1703. [Google Scholar]
- Georgakopoulos, D.G.; Hendson, M.; Panopoulos, N.J.; Schroth, M.N. Cloning of a phenazine biosnthetic locus of pseudomonas aureofaciens pgs12 and analsis of its expression in vitro with the ice nucleation reporter gene. Appl. Environ. Microbiol. 1994, 60, 2931–2938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keel, C.; Oberhansli, T.; Wirthner, P.; Voisard, C.; Haas, D.; Défago, G. Pseudomonas as Antagonists of Plant Pathogens in the Rhizosphere: Role of the Antibiotic 2,4-Diacetlphloroglucinol in the Suppression of Black Rot of Tobacco. Smbiosis 1994, 9, 327–341. [Google Scholar]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of blue agar CAS assay for siderophore detection. J. Microbiol. Biol. Educ. 2011, 12, 51–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavrodi, O.V.; McSpadden Gardener, B.B.; Mavrodi, D.V.; Bonsall, R.F.; Weller, D.M.; Thomashow, L.S. Genetic diversit of PhlD from 2,4-diacetlphloroglucinol-producing fluorescent Pseudomonas spp. Phtopatholog 2001, 91, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picard, C.; Di Cello, F.; Ventura, M.; Fani, R.; Guckert, A. Frequenc and biodiversit of 2,4-diacetlphloroglucinol-producing bacteria isolated from the maize rhizosphere at different stages of plant growth. Appl. Environ. Microbiol. 2000, 66, 948–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, K.R.N.; Choudary, K.A.; Reddy, M.S. Antifungal metabolites of Pseudomonas fluorescens isolated from rhizosphere of rice crop. J. Mycol. Pl. Pathol. 2007, 37, 1–5. [Google Scholar]
- Tripathi, R.K.; Gottlieb, D. Mechanism of Action of the Antifungal Antibiotic Prrolnitrin. J. Bacteriol. Res. 1969, 100, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Huang, X.; Tang, L.; Wu, D.; Xu, Y. Global control of gaca in secondar metabolism, primar metabolism, secretion sstems, and motilit in the rhizobacterium Pseudomonas aeruginosa M18. J. Bacteriol. Res. 2013, 195, 3387–3400. [Google Scholar] [CrossRef] [Green Version]
- Weller, D.M. Pseudomonas biocontrol agents of soilborne pathogens: Looking back over 30 years. Phytopathology 2003, 97, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goy, R.C.; Britto, D.; Assis, O.B.G. A review of the antimicrobial activity of chitosan. Assis Polim. Cienc. E Tecnol. 2009, 19, 241–247. [Google Scholar] [CrossRef]
- Gol, N.B.; Patel, P.R.; Rao, T.V.R. Improvement of quality and shelflife of strawberries with edible coatings enriched with chitosan. Postharv. Biol. Technol. 2013, 85, 185–195. [Google Scholar] [CrossRef]
- Bautista-Baños, S.; Hernandez-Lauzardo, A.N.; Velazquez-del Valle, M.G.; Hernandez-Lopez, M.; Ait Barka, E.; Bosquez-Molina, E. Chitosan as a potential natural compound to control pre and postharvest diseases of horticultural commodities. Crop. Prot. 2016, 25, 108–118. [Google Scholar] [CrossRef]
- Bautista-Baños, S.; Romanazzi, G.; Jiménez-Aparicio, A. Chitosan in the Preservation of Agricultural Commodities; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Bautista-Baños, S.; Ventura-Aguilar, R.I.; Correa-Pacheco, Z.; Corona-Rangel, M.L. Chitosan: A versatile antimicrobial polysaccharide for fruit and vegetables in postharvest—a review. Rev. Chapingo Ser. Hortic. 2017, 12, 103–121. [Google Scholar] [CrossRef]
- Ing, Y.L.; Zin, N.M.; Sarwar, A.; Katas, H. Antifungal activity of chitosan nanoparticles and correlation with their physical properties. Int. J. Biomater. 2012, 2012, 632698. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Junior, E.N.; de Melo, I.S.; Teixeira, T. Changes in Hyphal Morphology due to Chitosan Treatment in Some Fungal Species. Braz. Arch. Biol. Technol. 2012, 555, 637–646. [Google Scholar] [CrossRef]
- Atia, M.M.M.; Buchenauer, H.; Aly, A.Z.; Abou-Zaid, M.I. Antifungal activity of chitosan against Phytophthora infestans and activation of defence mechanisms in tomato to late blight. Biological. Agric. Hortic. 2002, 23, 175–197. [Google Scholar] [CrossRef]
- Gupta, C.P.; Dubey, R.C.; Kang, S.C.; Maheshwari, D.K. Antibiosis-mediated necrotrophic effect of Pseudomonas GRC2 against two fungal plant pathogens. Curr. Sci. 2001, 81, 91–94. [Google Scholar]
- Bhatia, S.; Bhatia, S.; Dubey, R.C.; Maheshwari, D.K. Antagonistic effect of fluorescent pseudomonads against Macrophomina phaseolina that causes charcoal rot of groundnut. NISCAIR-CSIR 2003, 41, 14142. [Google Scholar]
- Weber, R.W.S. Vacuoles and the fungal lifestyle. Mycologist 2002, 16, 10–20. [Google Scholar] [CrossRef]
- Bent, K.; Morton, A.G. Formation and nature of swollen hyphae in Penicillium and related fungi. Trans. Brit. Mycol. Soc. 1963, 46, 401–408. [Google Scholar] [CrossRef]
- Palma-Guerrero, J.; Lopez-Jimenez, J.A.; Pérez-Berná, A.J.; Huang, I.C.; Jansson, H.B.; Salinas, J.; Villalaín, J.; Read, N.D.; Lopez-Llorca1, L.V. Membrane fluidity determines sensitivity of filamentous fungi to chitosan. Mol. Microb. 2010, 75, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Delidow , B.C.; Lynch , J.P.; Peluso , J.J.; White , B.A. Polymerase Chain Reaction. In Basic DNA and RNA protocols, Methods in Molecular Biology; Hanwood Adrian, J., Ed.; Human Press: Totowa, NJ, USA, 1996; Volume 58. [Google Scholar]
- Fekria, M.A.; Saber, A.; Ahmed, A.; Abdelhafez, B.; Enas, A.; Hassan, B.; Elshahat, M.; Ramadan, C. Characterization of fluorescent pseudomonads isolates and their efficiency on the growth promotion of tomato plant. Ann. Agric. Sci. 2015, 60, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Korsten, L.; Jager, E.E. Mode of action of Bacillus subtilis for control of avocado post-harvest pathogens. SAAGA Yearb 1988, 18, 124–130. [Google Scholar]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Hickey, P.C.; Swift, S.R.; Roca, M.G.; Read, N.D. Live-cell imaging of filamentous fungi using vital fluorescent dyes and confocal microscopy. Methods Microbiol. 2004, 34, 63–87. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytopathogens | |||||
|---|---|---|---|---|---|
| A. alternata | F. solani | ||||
| Controls and Incubation Time (h) of P. fluorescens | Biomass Dry Weight (g/L) | Mycelial Growth * (cm) | Inhibition (%) | Mycelial Growth * (cm) | Inhibition (%) |
| Distilled water | NA | 5.50 ± 0.00 a | 0.00 | 5.50 ± 0.00 a | 0.00 |
| Captan® 0.40 % | NA | 4.23 ± 0.10 cde | 23.09 | 4.31 ± 0.17 bc | 21.64 |
| 2 | 1.09 ± 0.05 | 5.20 ± 0.00 ab | 5.45 | 5.17 ± 0.06 ab | 6.00 |
| 4 | 1.74 ± 0.12 | 5.20 ± 0.00 ab | 5.45 | 5.18 ± 0.04 ab | 5.82 |
| 6 | 2.47 ± 0.03 | 5.15 ± 0.00 ab | 6.36 | 5.20 ± 0.00 ab | 5.45 |
| 8 | 2.94 ± 0.09 | 5.13 ± 0.00 ab | 6.73 | 5.20 ± 0.00 ab | 5.45 |
| 10 | 3.58 ± 0.03 | 5.10 ± 0.00 bc | 7.27 | 5.18 ± 0.04 ab | 5.82 |
| 18 | 5.08 ± 0.02 | 4.90 ± 0.00 bc | 10.91 | 4.89 ± 0.26 abc | 11.09 |
| 24 | 6.39 ± 0.03 | 4.37 ± 0.00 cd | 20.55 | 4.80 ± 0.43 abc | 12.73 |
| 30 | 6.88 ± 0.07 | 4.22 ± 0.03 de | 23.27 | 4.55 ± 0.7 bcd | 17.27 |
| 42 | 7.02 ± 0.03 | 3.86 ± 0.36 e | 29.82 | 4.40 ± 0.32 cd | 20.00 |
| 48 | 7.31 ± 0.03 | 3.36 ± 0.11 f | 38.91 | 4.48 ± 0.18 cd | 18.55 |
| 54 | 7.58 ± 0.01 | 3.4 ± 0.06 f | 38.18 | 4.49 ± 0.02 cd | 18.36 |
| 72 | 7.75 ± 0.03 | 3.03 ± 0.24 g | 44.91 | 4.31 ± 0.17 d | 21.64 |
| 96 | 8.9 ± 0.02 | 2.87 ± 0.17 gh | 47.82 | 4.07 ± 0.38 d | 26.00 |
| 120 | 8.88 ± 0.01 | 2.47 ± 0.41 gh | 55.09 | 3.26 ± 0.22 e | 40.73 |
| 168 | 8.93 ± 0.08 | 2.30 ± 0.19 h | 58.18 | 3.09 ± 0.41 e | 43.82 |
| Sample | Rf |
|---|---|
| CE1 | 0.16 |
| CE2 | 0.26 |
| CE3 | 0.61 |
| CE4 | 0.80 |
| Ph | 0.61 |
| DAGP | 0.17 |
| A. alternata | F. solani | |||
|---|---|---|---|---|
| Treatments | Mycelial Growth * (cm) | Inhibition (%) | Mycelial Growth * (cm) | Inhibition (%) |
| Distilled water | 5.50 ± 0.00 a | 0.00 | 5.50 ± 0.00 a | 0.00 |
| Captan® (0.25% p/v) | 4.23 ± 0.10 b | 23.09 | 4.31 ± 0.17 b | 21.64 |
| Chitosan 1.5% | 3.67 ± 0.17 c | 33.27 | 4.72 ± 0.14 b | 14.18 |
| Cell-free extracts of P. fluorescens (sample at 120 h) | 2.47 ± 0.41 d | 55.09 | 3.26 ± 0.22 c | 40.73 |
| Combination of cell-free extracts of P. fluorescens with chitosan [50%–1.5% (v/v)] | 2.19 ± 0.06 d | 60.18 | 3.06 ± 0.10 c | 44.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trejo-Raya, A.B.; Rodríguez-Romero, V.M.; Bautista-Baños, S.; Quiroz-Figueroa, F.R.; Villanueva-Arce, R.; Durán-Páramo, E. Effective In Vitro Control of Two Phytopathogens of Agricultural Interest Using Cell-Free Extracts of Pseudomonas fluorescens and Chitosan. Molecules 2021, 26, 6359. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216359
Trejo-Raya AB, Rodríguez-Romero VM, Bautista-Baños S, Quiroz-Figueroa FR, Villanueva-Arce R, Durán-Páramo E. Effective In Vitro Control of Two Phytopathogens of Agricultural Interest Using Cell-Free Extracts of Pseudomonas fluorescens and Chitosan. Molecules. 2021; 26(21):6359. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216359
Chicago/Turabian StyleTrejo-Raya, Ariadna Berenice, Víctor Manuel Rodríguez-Romero, Silvia Bautista-Baños, Francisco Roberto Quiroz-Figueroa, Ramón Villanueva-Arce, and Enrique Durán-Páramo. 2021. "Effective In Vitro Control of Two Phytopathogens of Agricultural Interest Using Cell-Free Extracts of Pseudomonas fluorescens and Chitosan" Molecules 26, no. 21: 6359. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216359