
Neuroprotective Effects of B-Type Cinnamon Procyanidin Oligomers on MPP+-Induced Apoptosis in a Cell Culture Model of Parkinson’s Disease

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Cytotoxic Effects of MPP+ and CPO-B
2.2. CPO-B Protects SH-SY5Y Cells from MPP+-Induced Intracellular ROS Production and Apoptosis
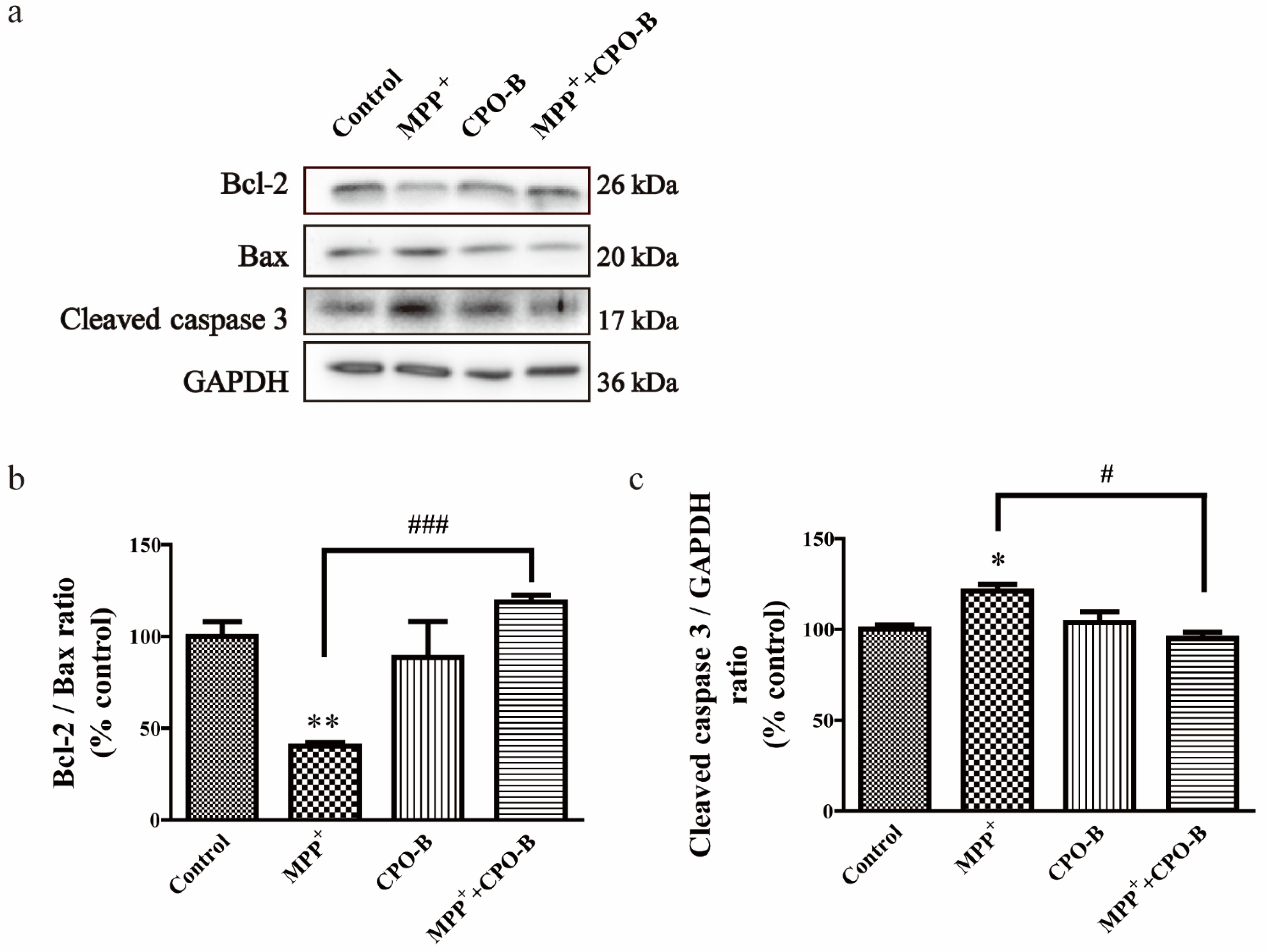
2.3. CPO-B Blocks MPP+-Induced Alteration of Apoptosis-Associated Proteins in SH-SY5Y Cells
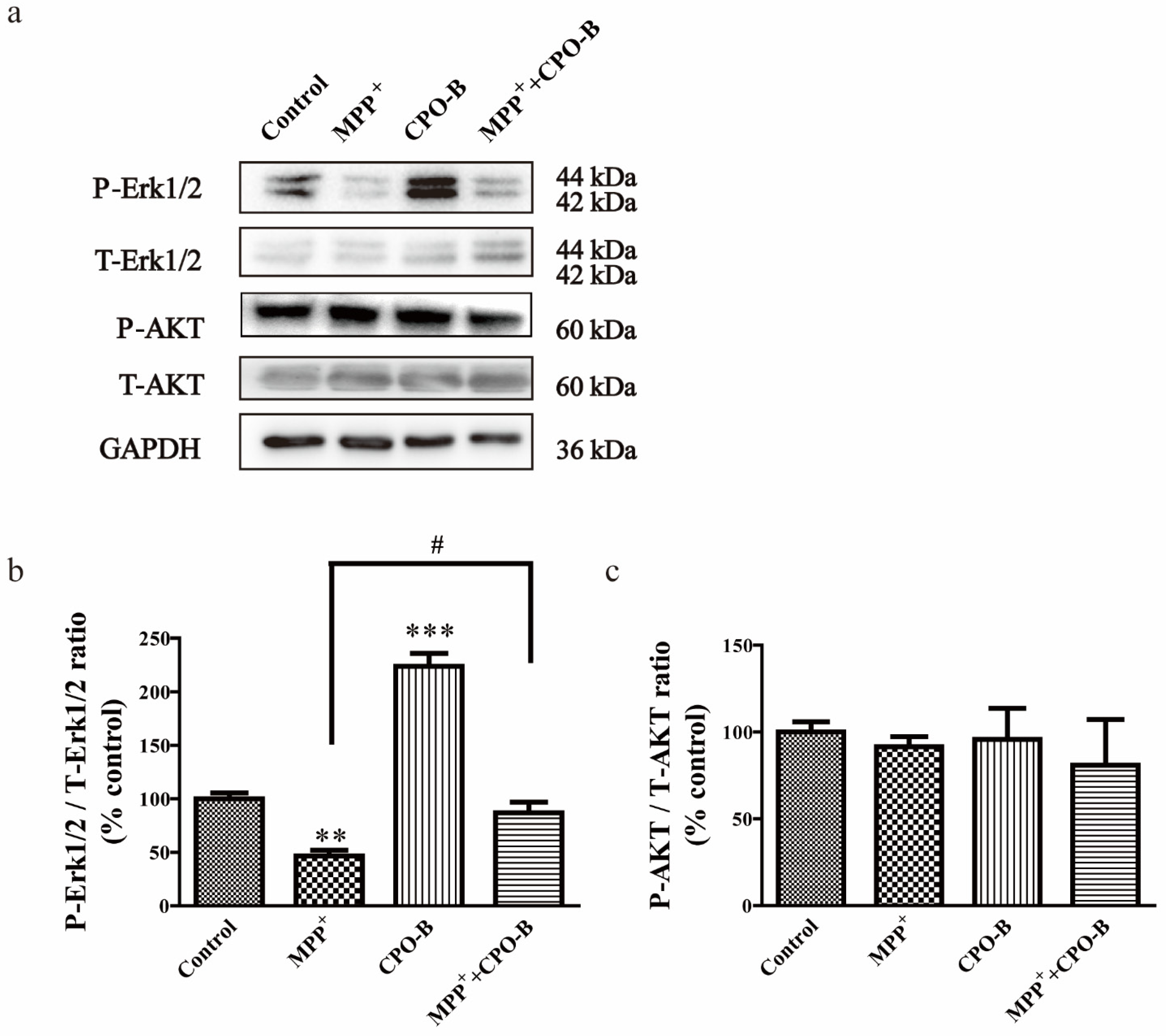
2.4. CPO-B Exerts Neuroprotection through the Upregulation of Phosphorylation of Erk1/2 but Not AKT Expression
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. The Extraction of Procyanidin
4.3. Design of Experiments
4.4. Cell Viability
4.5. Measurement of Intracellular ROS
4.6. Nuclear Staining with Hoechst 33258
4.7. Western Blotting Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Blesa, J.; Przedborski, S. Parkinson’s disease: Animal models and dopaminergic cell vulnerability. Front. Neuroanat. 2014, 8, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzaei, M.H.; Tewari, D.; Momtaz, S.; Arguelles, S.; Nabavi, S.M. Targeting ERK signaling pathway by polyphenols as novel therapeutic strategy for neurodegeneration. Food Chem. Toxicol. 2018, 120, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Mebratu, Y.; Tesfaigzi, Y. How ERK1/2 activation controls cell proliferation and cell death: Is subcellular localization the answer? Cell Cycle 2009, 8, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Dexter, D.T.; Jenner, P. Parkinson disease: From pathology to molecular disease mechanisms. Free Radic. Biol. Med. 2013, 62, 132–144. [Google Scholar] [CrossRef]
- Jha, S.K.; Jha, N.K.; Kar, R.; Ambasta, R.K.; Kumar, P. p38 MAPK and PI3K/AKT Signalling Cascades inParkinson’s Disease. Int. J. Mol. Cell. Med. 2015, 4, 67–86. [Google Scholar]
- Subramaniam, S.R.; Chesselet, M.F. Mitochondrial dysfunction and oxidative stress in Parkinson’s disease. Prog. Neurobiol. 2013, 106-107, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Teng, L.; Kou, C.; Lu, C.; Xu, J.; Xie, J.; Lu, J.; Liu, Y.; Wang, Z.; Wang, D. Involvement of the ERK pathway in the protective effects of glycyrrhizic acid against the MPP+-induced apoptosis of dopaminergic neuronal cells. Int. J. Mol. Med. 2014, 34, 742–748. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.; Zhang, W.; Lu, F.; Gao, L.; Gao, G. Resveratrol attenuates MPP(+)-induced mitochondrial dysfunction and cell apoptosis via AKT/GSK-3beta pathway in SN4741 cells. Neurosci. Lett. 2017, 637, 50–56. [Google Scholar] [CrossRef]
- Wang, G.; Ma, W.; Du, J. beta-Caryophyllene (BCP) ameliorates MPP+ induced cytotoxicity. Biomed. Pharmacother. 2018, 103, 1086–1091. [Google Scholar] [CrossRef]
- Hu, M.; Li, F.; Wang, W. Vitexin protects dopaminergic neurons in MPTP-induced Parkinson’s disease through PI3K/Akt signaling pathway. Drug Des. Dev. Ther. 2018, 12, 565–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalevich, J.; Langford, D. Considerations for the use of SH-SY5Y neuroblastoma cells in neurobiology. Methods Mol. Biol. 2013, 1078, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, Y.T.; Lau, W.K.; Yu, M.S.; Lai, C.S.; Yeung, S.C.; So, K.F.; Chang, R.C. Effects of all-trans-retinoic acid on human SH-SY5Y neuroblastoma as in vitro model in neurotoxicity research. Neurotoxicology 2009, 30, 127–135. [Google Scholar] [CrossRef]
- Rao, P.V.; Gan, S.H. Cinnamon: A multifaceted medicinal plant. Evid. Based Complement. Altern. Med. 2014, 2014, 642942. [Google Scholar] [CrossRef] [Green Version]
- Frydman-Marom, A.; Levin, A.; Farfara, D.; Benromano, T.; Scherzer-Attali, R.; Peled, S.; Vassar, R.; Segal, D.; Gazit, E.; Frenkel, D.; et al. Orally administrated cinnamon extract reduces beta-amyloid oligomerization and corrects cognitive impairment in Alzheimer’s disease animal models. PLoS ONE 2011, 6, e16564. [Google Scholar] [CrossRef] [Green Version]
- Jana, A.; Modi, K.K.; Roy, A.; Anderson, J.A.; van Breemen, R.B.; Pahan, K. Up-regulation of neurotrophic factors by cinnamon and its metabolite sodium benzoate: Therapeutic implications for neurodegenerative disorders. J. Neuroimmune Pharmacol. 2013, 8, 739–755. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yang, Y.; Yuan, P.; Yang, Y.; Chen, K.; Jia, Q.; Li, Y. Immunosuppressive Effects of A-Type Procyanidin Oligomers from Cinnamomum tamala. Evid. Based Complement. Altern. Med. 2014, 2014, 365258. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Wang, T.; Chen, L.; Yu, B.W.; Jia, Q.; Chen, K.X.; Fan, H.M.; Li, Y.M.; Wang, H.Y. Trimer procyanidin oligomers contribute to the protective effects of cinnamon extracts on pancreatic beta-cells in vitro. Acta Pharmacol. Sin. 2016, 37, 1083–1090. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Xu, J.; Lv, Y.; He, P.; Liu, C.; Jiao, J.; Li, S.; Mao, X.; Xue, X. Proanthocyanidins exert a neuroprotective effect via ROS/JNK signaling in MPTPinduced Parkinson’s disease models in vitro and in vivo. Mol. Med. Rep. 2018, 18, 4913–4921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Gao, S.S.; Yang, H.J.; Wang, M.; Cheng, B.F.; Feng, Z.W.; Wang, L. Neuroprotective Effects of Proanthocyanidins, Natural Flavonoids Derived From Plants, on Rotenone-Induced Oxidative Stress and Apoptotic Cell Death in Human Neuroblastoma SH-SY5Y Cells. Front. Neurosci. 2018, 12, 369. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Xin, C.; Zhang, C.W.; Lim, K.L.; Zhang, H.; Fu, Z.; Li, L.; Huang, W. Natural Molecules From Chinese Herbs Protecting Against Parkinson’s Disease via Anti-oxidative Stress. Front. Aging Neurosci. 2018, 10, 246. [Google Scholar] [CrossRef]
- Strathearn, K.E.; Yousef, G.G.; Grace, M.H.; Roy, S.L.; Tambe, M.A.; Ferruzzi, M.G.; Wu, Q.L.; Simon, J.E.; Lila, M.A.; Rochet, J.C. Neuroprotective effects of anthocyanin- and proanthocyanidin-rich extracts in cellular models of Parkinsons disease. Brain Res. 2014, 1555, 60–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blade, C.; Aragones, G.; Arola-Arnal, A.; Muguerza, B.; Bravo, F.I.; Salvado, M.J.; Arola, L.; Suarez, M. Proanthocyanidins in health and disease. BioFactors 2016, 42, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Neilson, A.P.; O’Keefe, S.F.; Bolling, B.W. High-Molecular-Weight Proanthocyanidins in Foods: Overcoming Analytical Challenges in Pursuit of Novel Dietary Bioactive Components. Annu. Rev. Food Sci. Technol. 2016, 7, 43–64. [Google Scholar] [CrossRef] [PubMed]
- Bitzer, Z.T.; Glisan, S.L.; Dorenkott, M.R.; Goodrich, K.M.; Ye, L.; O’Keefe, S.F.; Lambert, J.D.; Neilson, A.P. Cocoa procyanidins with different degrees of polymerization possess distinct activities in models of colonic inflammation. J. Nutr. Biochem. 2015, 26, 827–831. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.A.; Kim, Y.J.; Cho, E.J.; Yokozawa, T. Ameliorative effects of proanthocyanidin on oxidative stress and inflammation in streptozotocin-induced diabetic rats. J. Agric. Food Chem. 2007, 55, 9395–9400. [Google Scholar] [CrossRef]
- Serra, A.; Macia, A.; Romero, M.P.; Valls, J.; Blade, C.; Arola, L.; Motilva, M.J. Bioavailability of procyanidin dimers and trimers and matrix food effects in in vitro and in vivo models. Br. J. Nutr. 2010, 103, 944–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Yamashita, Y.; Komeda, S.; Saito, A.; Ashida, H. Absorption, metabolism, distribution and faecal excretion of B-type procyanidin oligomers in mice after a single oral administration of black soybean seed coat extract. Food Funct. 2018, 9, 5362–5370. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Wang, S.; Wei, L.; Cui, Y.Y.; Chen, Y.H. Proanthocyanidins: Components, Pharmacokinetics and Biomedical Properties. Am. J. Chin. Med. 2020, 48, 813–869. [Google Scholar] [CrossRef]
- Serra, A.; Macia, A.; Rubio, L.; Angles, N.; Ortega, N.; Morello, J.R.; Romero, M.P.; Motilva, M.J. Distribution of procyanidins and their metabolites in rat plasma and tissues in relation to ingestion of procyanidin-enriched or procyanidin-rich cocoa creams. Eur. J. Nutr. 2013, 52, 1029–1038. [Google Scholar] [CrossRef]
- Serra, A.; Macia, A.; Romero, M.P.; Angles, N.; Morello, J.R.; Motilva, M.J. Distribution of procyanidins and their metabolites in rat plasma and tissues after an acute intake of hazelnut extract. Food Funct. 2011, 2, 562–568. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, Z.; Zhu, B.; Wang, G.; Jia, Q.; Li, Y.; Wu, X. A-Type Cinnamon Procyanidin Oligomers Protect Against 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Induced Neurotoxicity in Mice Through Inhibiting the P38 Mitogen-Activated Protein Kinase/P53/BCL-2 Associated X Protein Signaling Pathway. J. Nutr. 2020, 150, 1731–1737. [Google Scholar] [CrossRef]
- Catalkaya, G.; Venema, K.; Lucini, L.; Rocchetti, G.; Delmas, D.; Daglia, M.; Filippis, A.D.; Xiao, H.; Quiles, J.L.; Xiao, J.; et al. Interaction of dietary polyphenols and gut microbiota: Microbial metabolism of polyphenols, influence on the gut microbiota, and implications on host health. Food Front. 2020, 1, 109–133. [Google Scholar] [CrossRef]
- Hashimoto, R.; Yu, J.; Koizumi, H.; Ouchi, Y.; Okabe, T. Ginsenoside Rb1 Prevents MPP(+)-Induced Apoptosis in PC12 Cells by Stimulating Estrogen Receptors with Consequent Activation of ERK1/2, Akt and Inhibition of SAPK/JNK, p38 MAPK. Evid. Based Complement. Altern. Med. 2012, 2012, 693717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pariyar, R.; Lamichhane, R.; Jung, H.J.; Kim, S.Y.; Seo, J. Sulfuretin Attenuates MPP(+)-Induced Neurotoxicity through Akt/GSK3beta and ERK Signaling Pathways. Int. J. Mol. Sci. 2017, 18, 2753. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Sun, P.; Wang, T.; Chen, K.; Jia, Q.; Wang, H.; Li, Y. Diverse mechanisms of antidiabetic effects of the different procyanidin oligomer types of two different cinnamon species on db/db mice. J. Agric. Food Chem. 2012, 60, 9144–9150. [Google Scholar] [CrossRef] [PubMed]
- Janhom, P.; Dharmasaroja, P. Neuroprotective Effects of Alpha-Mangostin on MPP(+)-Induced Apoptotic Cell Death in Neuroblastoma SH-SY5Y Cells. J. Toxicol. 2015, 2015, 919058. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose | Cell Viability (%) |
|---|---|
| MPP+ (mM) | |
| 0 | 100 ± 0.6 |
| 0.1 | 93 ± 1.4 *** |
| 0.5 | 87 ± 1.3 *** |
| 1 | 72 ± 2.5 *** |
| 2.5 | 57 ± 1.7 *** |
| 5 | 45 ± 1.9 *** |
| 10 | 34 ± 0.2 *** |
| CPO-B (µM) | |
| 0 | 100 ± 2.0 |
| 5 | 94 ± 4.8 |
| 10 | 91 ± 6.1 |
| 25 | 83 ± 2.9 |
| 50 | 70 ± 2.5 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Q.; Chen, Z.; Zhu, B.; Li, Y.; Reddy, M.B.; Liu, H.; Dang, G.; Jia, Q.; Wu, X. Neuroprotective Effects of B-Type Cinnamon Procyanidin Oligomers on MPP+-Induced Apoptosis in a Cell Culture Model of Parkinson’s Disease. Molecules 2021, 26, 6422. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216422
Xu Q, Chen Z, Zhu B, Li Y, Reddy MB, Liu H, Dang G, Jia Q, Wu X. Neuroprotective Effects of B-Type Cinnamon Procyanidin Oligomers on MPP+-Induced Apoptosis in a Cell Culture Model of Parkinson’s Disease. Molecules. 2021; 26(21):6422. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216422
Chicago/Turabian StyleXu, Qi, Ziyu Chen, Borong Zhu, Yiming Li, Manju B. Reddy, Huilin Liu, Guodong Dang, Qi Jia, and Xiaojun Wu. 2021. "Neuroprotective Effects of B-Type Cinnamon Procyanidin Oligomers on MPP+-Induced Apoptosis in a Cell Culture Model of Parkinson’s Disease" Molecules 26, no. 21: 6422. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216422