
Physiological and Biochemical Parameters of Common Duckweed Lemna minor after the Exposure to Tetracycline and the Recovery from This Stress

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
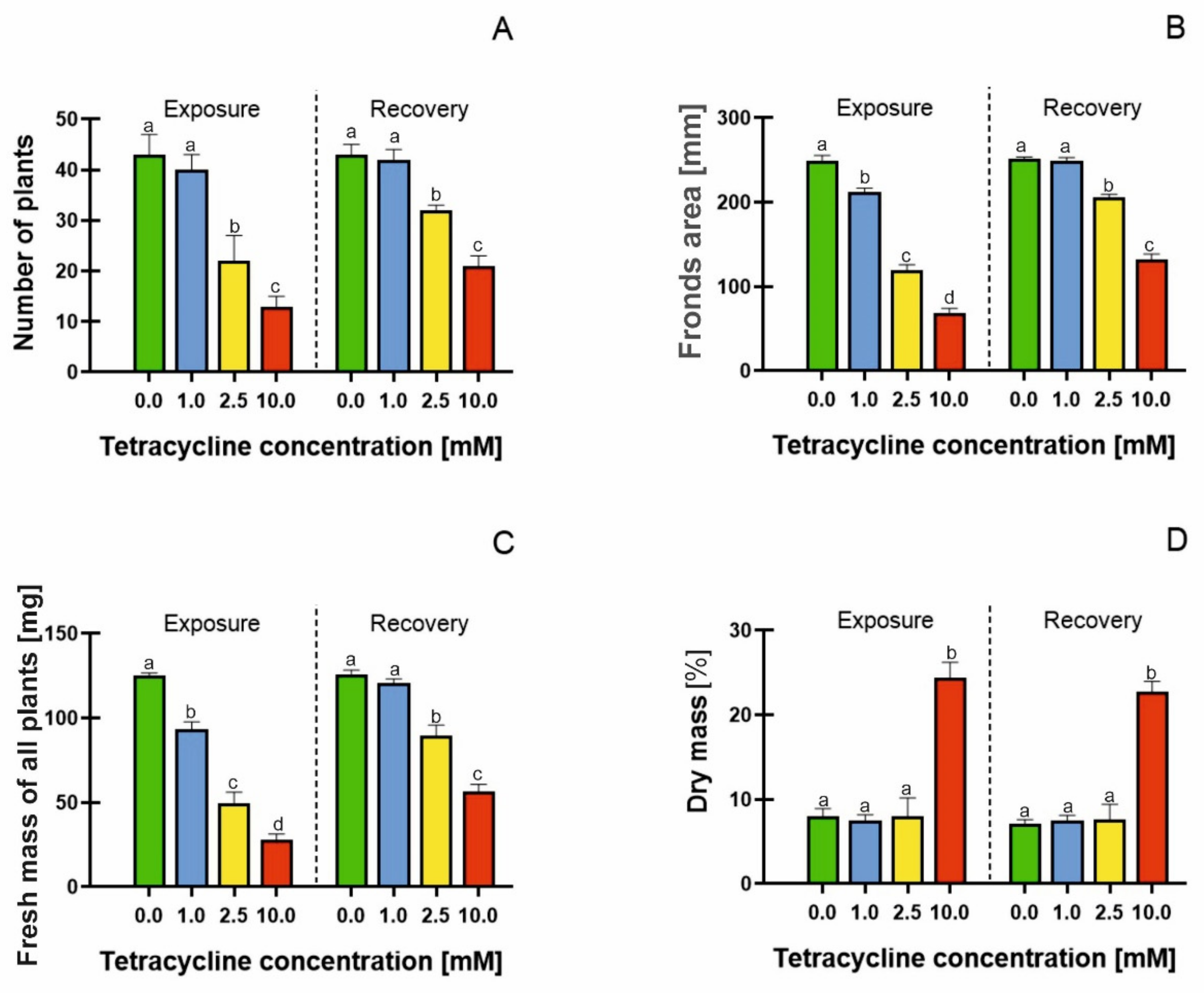
2.1. Growth Parameters
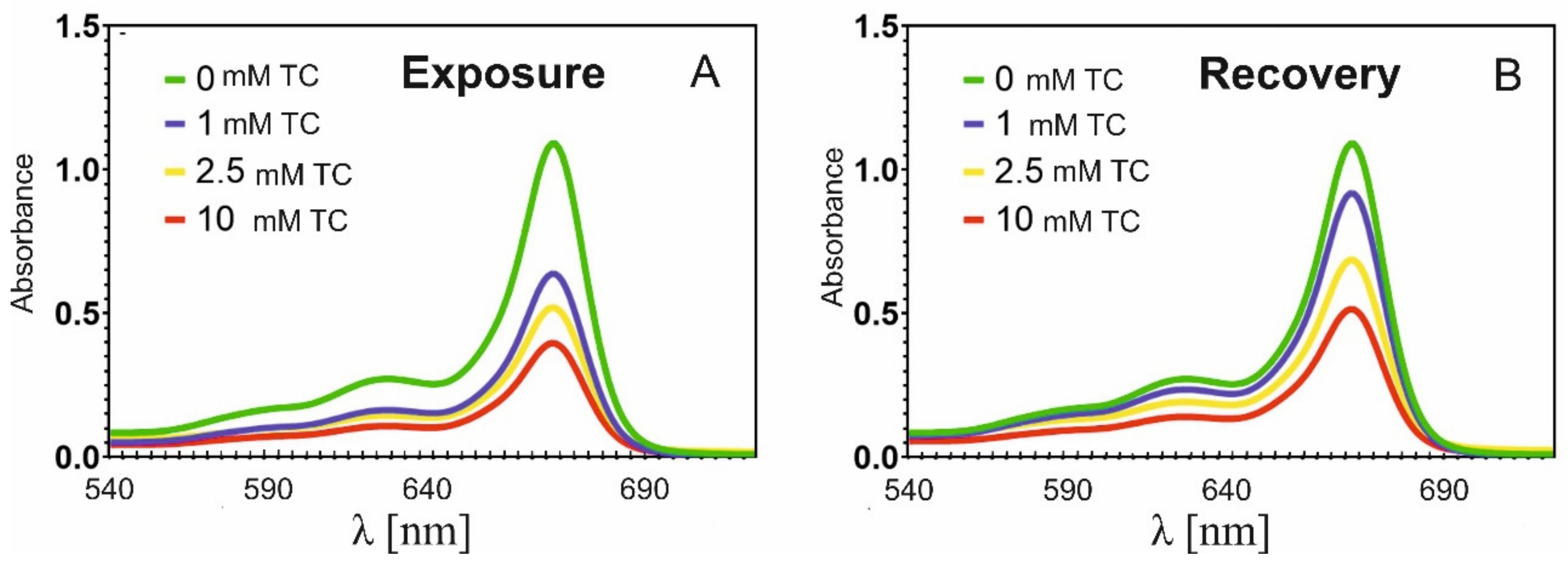
2.2. Effect of Tetracycline on Chlorophyll Content
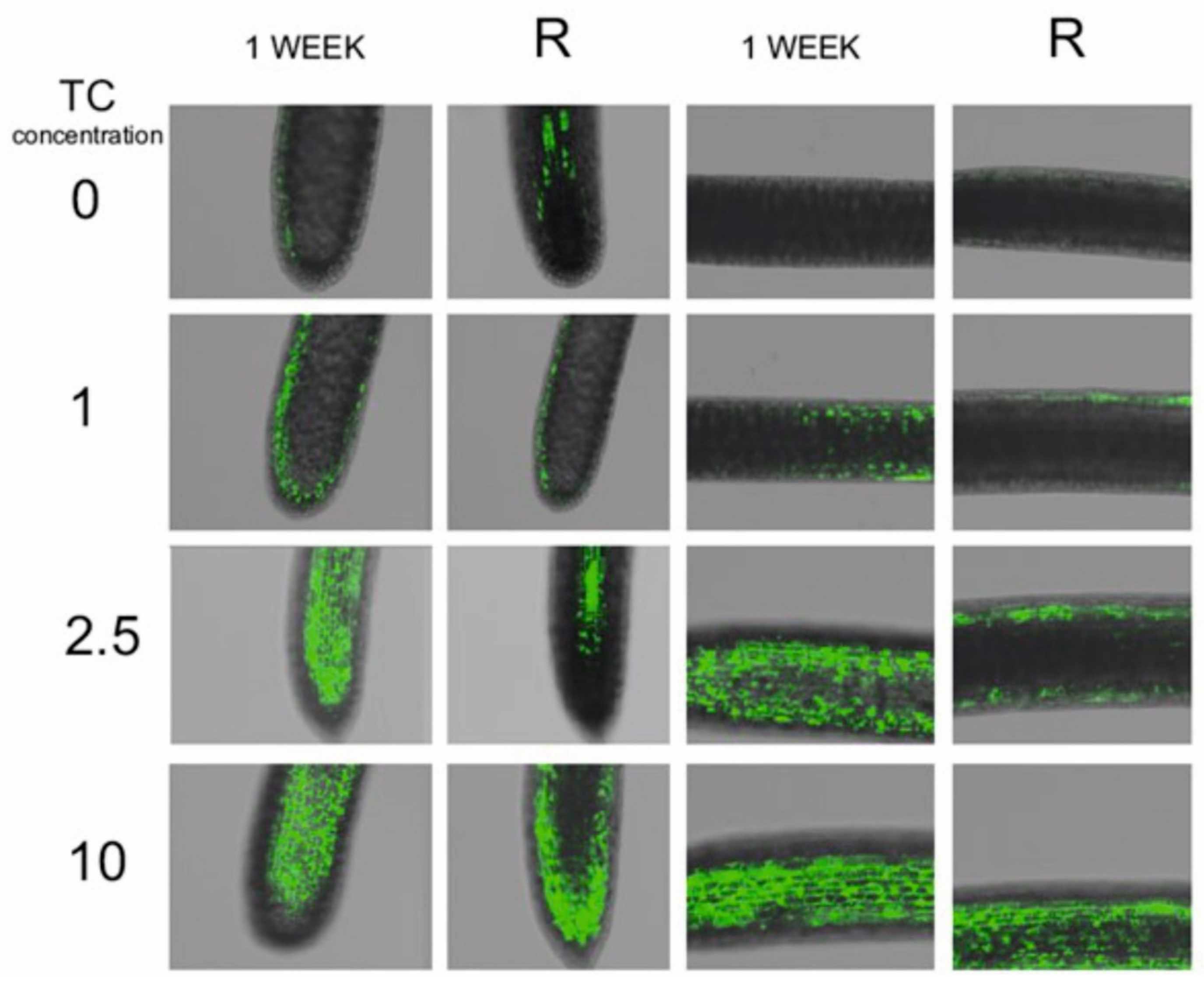
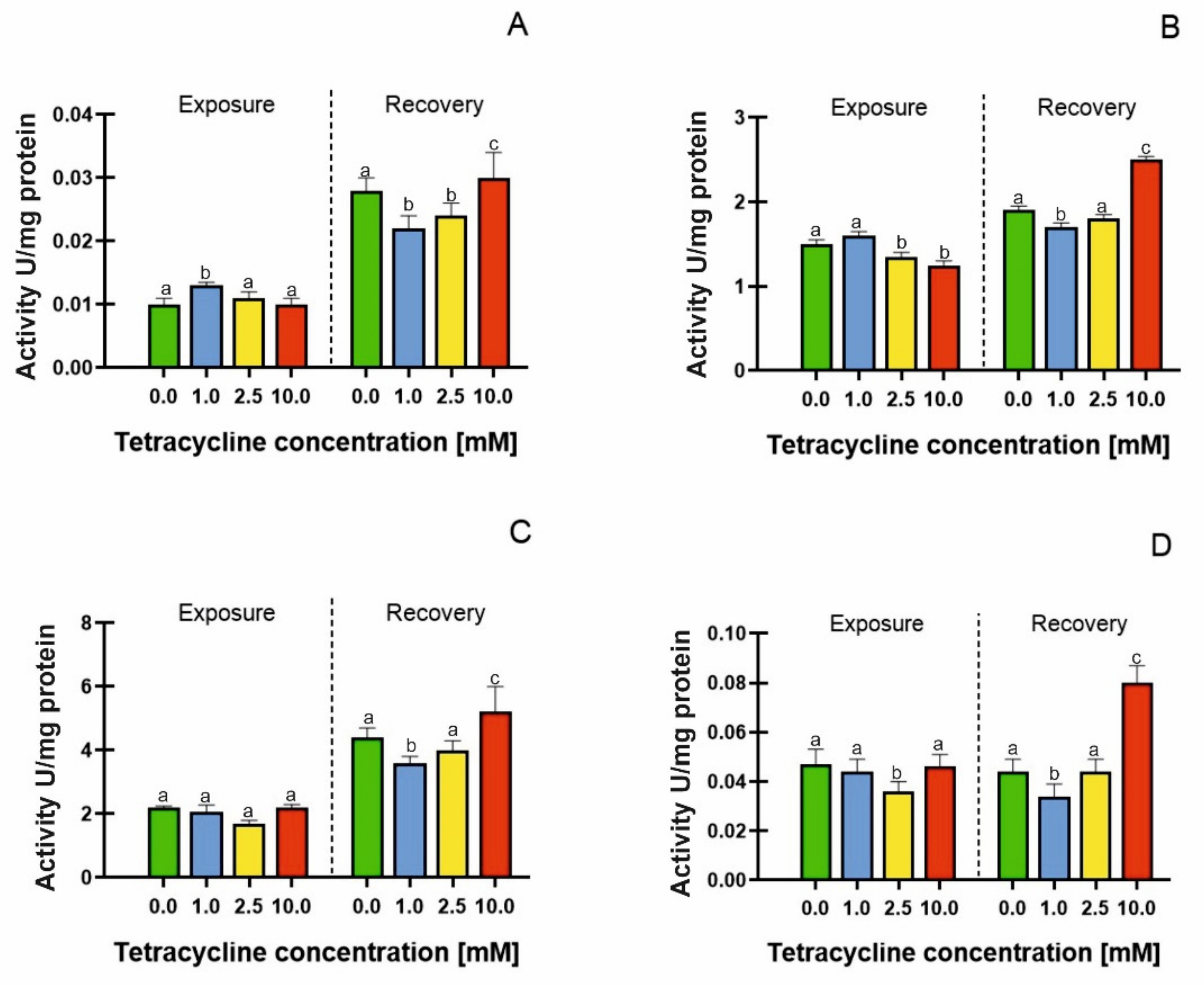
2.3. Free Radical Accumulation and Antioxidant Enzyme Activities
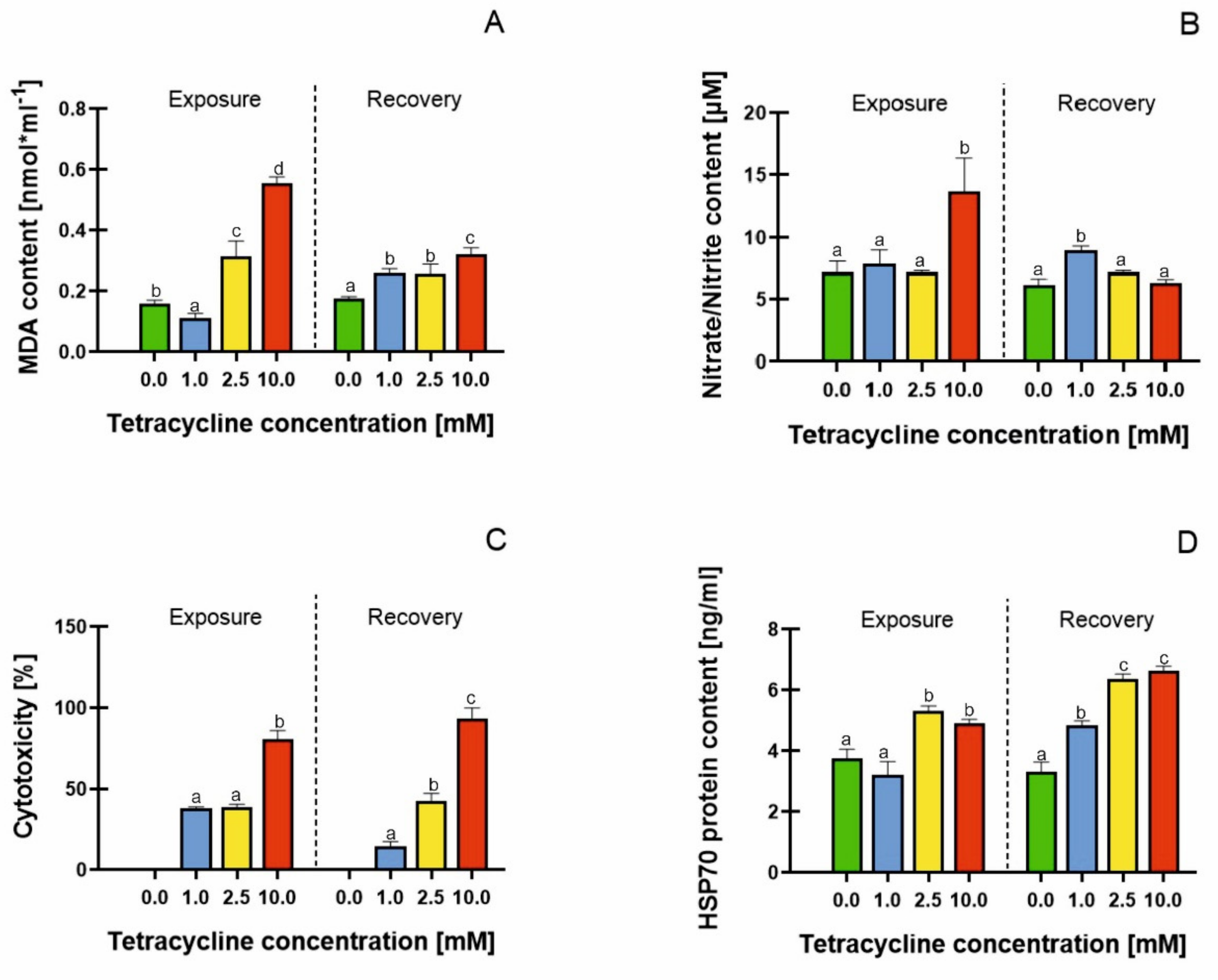
2.4. Malondialdehyde (MDA) Content, Cell Membrane Damage, Mitochondrial Activity and HSP70 Protein Content
3. Materials and Methods
3.1. Plant Material and Culture Conditions
3.2. Analyses of Growth, Frond Area, Dry and Fresh Weight
3.3. Isolation and Absorption Measurements of Chlorophyll
3.4. Presence of Free Radicals (ROS)
3.5. Enzyme Activity
3.6. Lipid Peroxidation—TBARS Assay
3.7. Assessment of Damages to Cell Membranes
3.8. Assessment of Mitochondrial Damages—WST-1 Test
3.9. Protein Isolation and HSP70 Protein Content
3.10. Statistics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- UNICEF. Water, Sanitation and Hygiene. Available online: www.unicef.org/wash (accessed on 23 May 2020).
- Murata, K.; Weihe, P.; Budtz-Jørgensen, E.; Jørgensen, P.J.; Grandjean, P. Delayed brainstem auditory evoked potential latencies in 14-year-old children exposed to methylmercury. J. Pediatr. 2004, 144, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. 2009. Available online: https://www.voanews.com/archive/who-waterborne-disease-worlds-leading-killer (accessed on 14 January 2021).
- Sengupta, B.; Trivedi, C.; Tandon, K.B.; Sinha, A.K.; Nazimuddin, M.; Swami, R. Evaluation of Operation and Maintenance of Sewage Treatment Plants in India-2007: Control of Urban Pollution Series: CUPS/68/2007; 1–67. 2008. Available online: http://admin.indiaenvironmentportal.org.in/files/sewage.pdf (accessed on 14 January 2021).
- Daud, M.K.; Nafees, M.; Ali, S.; Rizwan, M.; Bajwa, R.A.; Shakoor, M.B.; Arshad, M.U.; Chatha, S.A.S.; Deeba, F.; Murad, W.; et al. Drinking Water Quality Status and Contamination in Pakistan. BioMed. Res. Int. 2017, 2017, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Fact Sheet. Congress National Water Quality Inventory: Report to Congress (Report); United States Environmental Protection Agency (EPA): Washington, DC, USA, January 2017; EPA 841-R-16-011. Available online: https://www.epa.gov/sites/default/files/2017-12/documents/305brtc_finalowow_08302017.pdf (accessed on 15 January 2021).
- Qiao, Y.; Tao, J.; Chen, C.-H.; Qiu, J.; Tian, Y.; Hong, X.; Wu, J. A Miniature On-Chip Methane Sensor Based on an Ultra-Low Loss Waveguide and a Micro-Ring Resonator Filter. Micromachines 2017, 8, 160. [Google Scholar] [CrossRef] [Green Version]
- Sriram, A.; Kalanxhi, E.; Kapoor, G.; Craig, J.; Balasubramanian, R.; Brar, S.; Criscuolo, N.; Hamilton, A.; Klein, E.; Tseng, K.; et al. State of the World’s Antibiotics 2021: A Global Analysis of Antimicrobial Resistance and Its Drivers. Center for Disease Dynamics, Economics & Policy. 2021. Available online: https://www.anpario.com/wp-content/uploads/2021/03/cddep-antibiotics-2021.pdf (accessed on 20 January 2021).
- Kümmerer, K. The presence of pharmaceuticals in the environment due to human use—Present knowledge and future challenges. J. Environ. Manag. 2009, 90, 2354–2366. [Google Scholar] [CrossRef] [PubMed]
- EMA. Sales of Veterinary Antimicrobial Agents in 19 EU/EEA Countries in 2010: Second ESVAC Report, 74. 2012. Available online: https://0-doi-org.brum.beds.ac.uk/10.2809/676974 (accessed on 22 January 2021).
- EMEA. The European Agency for the Evaluation of Medicinal Products. Oxytetracycline, Tetracycline, Chlortetracycline. Summary Report (3); EMEA/MRL/023/95; EMEA: London, UK, 1995; Available online: https://www.ema.europa.eu/en/documents/mrl-report/oxytetracycline-tetracycline-chlortetracycline-summary-report-3-committee-veterinary-medicinal_en.pdf (accessed on 22 January 2021).
- Michalova, E.; Novotna, P.; Schlegelova, J. Tetracyclines in veterinary medicine and bacterial resistance to them. Vet. Med. 2004, 49, 79–100. [Google Scholar] [CrossRef] [Green Version]
- Agwuh, K.N. Pharmacokinetics and pharmacodynamics of the tetracyclines including glycylcyclines. J. Antimicrob. Chemother. 2006, 58, 256–265. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhang, H.; Luo, Y.; Song, J. Occurrence and assessment of veterinary antibiotics in swine manures: A case study in East China. Chin. Sci. Bull. 2012, 57, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Burke, V.; Richter, D.; Greskowiak, J.; Mehrtens, A.; Schulz, L.; Massmann, G. Occurrence of Antibiotics in Surface and Groundwater of a Drinking Water Catchment Area in Germany. Water Environ. Res. 2016, 88, 652–659. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, M.; Guo, C.; An, D.; Xu, J.; Zhang, Y.; Xi, B. Distribution and ecological risk of antibiotics in a typical effluent–receiving river (Wangyang River) in north China. Chemosphere 2014, 112, 267–274. [Google Scholar] [CrossRef]
- Borghi, A.A.; Palma, M.S.A. Tetracycline: Production, waste treatment and environmental impact assessment. Braz. J. Pharm. Sci. 2014, 50, 25–40. [Google Scholar] [CrossRef]
- Tang, J.; Shi, T.; Wu, X.; Cao, H.; Li, X.; Hua, R.; Tang, F.; Yue, Y. The occurrence and distribution of antibiotics in Lake Chaohu, China: Seasonal variation, potential source and risk assessment. Chemosphere 2015, 122, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Matsui, Y.; Ozu, T.; Inoue, T.; Matsushita, T. Occurrence of a veterinary antibiotic in streams in a small catchment area with livestock farms. Desalination 2002, 226, 215–221. [Google Scholar] [CrossRef]
- Kivits, T.; Broers, H.P.; Beeltje, H.; van Vliet, M.; Griffioen, J. Presence and fate of veterinary antibiotics in age-dated groundwater in areas with intensive livestock farming. Environ. Pollut. 2018, 241, 988–998. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Q.; Zhang, J.; Dong, J.; Yan, H.; Chen, C.; Feng, R. Characterization and source identification of tetracycline antibiotics in the drinking water sources of the lower Yangtze River. J. Environ. Manag. 2019, 244, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zhang, G.; Li, X.; Zou, S.; Li, P.; Hu, Z.; Li, J. Occurrence and elimination of antibiotics at four sewage treatment plants in the Pearl River Delta (PRD), South China. Water Res. 2007, 41, 4526–4534. [Google Scholar] [CrossRef]
- Rydzyński, D.; Piotrowicz-Cieślak, A.I.; Grajek, H.; Wasilewski, J. Investigation of chlorophyll degradation by tetracycline. Chemosphere 2019, 229, 409–417. [Google Scholar] [CrossRef]
- Gomes, M.P.; de Brito, J.C.M.; Rocha, D.C.; Navarro-Silva, M.A.; Juneau, P. Individual and combined effects of amoxicillin, enrofloxacin, and oxytetracycline on Lemna minor physiology. Ecotoxicol. Environ. Saf. 2020, 203, 111025. [Google Scholar] [CrossRef]
- Liu, X.-H.; Lv, Y.; Xu, K.; Xiao, X.; Xi, B.; Lu, S. Response of ginger growth to a tetracycline-contaminated environment and residues of antibiotic and antibiotic resistance genes. Chemosphere 2018, 201, 137–143. [Google Scholar] [CrossRef]
- Ali, Z.; Waheed, H.; Kazi, A.G.; Hayat, A.; Ahmad, M. Chapter 16—Duckweed: An efficient hyperaccumulator of heavy metals in water bodies. In Plant Metal Interaction; Ahmad, P., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 411–429. ISBN 9780128031582. [Google Scholar] [CrossRef]
- Devlamynck, R.; de Souza, M.F.; Bog, M.; Leenknegt, J.; Eeckhout, M.; Meers, E. Effect of the growth medium composition on nitrate accumulation in the novel protein crop Lemna minor. Ecotoxicol. Environ. Saf. 2020, 206, 111380. [Google Scholar] [CrossRef]
- Paterson, J.B.; Camargo-Valero, M.A.; Baker, A. Uncoupling growth from phosphorus uptake in Lemna: Implications for use of duckweed in wastewater remediation and P recovery in temperate climates. Food Energy Secur. 2020, 9, 244. [Google Scholar] [CrossRef]
- Baciak, M.; Sikorski, Ł.; Piotrowicz-Cieślak, A.I.; Adomas, B. Content of biogenic amines in Lemna minor (common duckweed) growing in medium contaminated with tetracycline. Aquat. Toxicol. 2016, 180, 95–102. [Google Scholar] [CrossRef]
- OECD. Lemna sp. growth inhibition test. In OECD Guidelines for the Testing of Chemicals Section 2—Effects on Biotic Systems; OECD Publishing: Paris, France, 2006; p. 22. [Google Scholar] [CrossRef]
- Ge, X.; Zhang, N.; Phillips, G.C.; Xu, J. Growing Lemna minor in agricultural wastewater and converting the duckweed biomass to ethanol. Bioresour. Technol. 2012, 124, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Goopy, J.P.; Murray, P. A Review on the Role of Duckweed in Nutrient Reclamation and as a Source of Animal Feed. Asian-Australas. J. Anim. Sci. 2003, 16, 297–305. [Google Scholar] [CrossRef]
- Toyama, T.; Hanaoka, T.; Tanaka, Y.; Morikawa, M.; Mori, K. Comprehensive evaluation of nitrogen removal rate and biomass, ethanol, and methane production yields by combination of four major duckweeds and three types of wastewater effluent. Bioresour. Technol. 2018, 250, 464–473. [Google Scholar] [CrossRef]
- Appenroth, K.-J.; Sree, K.S.; Bog, M.; Ecker, J.; Seeliger, C.; Böhm, V.; Lorkowski, S.; Sommer, K.; Vetter, W.; Tolzin-Banasch, K.; et al. Nutritional Value of the Duckweed Species of the Genus Wolffia (Lemnaceae) as Human Food. Front. Chem. 2018, 6, 483. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Raju, R.; Mrad, M.; Reddell, P.; Münch, G. The reciprocal EC50 value as a convenient measure of the potency of a compound in bioactivity-guided purification of natural products. Fitoterapia 2020, 143, 104598. [Google Scholar] [CrossRef]
- Pomati, F.; Netting, A.G.; Calamari, D.; Neilan, B.A. Effects of erythromycin, tetracycline and ibuprofen on the growth of Synechocystis sp. and Lemna minor. Aquat. Toxicol. 2004, 67, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Brain, R.A.; Johnson, D.J.; Richards, S.M.; Hanson, M.L.; Sanderson, H.; Lam, M.W.; Young, C.; Mabury, S.A.; Sibley, P.; Solomon, K.R. Microcosm evaluation of the effects of an eight pharmaceutical mixture to the aquatic macrophytes Lemna gibba and Myriophyllum sibiricum. Aquat. Toxicol. 2004, 70, 23–40. [Google Scholar] [CrossRef]
- Hussain, S.; Naeem, M.; Chaudhry, M.N.; Iqbal, M.A. Accumulation of Residual Antibiotics in the Vegetables Irrigated by Pharmaceutical Wastewater. Expo. Health 2015, 8, 107–115. [Google Scholar] [CrossRef]
- Margas, M.; Piotrowicz-Cieślak, A.I.; Ziółkowska, A.; Adomas, B. Tetracycline Accumulation in Pea Seedlings and its Effects on Proteome and Enzyme Activities. Int. J. Agric. Biol. 2016, 18, 789–796. [Google Scholar] [CrossRef]
- Di Marco, G.; Gismondi, A.; Canuti, L.; Scimeca, M.; Volpe, A.; Canini, A. Tetracycline accumulates in Iberis sempervirens L. through apoplastic transport inducing oxidative stress and growth inhibition. Plant Biol. 2013, 16, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Rydzyński, D.; Piotrowicz-Cieślak, A.I.; Grajek, H.; Michalczyk, D. Instability of chlorophyll in yellow lupin seedlings grown in soil contaminated with ciprofloxacin and tetracycline. Chemosphere 2017, 184, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Piotrowicz-Cieślak, A.I.; Adomas, B.; Nałęcz-Jawecki, G.; Michalczyk, D.J. Phytotoxicity of Sulfamethazine Soil Pollutant to Six Legume Plant Species. J. Toxicol. Environ. Health Part A 2010, 73, 1220–1229. [Google Scholar] [CrossRef]
- Michelini, L.; Reichel, R.; Werner, W.; Ghisi, R.; Thiele-Bruhn, S. Sulfadiazine uptake and effects on Salix fragilis L. and Zea mays L. plants. Air. Soil Pollut. 2012, 223, 5243–5257. [Google Scholar] [CrossRef]
- Schopfer, P. Hydrogen peroxide-mediated cell-wall stiffening in vitro in maize coleoptiles. Planta 1996, 199, 43–49. [Google Scholar] [CrossRef]
- Zaltauskaite, J.; Kaciene, G. Lemna minor recovery potential after short-term exposure to sulfonylurea herbicide. Acta Physiol. Plant. 2020, 42, 35. [Google Scholar] [CrossRef]
- Seely, G.; Jensen, R. Effect of solvent on the spectrum of chlorophyll. Spectrochim. Acta 1965, 21, 1835–1845. [Google Scholar] [CrossRef]
- Margas, M.; Piotrowicz-Cieślak, A.I.; Michalczyk, D.; Głowacka, K. A Strong Impact of Soil Tetracycline on Physiology and Biochemistry of Pea Seedlings. Scientifica 2019, 2019, 1–14. [Google Scholar] [CrossRef] [Green Version]
- LeBel, C.P.; Ischiropoulos, H.; Bondy, S.C. Evaluation of the probe 2′,7′-dichlorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chem. Res. Toxicol. 1992, 5, 227–231. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, J.; Vytášek, R.; Uhlik, J.; Vajner, L. Oxidative Stress in the Developing Rat Brain due to Production of Reactive Oxygen and Nitrogen Species. Oxidative Med. Cell. Longev. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tarpey, M.M.; Fridovich, I. Methods of Detection of Vascular Reactive Species. Circ. Res. 2001, 89, 224–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majer, P.; Vidović, M.; Czégény, G.; Jovanović, S.V.; Strid, Å.; Hideg, É. Evaluation of procedures for assessing anti- and pro-oxidants in plant samples. Anal. Methods 2016, 8, 5569–5580. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; He, Z.; Chen, N.; Tang, Z.; Wang, Q.; Cai, Y. The Roles of Environmental Factors in Regulation of Oxidative Stress in Plant. BioMed. Res. Int. 2019, 2019, 1–11. [Google Scholar] [CrossRef]
- Orzoł, A.; Piotrowicz-Cieślak, A.I. Levofloxacin is phytotoxic and modifies the protein profile of lupin seedlings. Environ. Sci. Pollut. Res. 2017, 24, 22226–22240. [Google Scholar] [CrossRef] [Green Version]
- Tsukagoshi, H. Defective root growth triggered by oxidative stress is controlled through the expression of cell cycle-related genes. Plant Sci. 2012, 197, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Tsukagoshi, H. Control of root growth and development by reactive oxygen species. Curr. Opin. Plant Biol. 2016, 29, 57–63. [Google Scholar] [CrossRef]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate Peroxidase and Catalase Activities and Their Genetic Regulation in Plants Subjected to Drought and Salinity Stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef] [Green Version]
- Drobniewska, A.; Wójcik, D.; Kapłan, M.; Adomas, B.; Piotrowicz-Cieślak, A.; Nałęcz-Jawecki, G. Recovery of Lemna minor after exposure to sulfadimethoxine irradiated and non-irradiated in a solar simulator. Environ. Sci. Pollut. Res. 2016, 24, 27642–27652. [Google Scholar] [CrossRef]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef]
- Nunes, B.; Veiga, V.; Frankenbach, S.; Serôdio, J.; Pinto, G. Evaluation of physiological changes induced by the fluoroquinolone antibiotic ciprofloxacin in the freshwater macrophyte species Lemna minor and Lemna gibba. Environ. Toxicol. Pharmacol. 2019, 72, 103242. [Google Scholar] [CrossRef]
- Corpas, F.J.; Leterrier, M.; Valderrama, R.; Airaki, M.; Chaki, M.; Palma, J.M.; Barroso, J.B. Nitric oxide imbalance provokes a nitrosative response in plants under abiotic stress. Plant Sci. 2011, 181, 604–611. [Google Scholar] [CrossRef]
- Valderrama, R.; Corpas, F.J.; Carreras, A.; Ocaña, A.M.F.; Chaki, M.; Luque, F.; Rodríguez, M.V.G.; Colmenero-Varea, P.; Del Río, L.A.; Barroso, J.B. Nitrosative stress in plants. FEBS Lett. 2007, 581, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Kummerová, M.; Zezulka, Š.; Babula, P.; Tříska, J. Possible ecological risk of two pharmaceuticals diclofenac and paracetamol demonstrated on a model plant Lemna minor. J. Hazard. Mater. 2016, 302, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, A.D. Nitric Oxide Signaling in Plants. Vitam. Horm. 2005, 72, 339–398. [Google Scholar] [CrossRef] [PubMed]
- Geisler, D.A.; Johansson, F.I.; Svensson, A.S.; Rasmusson, A.G. Antimycin A treatment decreases respiratory internal rotenone-insensitive NADH oxidation capacity in potato leaves. BMC Plant Biol. 2004, 4, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, M.P.; Gonçalves, C.A.; de Brito, J.C.M.; Souza, A.M.; Cruz, F.V.D.S.; Bicalho, E.M.; Figueredo, C.; Garcia, Q.S. Ciprofloxacin induces oxidative stress in duckweed (Lemna minor L.): Implications for energy metabolism and antibiotic-uptake ability. J. Hazard. Mater. 2017, 328, 140–149. [Google Scholar] [CrossRef]
- Rocha, D.C.; Rocha, C.D.S.; Tavares, D.S.; Calado, S.L.D.M.; Gomes, M.P. Veterinary antibiotics and plant physiology: An overview. Sci. Total. Environ. 2021, 767, 144902. [Google Scholar] [CrossRef] [PubMed]
- Usman, M.G.; Rafii, M.Y.; Martini, M.Y.; Yusuff, O.A.; Ismail, M.R.; Miah, G. Molecular analysis of Hsp70 mechanisms in plants and their function in response to stress. Biotechnol. Genet. Eng. Rev. 2017, 33, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Gorovits, R.; Sobol, I.; Akama, K.; Chefetz, B.; Czosnek, H. Pharmaceuticals in treated wastewater induce a stress response in tomato plants. Sci. Rep. 2020, 10, 1856. [Google Scholar] [CrossRef] [PubMed]
- Ziółkowska, A.; Piotrowicz-Cieślak, A.I. Sparfloxacin—A potential contaminant of organically grown plants? Appl. Ecol. Environ. Res. 2016, 14, 29–44. [Google Scholar] [CrossRef]
- Das, A.; Eldakak, M.; Paudel, B.; Kim, D.-W.; Hemmati, H.; Basu, C.; Rohila, J.S. Leaf Proteome Analysis Reveals Prospective Drought and Heat Stress Response Mechanisms in Soybean. BioMed. Res. Int. 2016, 2016, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Rydzyński, D.; Piotrowicz-Cieślak, A.I.; Grajek, H.; Michalczyk, D. Chlorophyll degradation by tetracycline and cadmium in spinach (Spinacia oleracea L.) leaves. Int. J. Environ. Sci. Technol. 2018, 16, 6301–6314. [Google Scholar] [CrossRef] [Green Version]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Murshed, R.; Lopez-Lauri, F.; Keller, C.; Monnet, F.; Sallanon, H. Acclimation to drought stress enhances oxidative stress tolerance in Solanum lycopersicum L. fruits. Plant Stress 2008, 2, 145–151. [Google Scholar]
- Chance, B.; Maehly, A.C. Methods in Enzymology, II; Elsevier: Amsterdam, The Netherlands, 1955; pp. 773–775. [Google Scholar]
- Radić, S.; Radić-Stojković, M.; Pevalek-Kozlina, B. Influence of NaCl and mannitol on peroxidase activity and lipid peroxidation in Centaurea ragusina L. roots and shoots. J. Plant Physiol. 2005, 163, 1284–1292. [Google Scholar] [CrossRef] [PubMed]
- Murshed, R.; Lopez-Lauri, F.; Sallanon, H. Microplate quantification of enzymes of the plant ascorbate–glutathione cycle. Anal. Biochem. 2008, 383, 320–322. [Google Scholar] [CrossRef] [PubMed]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Jett, M.; Seed, T.M.; Jamieson, G.A. Isolation and characterization of plasma membranes and intact nuclei from lymphoid cells. J. Biol. Chem. 1977, 252, 2134–2142. [Google Scholar] [CrossRef]
- Heckman, N.L.; Elthon, T.E.; Horst, G.L.; Gaussoin, R.E. Influence of Trinexapac-Ethyl on Respiration of Isolated Wheat Mitochondria. Crop. Sci. 2002, 42, 423–427. [Google Scholar] [CrossRef]
- Isaacson, T.; Damasceno, C.M.B.; Saravanan, R.S.; He, Y.; Catalá, C.; Saladié, M.; Rose, J.K.C. Sample extraction techniques for enhanced proteomic analysis of plant tissues. Nat. Protoc. 2006, 1, 769–774. [Google Scholar] [CrossRef]
- Böger, B.; Surek, M.; de O Vilhena, R.; Fachi, M.M.; Junkert, A.M.; Santos, J.M.; Pontarolo, R. Occurrence of antibiotics and antibiotic resistant bacteria in subtropical urban rivers in Brazil. J. Hazard. Mater. 2020, 402, 123448. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.J.; Paíga, P.; Silva, A.; Llaguno, C.P.; Carvalho, M.; Vázquez, F.M.; Delerue-Matos, C. Antibiotics and antidepressants occurrence in surface waters and sediments collected in the north of Portugal. Chemosphere 2019, 287, 124729. [Google Scholar] [CrossRef] [PubMed]
- Giebułtowicz, J.; Tyski, S.; Wolinowska, R.; Grzybowska, W.; Zaręba, T.; Drobniewska, A.; Nałęcz-Jawecki, G. Occurrence of antimicrobial agents, drug-resistant bacteria, and genes in the sewage-impacted Vistula River (Poland). Environ. Sci. Pollut. Res. 2017, 25, 5788–5807. [Google Scholar] [CrossRef]
- López-Serna, R.; Petrović, M.; Barceló, D. Development of a fast instrumental method for the analysis of pharmaceuticals in environmental and wastewaters based on ultra high performance liquid chromatography (UHPLC)–tandem mass spectrometry (MS/MS). Chemosphere 2011, 85, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Y.; Guo, X.; Hu, J. Occurrence and risks of antibiotics in an urban river in northeastern Tibetan Plateau. Sci. Rep. 2020, 10, 20054. [Google Scholar] [CrossRef]
- Fonseca, E.; Hernández, F.; Ibáñez, M.; Rico, A.; Pitarch, E.; Bijlsma, L. Occurrence and ecological risks of pharmaceuticals in a Mediterranean river in Eastern Spain. Environ. Int. 2020, 144, 106004. [Google Scholar] [CrossRef]
- Jiang, L.; Hu, X.; Yin, D.; Zhang, H.; Yu, Z. Occurrence, distribution and seasonal variation of antibiotics in the Huangpu River, Shanghai, China. Chemosphere 2011, 82, 822–828. [Google Scholar] [CrossRef]
- Joshua, D.I.; Praveenkumarreddy, Y.; Prabhasankar, V.P.; D’Souza, A.P.; Yamashita, N.; Balakrishna, K. First report of pharmaceuticals and personal care products in two tropical rivers of southwestern India. Environ. Monit. Assess. 2020, 192, 529. [Google Scholar] [CrossRef]
- Yan, C.; Yang, Y.; Zhou, J.; Liu, M.; Nie, M.; Shi, H.; Gu, L. Antibiotics in the surface water of the Yangtze Estuary: Occurrence, distribution and risk assessment. Environ. Pollut. 2013, 175, 22–29. [Google Scholar] [CrossRef]
- Kolpin, D.W.; Furlong, E.T.; Meyer, M.T.; Thurman, E.M.; Zaugg, S.D.; Barber, L.B.; Buxton, H.T. Pharmaceuticals, Hormones, and Other Organic Wastewater Contaminants in U.S. Streams, 1999−2000: A National Reconnaissance. Environ. Sci. Technol. 2002, 36, 1202–1211. [Google Scholar] [CrossRef] [Green Version]
- Arsand, J.B.; Hoff, R.B.; Jank, L.; Bussamara, R.; Dallegrave, A.; Bento, F.M.; Pizzolato, T.M. Presence of antibiotic resistance genes and its association with antibiotic occurrence in Dilúvio River in southern Brazil. Sci. Total Environ. 2020, 738, 139781. [Google Scholar] [CrossRef]
- Kruć, R.; Dragon, K.; Górski, J. Migration of Pharmaceuticals from the Warta River to the Aquifer at a Riverbank Filtration Site in Krajkowo (Poland). Water 2019, 11, 2238. [Google Scholar] [CrossRef] [Green Version]
- Managaki, S.; Murata, A.; Takada, H.; Tuyen, B.C.; Chiem, N.H. Distribution of Macrolides, Sulfonamides, and Trimethoprim in Tropical Waters: Ubiquitous Occurrence of Veterinary Antibiotics in the Mekong Delta. Environ. Sci. Technol. 2007, 41, 8004–8010. [Google Scholar] [CrossRef]
- Tamtam, F.; Mercier, F.; Le Bot, B.; Eurin, J.; Tuc Dinh, Q.; Clément, M.; Chevreuil, M. Occurrence and fate of antibiotics in the Seine River in various hydrological conditions. Sci. Total Environ. 2008, 393, 84–95. [Google Scholar] [CrossRef]
- Feitosa-Felizzola, J.; Chiron, S. Occurrence and distribution of selected antibiotics in a small Mediterranean stream (Arc River, Southern France). J. Hydrol. 2009, 364, 50–57. [Google Scholar] [CrossRef]
- White, D.; Lapworth, D.J.; Civil, W.; Williams, P. Tracking changes in the occurrence and source of pharmaceuticals within the River Thames, UK; from source to sea. Environ. Pollut. 2019, 249, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Ngumba, E.; Gachanja, A.; Tuhkanen, T. Occurrence of selected antibiotics and antiretroviral drugs in Nairobi River Basin, Kenya. Sci. Total Environ. 2016, 539, 206–213. [Google Scholar] [CrossRef]
- Deng, W.J.; Li, N.; Ying, G.G. Antibiotic distribution, risk assessment, and microbial diversity in river water and sediment in Hong Kong. Environ. Geochem. Health 2018, 40, 2191–2203. [Google Scholar] [CrossRef]
- Tran, N.H.; Hoang, L.; Nghiem, L.D.; Nguyen, N.M.H.; Ngo, H.H.; Guo, W.; Gin, K.Y.H. Occurrence and risk assessment of multiple classes of antibiotics in urban canals and lakes in Hanoi, Vietnam. Sci. Total Environ. 2019, 692, 157–174. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lei, L.; Liu, S.; Han, J.; Li, R.; Men, L.L.; Wei, L.; Sheng, Y.; Yang, L.; Zhou, B.; et al. Occurrence and risk assessment of pharmaceuticals and personal care products (PPCPs) against COVID-19 in lakes and WWTP-river-estuary system in Wuhan, China. Sci. Total Environ. 2021, 792, 148352. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Lu, J.; Liu, Z.; Tong, Y.; Li, S. Concentration and distribution of antibiotics in water–sediment system of Bosten Lake, Xinjiang. Environ. Sci. Pollut. Res. 2014, 22, 1670–1678. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Y.; Zhou, C.; Guo, C.; Wang, D.; Du, P.; Meng, W. Distribution, sources and composition of antibiotics in sediment, overlying water and pore water from Taihu Lake, China. Sci. Total Environ. 2014, 497, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Wu, Y.; Zhang, W.; Zhong, J.; Lou, Q.; Yang, P.; Fang, Y. Occurrence, distribution, and risk assessment of antibiotics in the surface water of Poyang Lake, the largest freshwater lake in China. Chemosphere 2017, 184, 137–147. [Google Scholar] [CrossRef]
- Blair, B.D.; Crago, J.P.; Hedman, C.J.; Klaper, R.D. Pharmaceuticals and personal care products found in the Great Lakes above concentrations of environmental concern. Chemosphere 2013, 93, 2116–2123. [Google Scholar] [CrossRef] [Green Version]
- Park, M.; Reckhow, D.; Lavine, M.; Rosenfeldt, E.; Stanford, B.; Park, M.-H. Multivariate Analyses for Monitoring EDCs and PPCPs in a Lake Water. Water Environ. Res. 2014, 86, 2233–2241. [Google Scholar] [CrossRef]
- Li, H.; Helm, P.A.; Metcalfe, C.D. Sampling in the Great Lakes for pharmaceuticals, personal care products, and endocrine-disrupting substances using the passive polar organic chemical integrative sampler. Environ. Toxicol. Chem. 2010, 29, 751–762. [Google Scholar] [CrossRef]
- Minh, T.B.; Leung, H.W.; Loi, I.H.; Chan, W.H.; So, M.K.; Mao, J.Q.; Richardson, B.J. Antibiotics in the Hong Kong metropolitan area: Ubiquitous distribution and fate in Victoria Harbour. Mar. Pollut. Bull. 2009, 58, 1052–1062. [Google Scholar] [CrossRef]
- Du, J.; Zhao, H.; Liu, S.; Xie, H.; Wang, Y.; Chen, J. Antibiotics in the coastal water of the South Yellow Sea in China: Occurrence, distribution and ecological risks. Sci. Total Environ. 2017, 595, 521–552. [Google Scholar] [CrossRef]
- Tahrani, L.; Van Loco, J.; Ben Mansour, H.; Reyns, T. Occurrence of antibiotics in pharmaceutical industrial wastewater, wastewater treatment plant and sea waters in Tunisia. J. Water Health 2015, 14, 208–213. [Google Scholar] [CrossRef]
- Chen, L.; Lang, H.; Liu, F.; Jin, S.; Yan, T. Presence of Antibiotics in Shallow Groundwater in the Northern and Southwestern Regions of China. Groundwater 2017, 56, 451–457. [Google Scholar] [CrossRef]
- Rutgersson, C.; Fick, J.; Marathe, N.; Kristiansson, E.; Janzon, A.; Angelin, M.; Larsson, D.G.J. Fluoroquinolones and qnr Genes in Sediment, Water, Soil, and Human Fecal Flora in an Environment Polluted by Manufacturing Discharges. Environ. Sci. Technol. 2014, 48, 7825–7832. [Google Scholar] [CrossRef]
- Barnes, K.K.; Kolpin, D.W.; Furlong, E.T.; Zaugg, S.D.; Meyer, M.T.; Barber, L.B. A national reconnaissance of pharmaceuticals and other organic wastewater contaminants in the United States—I) Groundwater. Sci. Total Environ. 2008, 402, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Galan, M.; Diaz-Cruz, M.; Barcelo, D. Determination of 19 sulfonamides in environmental water samples by automated on-line solid-phase extraction-liquid chromatography–tandem mass spectrometry (SPE-LC-MS/MS). Talanta 2009, 81, 355–366. [Google Scholar] [CrossRef]
- Fram, M.S.; Belitz, K. Occurrence and concentrations of pharmaceutical compounds in groundwater used for public drinking-water supply in California. Sci. Total Environ. 2011, 409, 3409–3417. [Google Scholar] [CrossRef] [Green Version]
- Balzer, F.; Zühlke, S.; Hannappel, S. Antibiotics in groundwater under locations with high livestock density in Germany. Water Sci. Technol. Water Supply 2016, 16, 1361–1369. [Google Scholar] [CrossRef]
- Voigt, A.M.; Ciorba, P.; Döhla, M.; Exner, M.; Felder, C.; Lenz-Plet, F.; Faerber, H.A. The investigation of antibiotic residues, antibiotic resistance genes and antibiotic-resistant organisms in a drinking water reservoir system in Germany. Int. J. Hyg. Environ. Health 2020, 224, 113449. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Feng, M.; Wang, B.; Zhao, X.; Guo, R.; Bu, Y.; Chen, J. Distribution and potential risk assessment of antibiotic pollution in the main drinking water sources of Nanjing, China. Environ. Sci. Pollut. Res. 2020, 17, 21429–21441. [Google Scholar] [CrossRef]
- Wang, Q.J.; Mo, C.H.; Li, Y.W.; Gao, P.; Tai, Y.P.; Xu, J.W. Determination of four fluoroquinolone antibiotics in tap water in Guangzhou and Macao. Environ. Pollut. 2010, 158, 2350–2358. [Google Scholar] [CrossRef]
- Wang, H.; Wang, N.; Wang, B.; Zhao, Q.; Fang, H.; Fu, C.; Jiang, Q. Antibiotics in Drinking Water in Shanghai and Their Contribution to Antibiotic Exposure of School Children. Environ. Sci. Technol 2016, 50, 2692–2699. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Cheng, Y.; Zhang, Y.; Li, Z.; Yu, Y.; Feng, L.; Xu, L. Distribution and human health risk assessment of antibiotic residues in large-scale drinking water sources in Chongqing area of the Yangtze River. Environ. Res. 2020, 185, 109386. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Ho, K.W.K.; Ying, G.G.; Deng, W.J. Veterinary antibiotics in food, drinking water, and the urine of preschool children in Hong Kong. Environ. Int. 2017, 108, 246–252. [Google Scholar] [CrossRef]
- Díaz-Cruz, M.S.; García-Galán, M.J.; Barceló, D. Highly sensitive simultaneous determination of sulfonamide antibiotics and one metabolite in environmental waters by liquid chromatography–quadrupole linear ion trap–mass spectrometry. J. Chromatogr. A 2008, 1193, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Benotti, M.J.; Trenholm, R.A.; Vanderford, B.J.; Holady, J.C.; Stanford, B.D.; Snyder, S.A. Pharmaceuticals and Endocrine Disrupting Compounds in U.S. Drinking Water. Environ. Sci. Technol. 2009, 43, 597–603. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EC50 Value (Exposure) [mM] | EC50 Value (Recovery) [mM] | |
|---|---|---|
| Number of fronds | 1.944 | 2.515 |
| Fronds area | 1.724 | 2.850 |
| Fresh mass | 1.431 | 2.447 |
| Dry mass | 3.825 | 6.715 |
| Tetracycline Concentration [mM] during 7 Days | Chlorophyll Concentration [M] | |
|---|---|---|
| Exposure | Recovery | |
| 0 | 1.574 × 10−5 | 1.574 × 10−5 |
| 1 | 0.918 × 10−5 | 1.315 × 10−5 |
| 2.5 | 0.749 × 10−5 | 0.988 × 10−5 |
| 10 | 0.571 × 10−5 | 0.742 × 10−5 |
| Antioxidant Enzymes | EC50 Value (Exposure) [mM] | EC50 Value (Recovery) [mM] |
|---|---|---|
| Catalase activity | 2.711 | 6.506 |
| Ascorbate peroxidase | 2.173 | 5.231 |
| Pyrogallol peroxidase | 9.253 | 5.098 |
| Glutathione reductase | 1.612 | 3.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krupka, M.; Michalczyk, D.J.; Žaltauskaitė, J.; Sujetovienė, G.; Głowacka, K.; Grajek, H.; Wierzbicka, M.; Piotrowicz-Cieślak, A.I. Physiological and Biochemical Parameters of Common Duckweed Lemna minor after the Exposure to Tetracycline and the Recovery from This Stress. Molecules 2021, 26, 6765. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26226765
Krupka M, Michalczyk DJ, Žaltauskaitė J, Sujetovienė G, Głowacka K, Grajek H, Wierzbicka M, Piotrowicz-Cieślak AI. Physiological and Biochemical Parameters of Common Duckweed Lemna minor after the Exposure to Tetracycline and the Recovery from This Stress. Molecules. 2021; 26(22):6765. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26226765
Chicago/Turabian StyleKrupka, Magdalena, Dariusz J. Michalczyk, Jūratė Žaltauskaitė, Gintarė Sujetovienė, Katarzyna Głowacka, Hanna Grajek, Marta Wierzbicka, and Agnieszka I. Piotrowicz-Cieślak. 2021. "Physiological and Biochemical Parameters of Common Duckweed Lemna minor after the Exposure to Tetracycline and the Recovery from This Stress" Molecules 26, no. 22: 6765. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26226765