
Natural Herbal Estrogen-Mimetics (Phytoestrogens) Promote the Differentiation of Fallopian Tube Epithelium into Multi-Ciliated Cells via Estrogen Receptor Beta


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
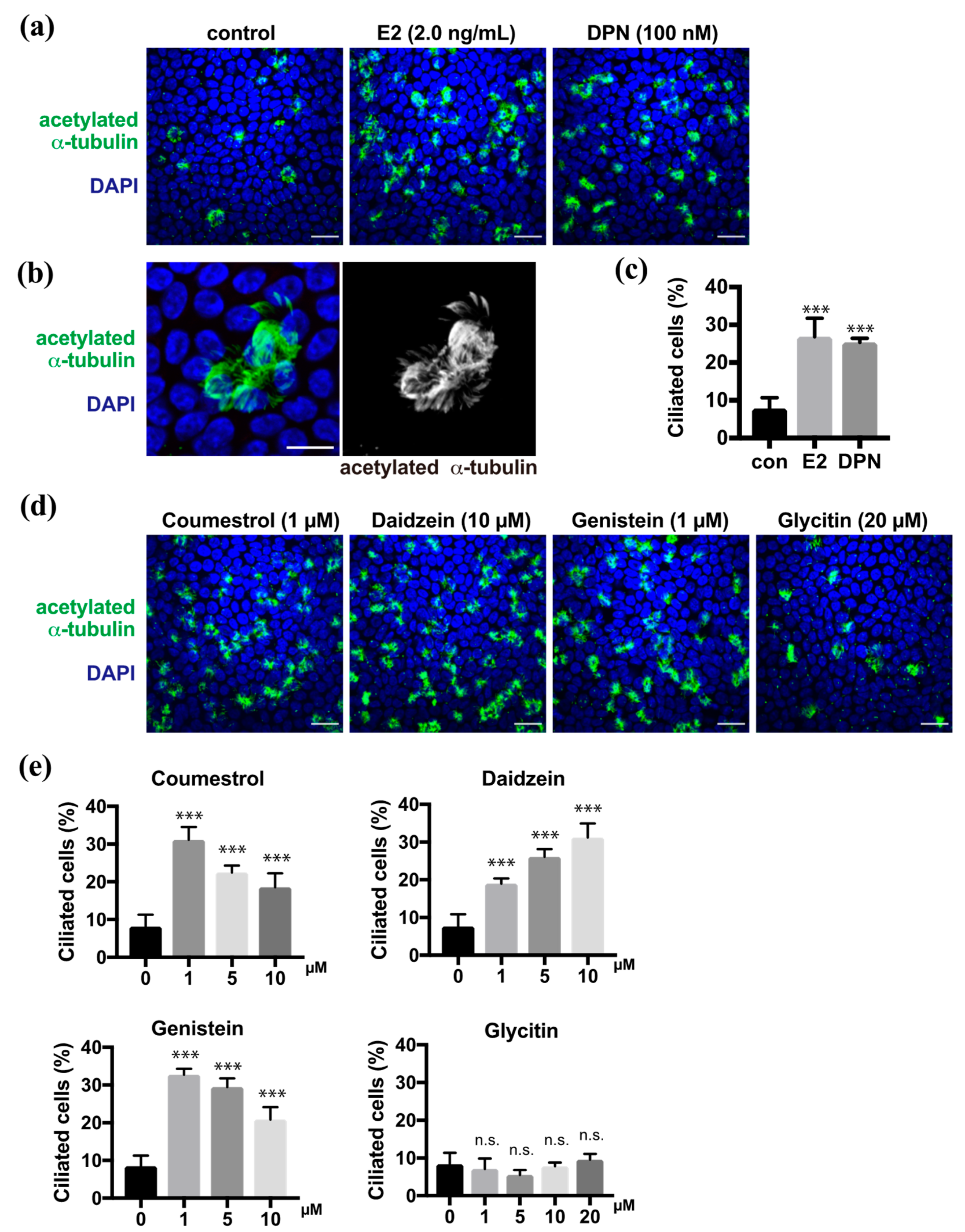
2.1. Phytoestrogens Promote Multi-Ciliated Cell Differentiation
2.2. Phytoestrogens Slightly Affect the Proportion of Secretory Cells and Basal Cells
2.3. Induction of Multi-Ciliated Cell Differentiation Depends on the ERβ Pathway
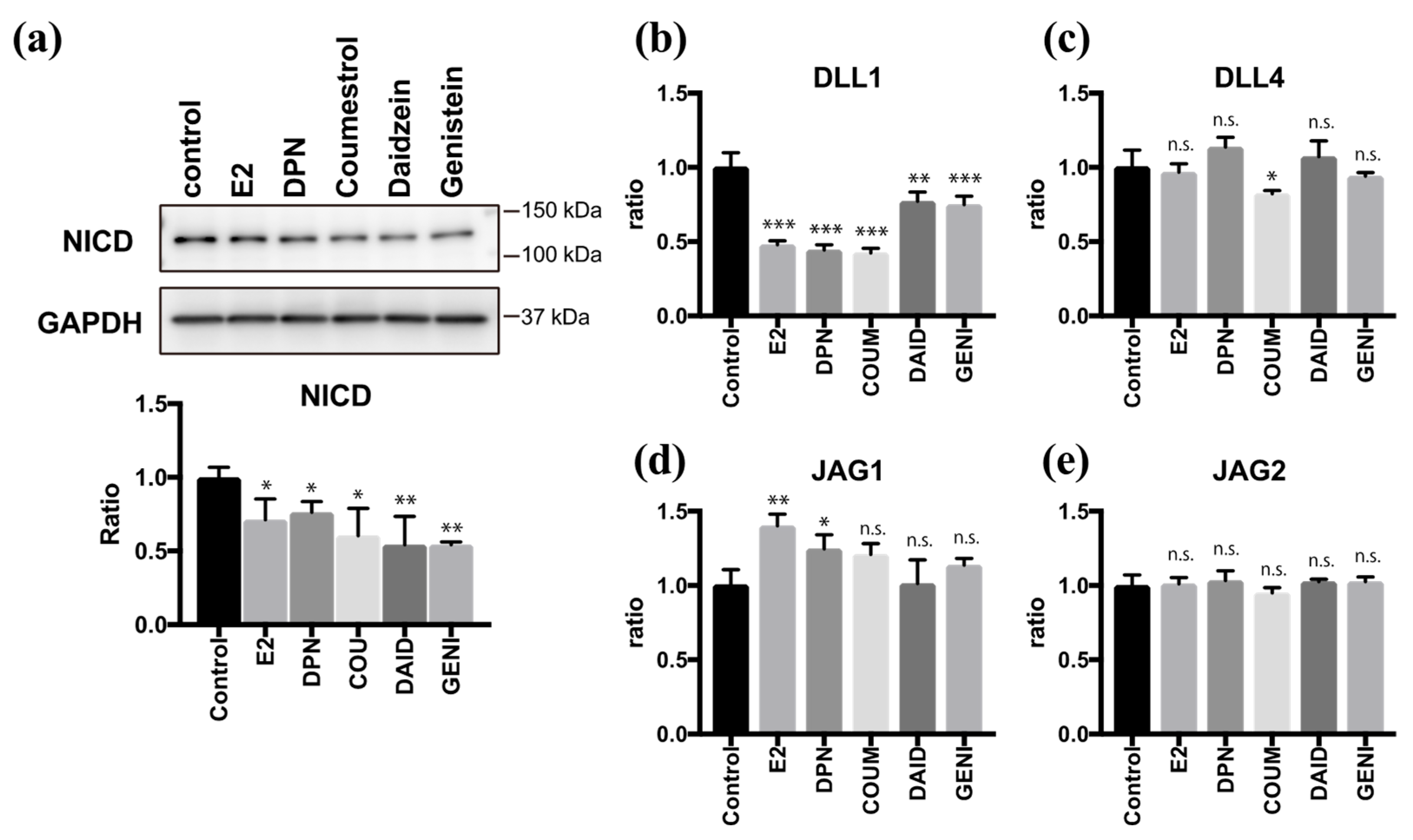
2.4. Coumestrol, Daidzein, and Genistein Suppress Notch Signaling
3. Discussion
4. Materials and Methods
4.1. Fallopian Tube Epithelial Cell Culture
4.2. Immunofluorescence
4.3. Immunoblot Analysis
4.4. Quantitative PCR
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lyons, R.A.; Saridogan, E.; Djahanbakhch, O. The reproductive significance of human Fallopian tube cilia. Hum. Reprod. Update 2006, 12, 363–372. [Google Scholar] [CrossRef]
- Pillai, V.V.; Weber, D.M.; Phinney, B.S.; Selvaraj, V. Profiling of proteins secreted in the bovine oviduct reveals diverse functions of this luminal microenvironment. PLoS ONE 2017, 12, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halbert, S.A.; Tam, P.Y.; Blandau, R.J. Egg transport in the rabbit oviduct: The roles of cilia and muscle. Science 1976, 191, 1052–1053. [Google Scholar] [CrossRef] [PubMed]
- Amso, N.N.; Crow, J.; Shaw, R.W. Endocrinology: Comparative immunohistochemical study of oestrogen and progesterone receptors in the fallopian tube and uterus at different stages of the menstrual cycle and the menopause. Hum. Reprod. 1994, 9, 1027–1037. [Google Scholar] [CrossRef]
- Verhage, H.G.; Mavrogianis, P.A.; Boice, M.L.; Li, W.; Fazleabas, A.T. Oviductal epithelium of the baboon: Hormonal control and the immuno-gold localization of oviduct-specific glycoproteins. Am. J. Anat. 1990, 187, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Donnez, J.; Casanas-Roux, F.; Ferin, J.; Thomas, K. Changes in ciliation and cell height in human tubal epithelium in the fertile and post-fertile years. Maturitas 1983, 5, 39–45. [Google Scholar] [CrossRef]
- Hewitt, S.C.; Korach, K.S. Estrogen receptors: New directions in the new millennium. Endocr. Rev. 2018, 39, 664–675. [Google Scholar] [CrossRef] [Green Version]
- Krege, J.H.; Hodgin, J.B.; Couse, J.F.; Enmark, E.; Warner, M.; Mahler, J.F.; Sar, M.; Korach, K.S.; Gustafsson, J.A.; Smithies, O. Generation and reproductive phenotypes of mice lacking estrogen receptor β. Proc. Natl. Acad. Sci. USA 1998, 95, 15677–15682. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Iwano, T.; Takeda, S. Estrogen and EGFR Pathways Regulate Notch Signaling in Opposing Directions for Multi-Ciliogenesis in the Fallopian Tube. Cells 2019, 8, 933. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-López, I.; Yago-Aragón, M.; Salas-Huetos, A.; Tresserra-Rimbau, A.; Hurtado-Barroso, S. Effects of dietary phytoestrogens on hormones throughout a human lifespan: A review. Nutrients 2020, 12, 1–25. [Google Scholar] [CrossRef]
- Alexander, V., S. Phytoestrogens and their effects. Eur. J. Pharmacol. 2014, 741, 230–236. [Google Scholar] [CrossRef]
- Poschner, S.; Maier-Salamon, A.; Zehl, M.; Wackerlig, J.; Dobusch, D.; Pachmann, B.; Sterlini, K.L.; Jäger, W. The Impacts of Genistein and Daidzein on Estrogen Conjugations in Human Breast Cancer Cells: A targeted metabolomics approach. Front. Pharmacol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, Y.; Kaihara, A.; Yoshii, K.; Tsumura, Y.; Ishimitsu, S.; Tonogai, Y. Content and composition of isoflavonoids in mature or immature beans and bean sprouts consumed in Japan. J. Heal. Sci. 2001, 47, 394–406. [Google Scholar] [CrossRef] [Green Version]
- Kostelac, D.; Rechkemmer, G.; Briviba, K. Phytoestrogens Modulate Binding Response of Estrogen Receptors α and β to the Estrogen Response Element. J. Agric. Food Chem. 2003, 51, 7632–7635. [Google Scholar] [CrossRef]
- Jiang, Y.; Gong, P.; Madak-Erdogan, Z.; Martin, T.; Jeyakumar, M.; Carlson, K.; Khan, I.; Smillie, T.J.; Chittiboyina, A.G.; Rotte, S.C.K.; et al. Mechanisms enforcing the estrogen receptor β selectivity of botanical estrogens. FASEB J. 2013, 27, 4406–4418. [Google Scholar] [CrossRef] [Green Version]
- Morito, K.; Hirose, T.; Kinjo, J.; Hirakawa, T.; Okawa, M.; Nohara, T.; Ogawa, S.; Inoue, S.; Muramatsu, M.; Masamune, Y. Interaction of phytoestrogens with estrogen receptors α andβ. Biol. Pharm. Bull. 2001, 24, 351–356. [Google Scholar] [CrossRef]
- Chan, K.K.L.; Siu, M.K.Y.; Jiang, Y. xin; Wang, J. jing; Leung, T.H.Y.; Ngan, H.Y.S. Estrogen receptor modulators genistein, daidzein and ERB-041 inhibit cell migration, invasion, proliferation and sphere formation via modulation of FAK and PI3K/AKT signaling in ovarian cancer. Cancer Cell Int. 2018, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Russo, M.; Russo, G.L.; Daglia, M.; Kasi, P.D.; Ravi, S.; Nabavi, S.F.; Nabavi, S.M. Understanding genistein in cancer: The “good” and the “bad” effects: A review. Food Chem. 2016, 196, 589–600. [Google Scholar] [CrossRef]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, V.; Bishayee, A. Molecular mechanisms of action of genistein in cancer: Recent advances. Front. Pharmacol. 2019, 10, 1336. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Iwano, T.; Takeda, S. Fallopian tube basal stem cells reproducing the epithelial sheets in vitro—stem cell of fallopian epithelium. Biomolecules 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Kessler, M.; Hoffmann, K.; Brinkmann, V.; Thieck, O.; Jackisch, S.; Toelle, B.; Berger, H.; Mollenkopf, H.-J.; Mangler, M.; Sehouli, J.; et al. The Notch and Wnt pathways regulate stemness and differentiation in human fallopian tube organoids. Nat. Commun. 2015, 6, 8989. [Google Scholar] [CrossRef] [PubMed]
- Morton, M.S.; Arisaka, O.; Miyake, N.; Morgan, L.D.; Evans, B.A.J. Phytoestrogen concentrations in serum from Japanese men and women over forty years of age. J. Nutr. 2002, 132, 3168–3171. [Google Scholar] [CrossRef] [PubMed]
- Valentín-Blasini, L.; Blount, B.C.; Caudill, S.P.; Needham, L.L. Urinary and serum concentrations of seven phytoestrogens in a human reference population subset. J. Expo. Anal. Environ. Epidemiol. 2003, 13, 276–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.M.; Huh, J.S.; Lim, Y.; Cho, M. Soy isoflavone glycitin (4′-hydroxy-6-methoxyisoflavone-7-D-Glucoside) promotes human dermal fibroblast cell proliferation and migration via TGF-β signaling. Phyther. Res. 2015, 29, 757–769. [Google Scholar] [CrossRef]
- Simintiras, C.A.; Sturmey, R.G. Genistein crosses the bioartificial oviduct and alters secretion composition. Reprod. Toxicol. 2017, 71, 63–70. [Google Scholar] [CrossRef]
- Jefferson, W.N.; Padilla-Banks, E.; Goulding, E.H.; Lao, S.P.C.; Newbold, R.R.; Williams, C.J. Neonatal exposure to genistein disrupts ability of female mouse reproductive tract to support preimplantation embryo development and implantation. Biol. Reprod. 2009, 80, 425–431. [Google Scholar] [CrossRef]




Sample Availability: Samples of the compounds and cells are available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, M.; Takeda, S.; Iwano, T. Natural Herbal Estrogen-Mimetics (Phytoestrogens) Promote the Differentiation of Fallopian Tube Epithelium into Multi-Ciliated Cells via Estrogen Receptor Beta. Molecules 2021, 26, 722. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26030722
Zhu M, Takeda S, Iwano T. Natural Herbal Estrogen-Mimetics (Phytoestrogens) Promote the Differentiation of Fallopian Tube Epithelium into Multi-Ciliated Cells via Estrogen Receptor Beta. Molecules. 2021; 26(3):722. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26030722
Chicago/Turabian StyleZhu, Maobi, Sen Takeda, and Tomohiko Iwano. 2021. "Natural Herbal Estrogen-Mimetics (Phytoestrogens) Promote the Differentiation of Fallopian Tube Epithelium into Multi-Ciliated Cells via Estrogen Receptor Beta" Molecules 26, no. 3: 722. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26030722