
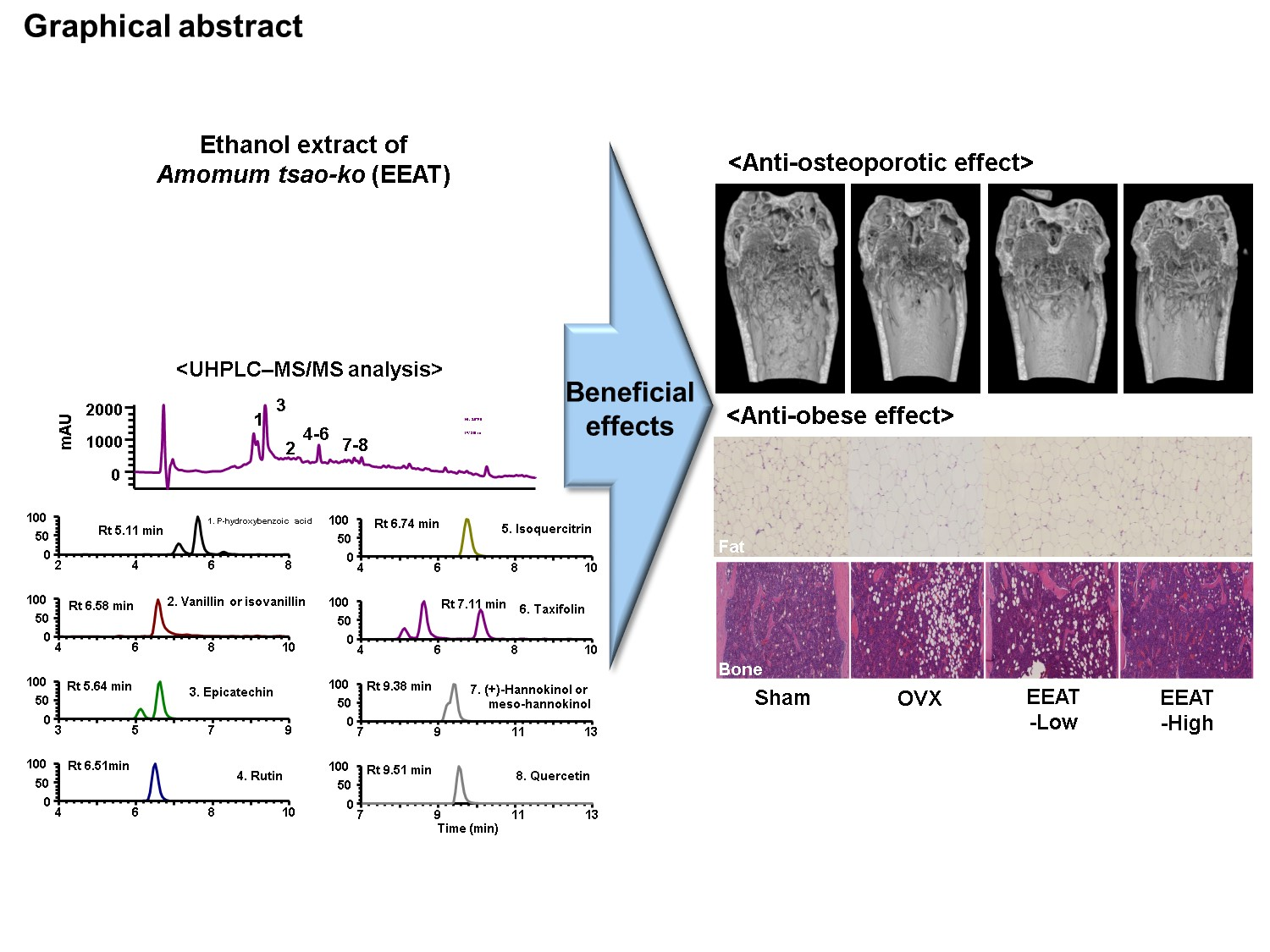
Ethanol Extract of Amomum tsao-ko Ameliorates Ovariectomy-Induced Trabecular Loss and Fat Accumulation


Abstract
:
1. Introduction
2. Results and Discussion
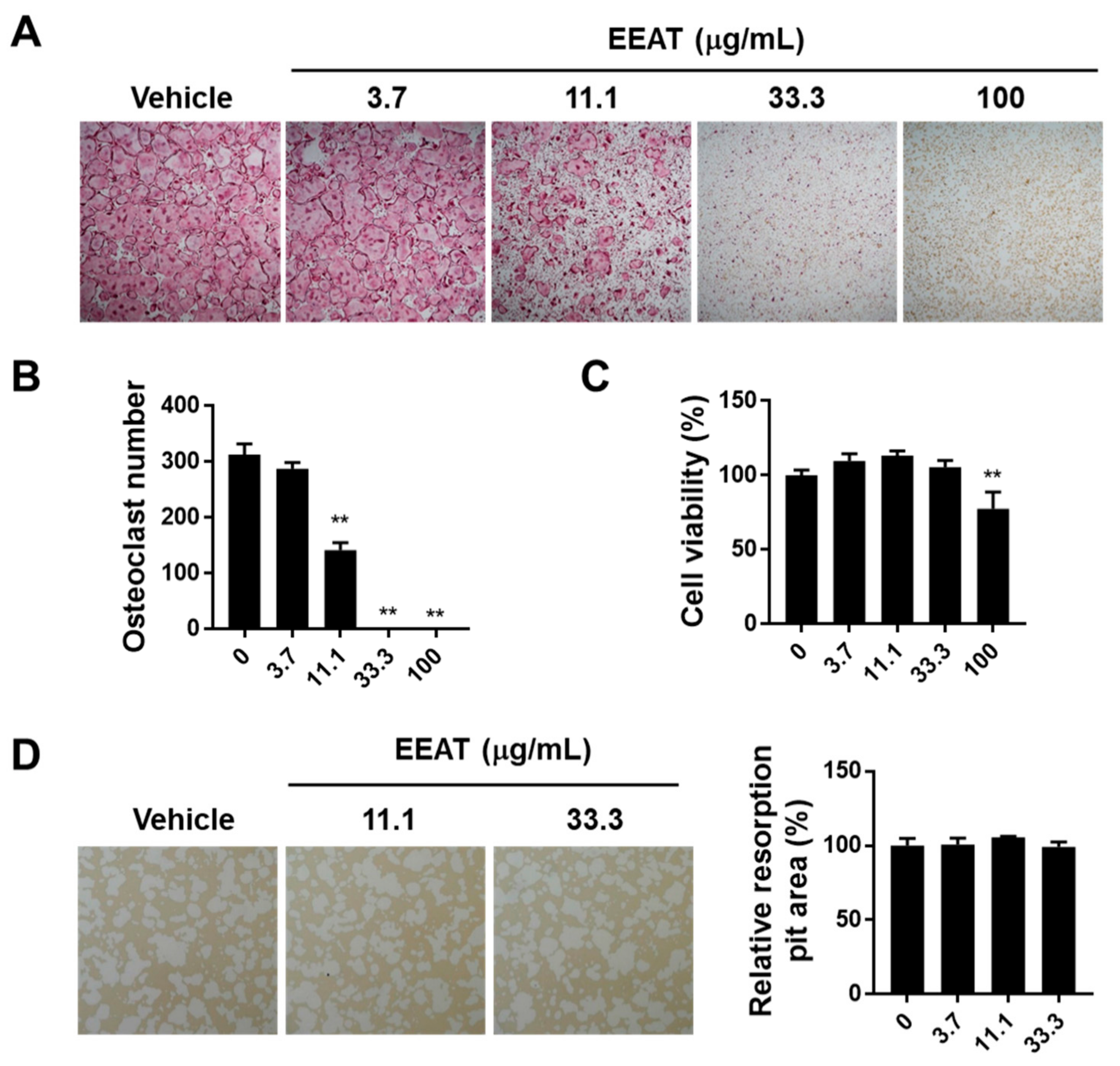
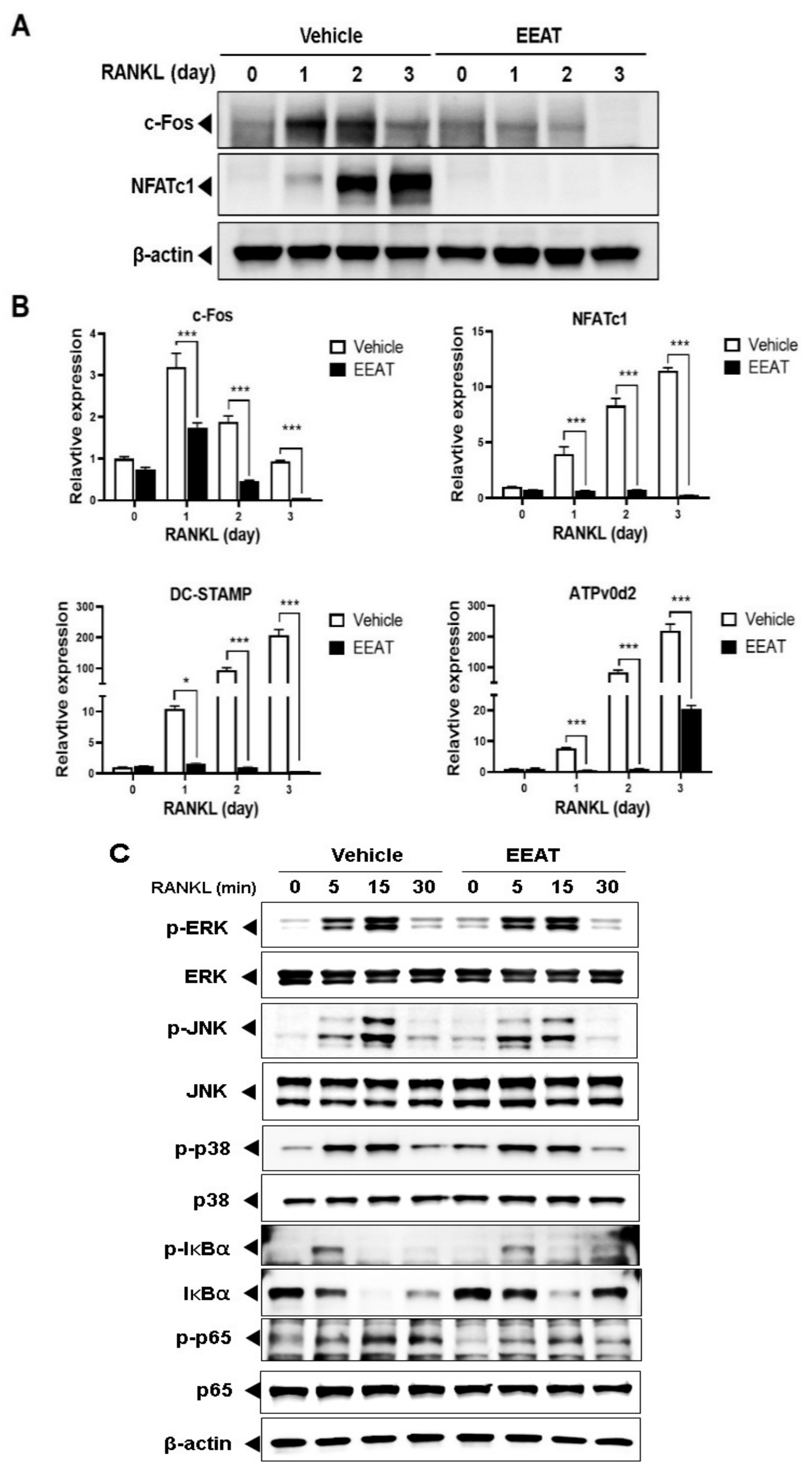
2.1. EEAT Inhibits RANKL Signaling during Osteoclast Differentiation
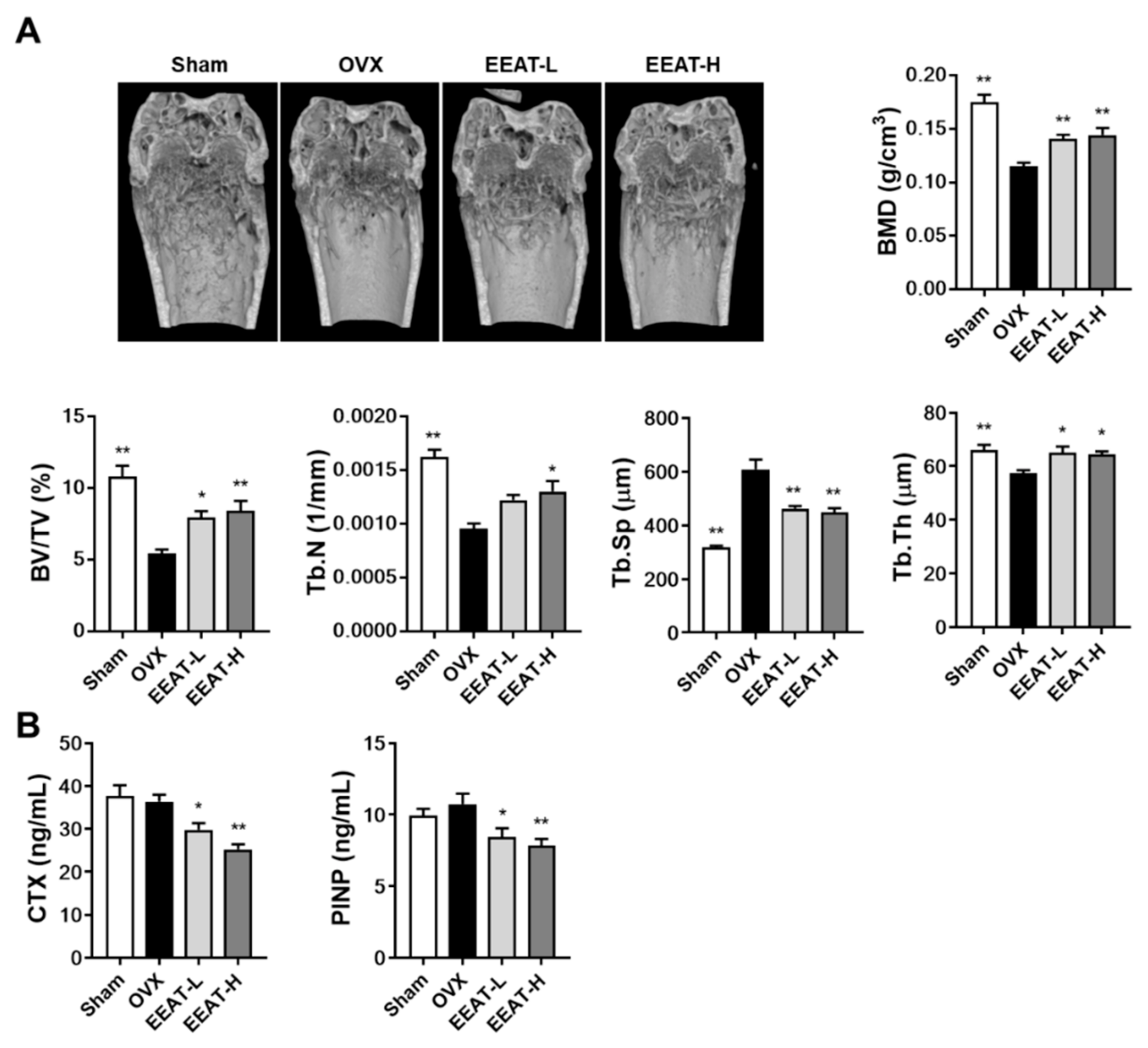
2.2. EEAT Ameliorates Bone Loss in OVX Mice
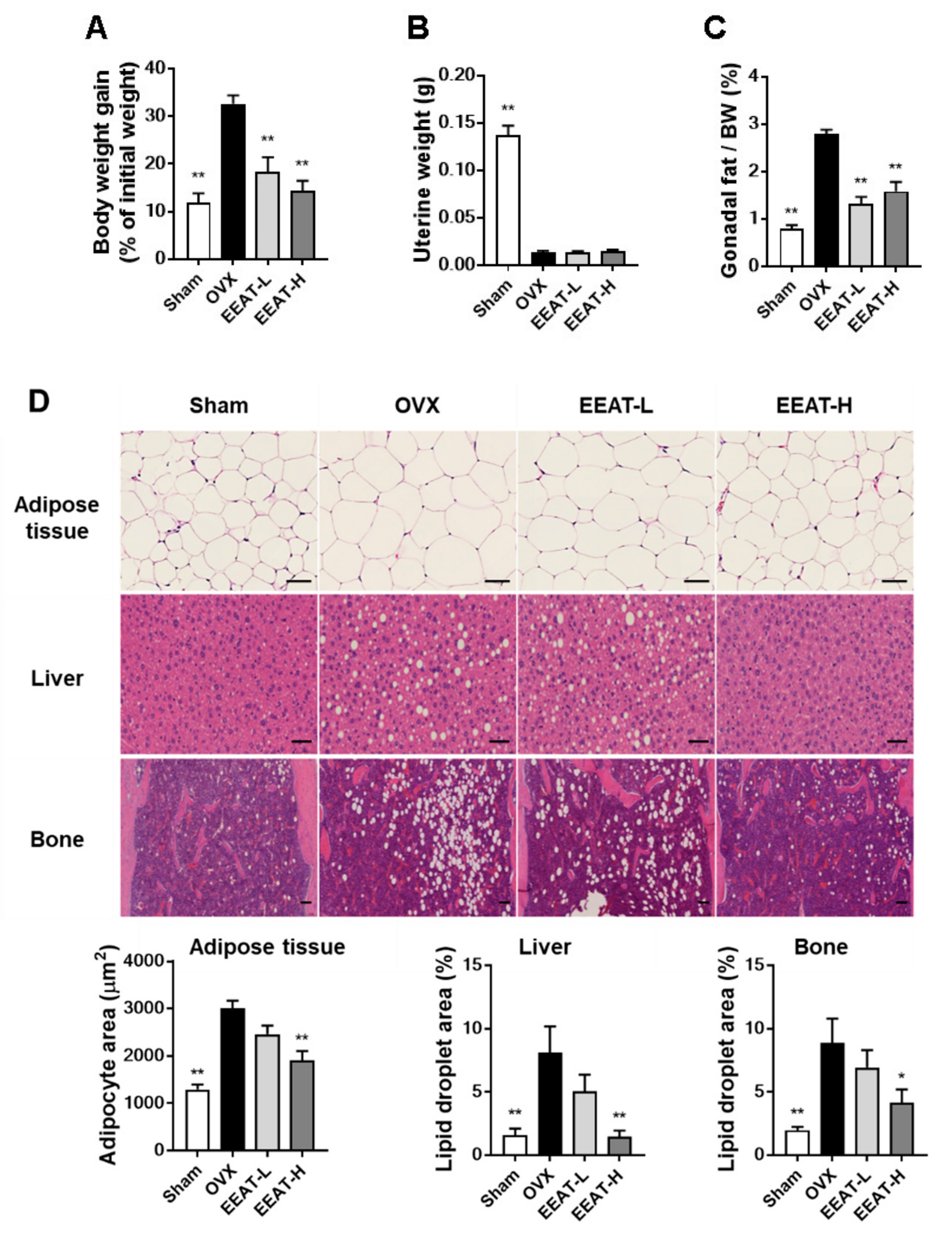
2.3. EEAT Inhibits Fat Accumulation in OVX Mice
2.4. Phytochemical Profiling of EEAT
3. Materials and Methods
3.1. Materials
3.2. Osteoclast Culture and Bone Resorption Assay
3.3. Quantitative Real-Time Polymerase Chain Reaction (RT-PCR)
3.4. Western Blot
3.5. Animal Study
3.6. Micro-CT Analysis
3.7. Measurement of Bone Turnover Markers
3.8. Histological Analysis
3.9. UHPLC–MS/MS
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Marjoribanks, J.; Farquhar, C.; Roberts, H.; Lethaby, A.; Lee, J. Long-term hormone therapy for perimenopausal and postmenopausal women. Cochrane Database Syst. Rev. 2017, 1, CD004143. [Google Scholar] [CrossRef] [PubMed]
- Gurney, E.P.; Nachtigall, M.J.; Nachtigall, L.E.; Naftolin, F. The Women’s Health Initiative trial and related studies: 10 years later: A clinician’s view. J. Steroid Biochem. Mol. Biol. 2014, 142, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, E.F.; Langdahl, B.; Vesterby, A.; Rungby, J.; Kassem, M. Hormone Replacement Therapy Prevents Osteoclastic Hyperactivity: A Histomorphometric Study in Early Postmenopausal Women. J. Bone Miner. Res. 1999, 14, 1217–1221. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, C.S.; Roberts, R.M.; Lubahn, D.B. Estrogen receptor- and aromatase-deficient mice provide insight into the roles of estrogen within the ovary and uterus. Mol. Reprod. Dev. 2001, 59, 336–346. [Google Scholar] [CrossRef]
- Modder, U.I.L.; Riggs, B.L.; Spelsberg, T.C.; Fraser, D.G.; Atkinson, E.J.; Arnold, R.; Khosla, S. Dose-response of estrogen on bone versus the uterus in ovariectomized mice. Eur. J. Endocrinol. 2004, 151, 503–510. [Google Scholar] [CrossRef]
- Manolagas, S.C.; O’Brien, C.A.; Almeida, M. The role of estrogen and androgen receptors in bone health and disease. Nat. Rev. Endocrinol. 2013, 9, 699–712. [Google Scholar] [CrossRef]
- Brown, L.M.; Clegg, D.J. Central effects of estradiol in the regulation of food intake, body weight, and adiposity. J. Steroid Biochem. Mol. Biol. 2010, 122, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Kanaley, J.; Sames, C.; Swisher, L.; Swick, A.; Ploutz-Snyder, L.; Steppan, C.; Sagendorf, K.; Feiglin, D.; Jaynes, E.; Meyer, R.; et al. Abdominal fat distribution in pre- and postmenopausal women: The impact of physical activity, age, and menopausal status. Metab. Clin. Exp. 2001, 50, 976–982. [Google Scholar] [CrossRef]
- Davis, S.R.; Castelo-Branco, C.; Chedraui, P.; Lumsden, M.A.; Nappi, R.E.; Shah, D.; Villaseca, P. As the Writing Group of the International Menopause Society for World Menopause Day 2012 Understanding weight gain at menopause. Climacteric 2012, 15, 419–429. [Google Scholar] [CrossRef]
- De Franciscis, P.; Colacurci, N.; Riemma, G.; Conte, A.; Pittana, E.; Guida, M.; Schiattarella, A. A Nutraceutical Approach to Menopausal Complaints. Medicina 2019, 55, 544. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.-S.; Lee, J.-Y.; Cho, S.-C. Isotsaokoin, an Antifungal Agent from Amomumtsao-ko. J. Nat. Prod. 2004, 67, 889–891. [Google Scholar] [CrossRef] [PubMed]
- Alam, K.; Pathak, D.; Ansari, S.H. Evaluation of anti-inflammatory activity of Ammomum subulatum fruit extract. Int. J. Pharm. Sci. Drug Res. 2011, 3, 35–37. [Google Scholar]
- Li, B.; Choi, H.-J.; Lee, D.-S.; Oh, H.; Kim, Y.-C.; Moon, J.-Y.; Park, W.-H.; Park, S.-D.; Kim, J.-E. Amomum tsao-ko Suppresses Lipopolysaccharide-Induced Inflammatory Responses in RAW264.7 Macrophages via Nrf2-Dependent Heme Oxygenase-1 Expression. Am. J. Chin. Med. 2014, 42, 1229–1244. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Wang, L.-T.; Liu, J.-Z.; Wang, H.-M.; Guo, N.; Gu, C.-B.; Fu, Y. Rapid extraction of Amomum tsao-ko essential oil and determination of its chemical composition, antioxidant and antimicrobial activities. J. Chromatogr. B 2017, 1061, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.S.; Kikuzaki, H.; Hisamoto, M.; Nakatani, N. Constituents of Amomum tsao-ko and their radical scavenging and antioxidant activities. J. Am. Oil Chem. Soc. 2000, 77, 667–673. [Google Scholar] [CrossRef]
- Bairwa, G.L.; Jasuja, N.D.; Joshi, S.C. Lipid lowering and antioxidant effects of Amomum subulatum seeds (Family Zingiberaceae) in cholesterol fed rabbits. Arch. Phytopathol. Plant Prot. 2011, 44, 1425–1431. [Google Scholar] [CrossRef]
- Bhaswant, M.; Poudyal, H.; Mathai, M.L.; Ward, L.C.; Mouatt, P.; Brown, L. Green and Black Cardamom in a Diet-Induced Rat Model of Metabolic Syndrome. Nutrients 2015, 7, 7691–7707. [Google Scholar] [CrossRef]
- Lee, J.A.; Park, Y.J.; Jeong, W.; Hong, S.S.; Ahn, E.-K. Anti-obesity effect ofAmomum taso-koethanol extract in 3T3-L1 adipocytes. J. Appl. Biol. Chem. 2017, 60, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Corbo, F.; Brunetti, G.; Crupi, P.; Bortolotti, S.; Storlino, G.; Piacente, L.; Carocci, A.; Catalano, A.; Milani, G.; Colaianni, G.; et al. Effects of Sweet Cherry Polyphenols on Enhanced Osteoclastogenesis Associated With Childhood Obesity. Front. Immunol. 2019, 10, 1001. [Google Scholar] [CrossRef]
- Huang, H.; Chang, E.-J.; Ryu, J.; Lee, Z.H.; Lee, Y.; Kim, H.-H. Induction of c-Fos and NFATc1 during RANKL-stimulated osteoclast differentiation is mediated by the p38 signaling pathway. Biochem. Biophys. Res. Commun. 2006, 351, 99–105. [Google Scholar] [CrossRef]
- Kim, K.; Lee, S.-H.; Kim, J.H.; Choi, Y.; Kim, N. NFATc1 Induces Osteoclast Fusion Via Up-Regulation of Atp6v0d2 and the Dendritic Cell-Specific Transmembrane Protein (DC-STAMP). Mol. Endocrinol. 2008, 22, 176–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Dou, C.E.; Xu, J.; Dong, S. DC-STAMP, the Key Fusion-Mediating Molecule in Osteoclastogenesis. J. Cell. Physiol. 2014, 229, 1330–1335. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Yao, Z.; Li, F.; Zhang, Q.; Badell, I.R.; Schwarz, E.M.; Takeshita, S.; Wagner, E.F.; Noda, M.; Matsuo, K.; et al. NF-κB p50 and p52 Regulate Receptor Activator of NF-κB Ligand (RANKL) and Tumor Necrosis Factor-induced Osteoclast Precursor Differentiation by Activating c-Fos and NFATc1. J. Biol. Chem. 2007, 282, 18245–18253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lufkin, E.G.; Wahner, H.W.; O’Fallon, W.M.; Hodgson, S.F.; Kotowicz, M.A.; Lane, A.W.; Judd, H.L.; Caplan, R.H.; Riggs, B.L. Treatment of Postmenopausal Osteoporosis with Transdermal Estrogen. Ann. Intern. Med. 1992, 117, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Väänänen, H.K.; Härkönen, P.L. Estrogen and bone metabolism. Maturitas 1996, 23, S65–S69. [Google Scholar] [CrossRef]
- Halade, G.V.; Rahman, M.; Williams, P.J.; Fernandes, G. High fat diet-induced animal model of age-associated obesity and osteoporosis. J. Nutr. Biochem. 2010, 21, 1162–1169. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Stubbins, R.E.; Smith, R.R.; Harvey, A.E.; Núñez, N.P. Differential susceptibility to obesity between male, female and ovariectomized female mice. Nutr. J. 2009, 8, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhuley, J.N. Anti-oxidant effects of cinnamon (Cinnamomum verum) bark and greater cardamom (Amomum subulatum) seeds in rats fed high fat diet. Indian J. Exp. Boil. 1999, 37, 238–242. [Google Scholar]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar] [CrossRef]
- Fenkci, V.; Rota, S.; Sabir, N.; Sermez, Y.; Guclu, A.; Akdag, B. Relationship of Serum Interleukin-6 and Tumor Necrosis Factor α Levels with Abdominal Fat Distribution Evaluated by Ultrasonography in Overweight or Obese Postmenopausal Women. J. Investig. Med. 2006, 54, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Botolin, S.; McCabe, L.R. Inhibition of PPARγ prevents type I diabetic bone marrow adiposity but not bone loss. J. Cell. Physiol. 2006, 209, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.-H.; Li, F.-G.; Chen, X.-Y.; Li, J.-T.; Wu, Y.-H.; Huang, L.-H.; Wang, Z.; Li, P.; Wang, T.; Lahn, B.T.; et al. PPARγ suppression inhibits adipogenesis but does not promote osteogenesis of human mesenchymal stem cells. Int. J. Biochem. Cell Biol. 2012, 44, 377–384. [Google Scholar] [CrossRef]
- Lee, H.-H.; Jang, J.-W.; Lee, J.-K.; Park, C.K. Rutin Improves Bone Histomorphometric Values by Reduction of Osteoclastic Activity in Osteoporosis Mouse Model Induced by Bilateral Ovariectomy. J. Korean Neurosurg. Soc. 2020, 63, 433–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayed, H.A.; Barakat, B.M.; Elshaer, S.S.; Abdel-Naim, A.B.; Menze, E.T. Antiosteoporotic activities of isoquercitrin in ovariectomized rats: Role of inhibiting hypoxia inducible factor-1 alpha. Eur. J. Pharmacol. 2019, 865, 172785. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Liu, C.; Zhao, L.; Liu, H.; Li, W.; Guan, H.; Zhao, L.; Xiao, J. Effects of Taxifolin on Osteoclastogenesis in vitro and in vivo. Front. Pharmacol. 2018, 9, 1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyung, T.-W.; Lee, J.-E.; Shin, H.-H.; Choi, H.-S. Rutin inhibits osteoclast formation by decreasing reactive oxygen species and TNF-α by inhibiting activation of NF-κB. Exp. Mol. Med. 2008, 40, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-Q.; Wang, Y.-J.; Yang, G.-T.; Gao, Q.-L.; Tang, M.-X. Taxifolin Inhibits Receptor Activator of NF-κB Ligand-Induced Osteoclastogenesis of Human Bone Marrow-Derived Macrophages in vitro and Prevents Lipopolysaccharide-Induced Bone Loss in vivo. Pharmacology 2018, 103, 101–109. [Google Scholar] [CrossRef]
- Jiang, H.; Yoshioka, Y.; Yuan, S.; Horiuchi, Y.; Yamashita, Y.; Croft, K.D.; Ashida, H. Enzymatically modified isoquercitrin promotes energy metabolism through activating AMPKα in male C57BL/6 mice. Food Funct. 2019, 10, 5188–5202. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Xu, N.; Zhao, W.; Su, J.; Liang, M.; Xie, Z.; Wu, X.; Li, Q. (-)-Epicatechin regulates blood lipids and attenuates hepatic steatosis in rats fed high-fat diet. Mol. Nutr. Food Res. 2017, 61, 1700303. [Google Scholar] [CrossRef]
- Perdicaro, D.J.; Lanzi, C.R.; Tudela, J.G.; Miatello, R.M.; Oteiza, P.I.; Prieto, M.A.V. Quercetin attenuates adipose hypertrophy, in part through activation of adipogenesis in rats fed a high-fat diet. J. Nutr. Biochem. 2020, 79, 108352. [Google Scholar] [CrossRef] [PubMed]
- Arts, I.C.W.; Sesink, A.L.A.; Faassen-Peters, M.; Hollman, P.C.H. The type of sugar moiety is a major determinant of the small intestinal uptake and subsequent biliary excretion of dietary quercetin glycosides. Br. J. Nutr. 2004, 91, 841–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, H.; Shim, K.-S.; Kim, T.; An, H.; Lee, C.-J.; Lee, K.J.; Ma, J.Y. Water Extract of Acer tegmentosum Reduces Bone Destruction by Inhibiting Osteoclast Differentiation and Function. Molecules 2014, 19, 3940–3954. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, G.; Storlino, G.; Oranger, A.; Colaianni, G.; Faienza, M.F.; Ingravallo, G.; Di Comite, M.; Reseland, J.E.; Celi, M.; Tarantino, U.; et al. LIGHT/TNFSF14 regulates estrogen deficiency-induced bone loss. J. Pathol. 2020, 250, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Shim, K.-S.; Hwang, Y.-H.; Jang, S.-A.; Kim, T.; Ha, H. Water Extract of Lysimachia christinae Inhibits Trabecular Bone Loss and Fat Accumulation in Ovariectomized Mice. Nutrients 2020, 12, 1927. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Rt (Min) | Calculated (m/z) | Estimated (m/z) | Adducts | Error (ppm) | Formula | MS/MS Fragments (m/z) | Identifications |
|---|---|---|---|---|---|---|---|---|
| 1 | 5.06 | 139.0390 | 139.0388 | [M + H]+ | −1.5296 | C7H6O3 | - | p-Hydroxybenzoic acid * |
| 2 | 6.52 | 153.0546 | 153.0543 | [M + H]+ | −1.852 | C8H8O3 | 153.0542, 125.0595, 93.0338 | Vanillin or isovanillin * |
| 3 | 5.64 | 289.0718 | 289.0723 | [M − H]− | 1.8547 | C15H14O6 | 289.0721, 245.0818, 203.0710, 179.0342 | Epicatechin * |
| 4 | 6.53 | 609.1461 | 609.1472 | [M − H]− | 1.817 | C27H30O16 | 609.1463, 301.0349, 300.0278 | Rutin * |
| 5 | 6.79 | 463.0882 | 463.0890 | [M − H]− | 1.7702 | C21H20O12 | 463.0888, 301.0348, 300.0278 | Isoquercitrin * |
| 6 | 7.12 | 303.0510 | 303.0515 | [M − H]− | 1.596 | C15H12O7 | 285.0408, 177.0185, 125.0231 | Taxifolin * |
| 7 | 9.48 | 315.1602 | 315.1606 | [M − H]− | 1.554 | C19H24O4 | 315.1605, 149.0597 | Meso-hannokinol or (+)-hannokinol |
| 8 | 9.58 | 301.0354 | 301.0358 | [M − H]− | 1.3856 | C15H10O7 | 301.0356, 178.9977, 151.0026 | Quercetin * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shim, K.-S.; Hwang, Y.-H.; Jang, S.-A.; Kim, T.; Ha, H. Ethanol Extract of Amomum tsao-ko Ameliorates Ovariectomy-Induced Trabecular Loss and Fat Accumulation. Molecules 2021, 26, 784. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26040784
Shim K-S, Hwang Y-H, Jang S-A, Kim T, Ha H. Ethanol Extract of Amomum tsao-ko Ameliorates Ovariectomy-Induced Trabecular Loss and Fat Accumulation. Molecules. 2021; 26(4):784. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26040784
Chicago/Turabian StyleShim, Ki-Shuk, Youn-Hwan Hwang, Seon-A Jang, Taesoo Kim, and Hyunil Ha. 2021. "Ethanol Extract of Amomum tsao-ko Ameliorates Ovariectomy-Induced Trabecular Loss and Fat Accumulation" Molecules 26, no. 4: 784. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26040784