
Effects of Abiotic Elicitors on Expression and Accumulation of Three Candidate Benzophenanthridine Alkaloids in Cultured Greater Celandine Cells

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
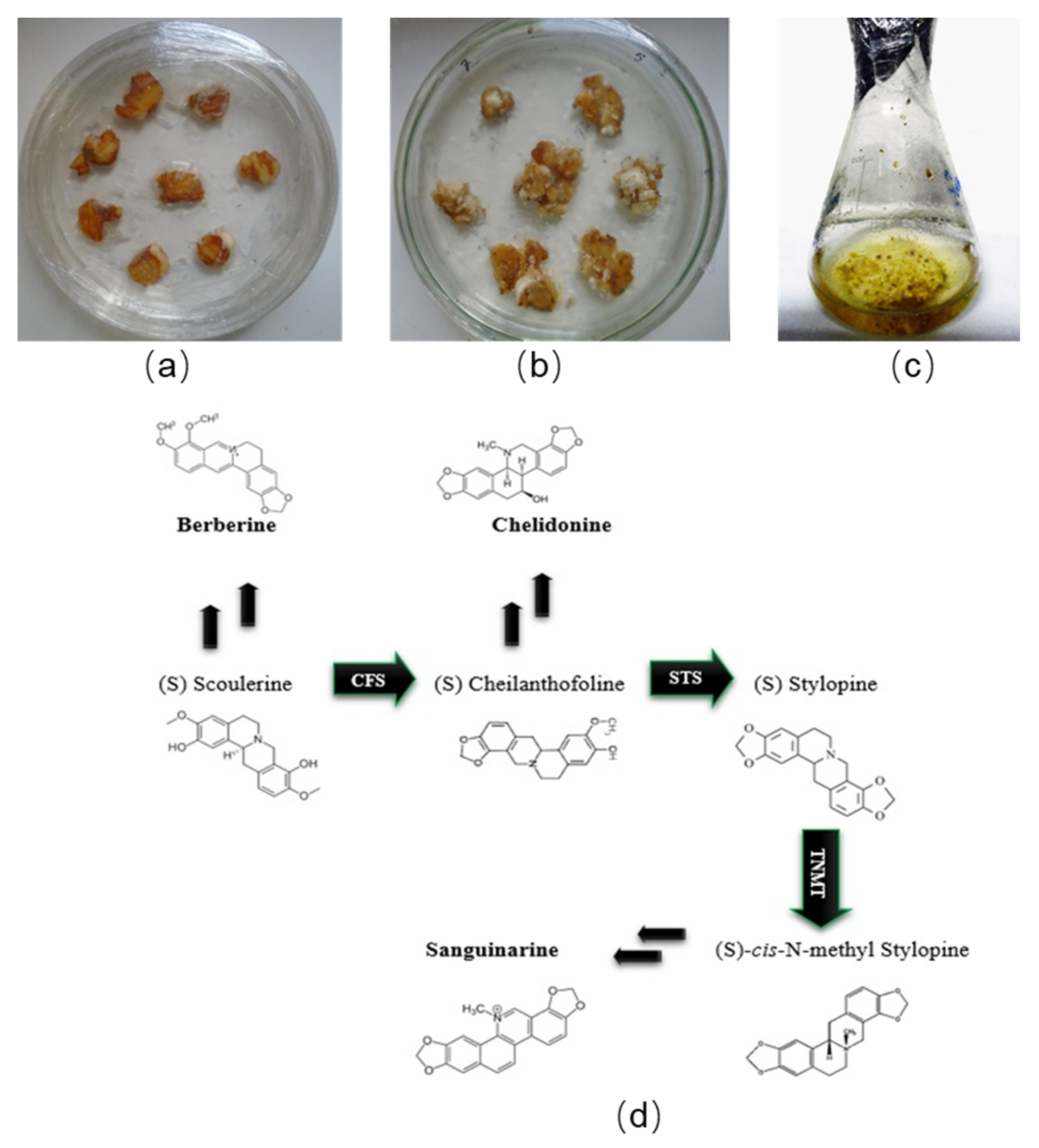
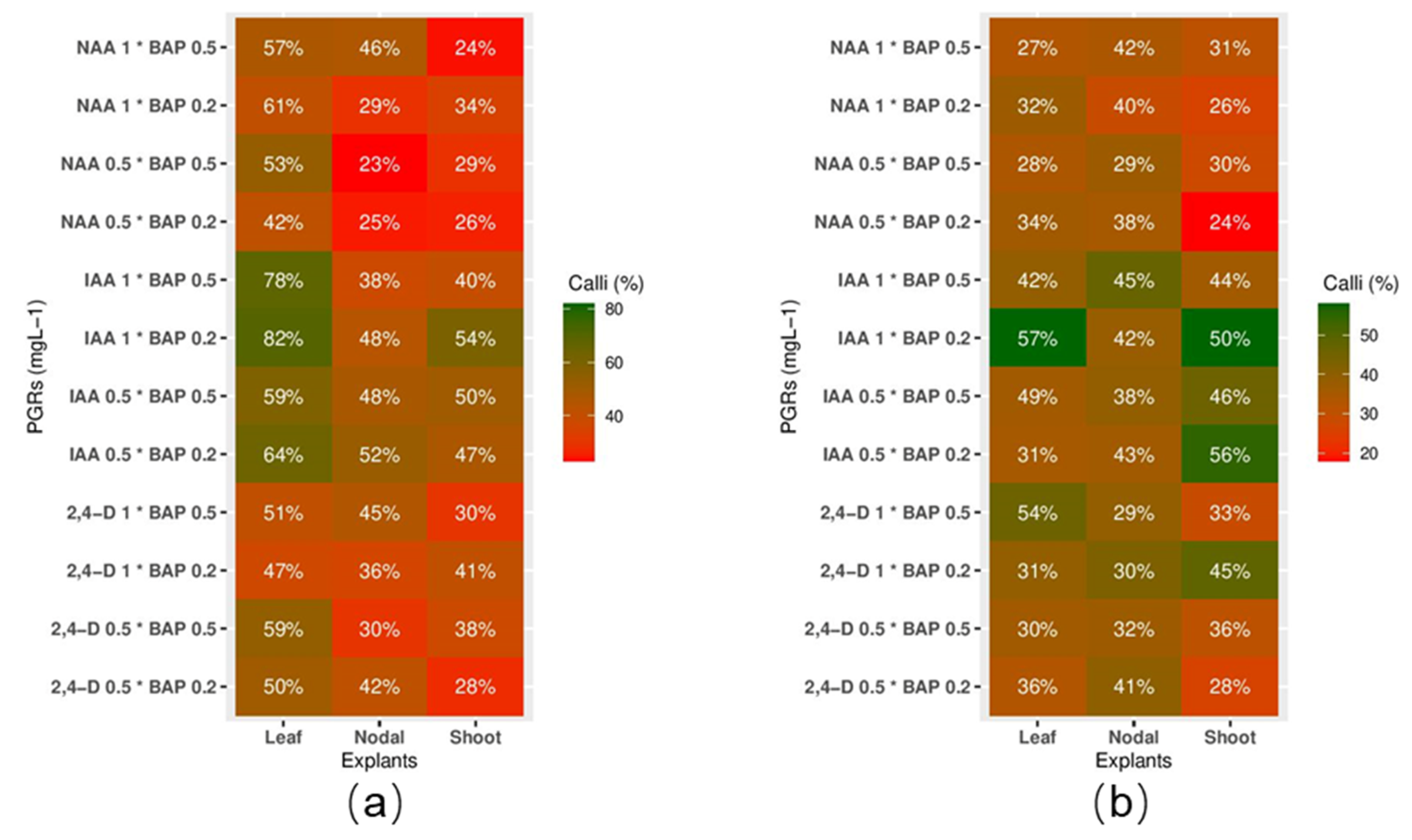
2.1. Propagation Optimization
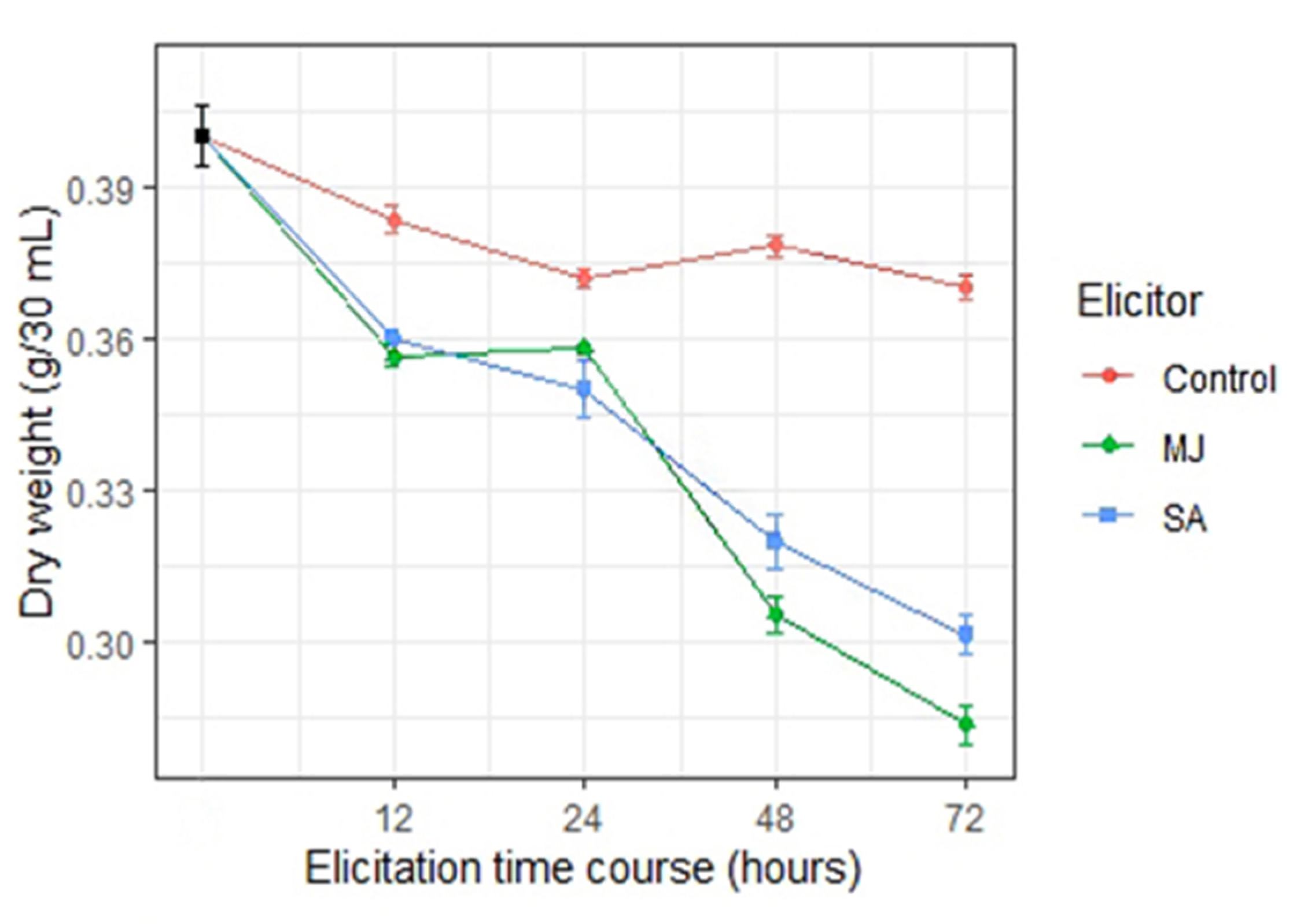
2.2. Cell Growth
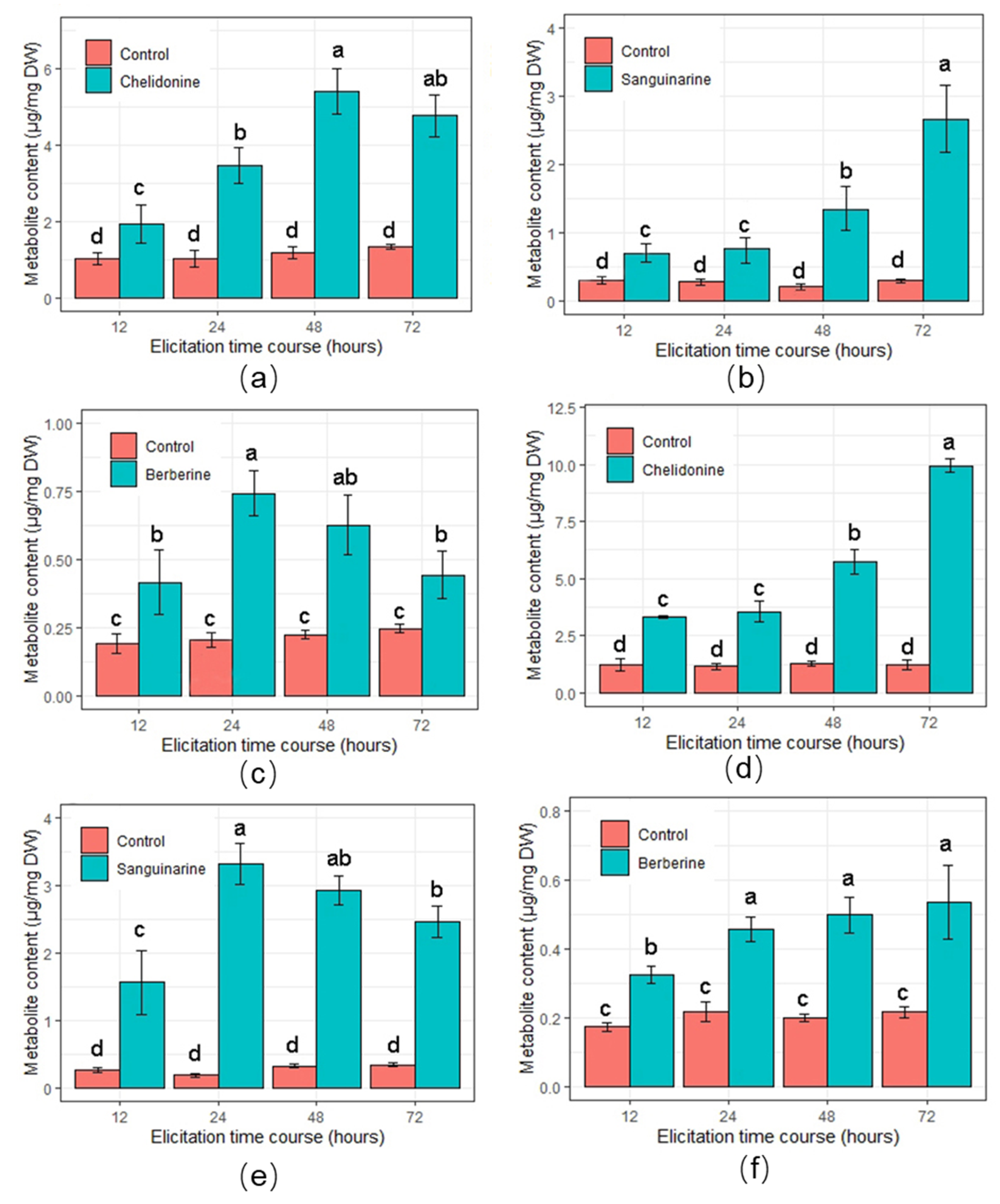
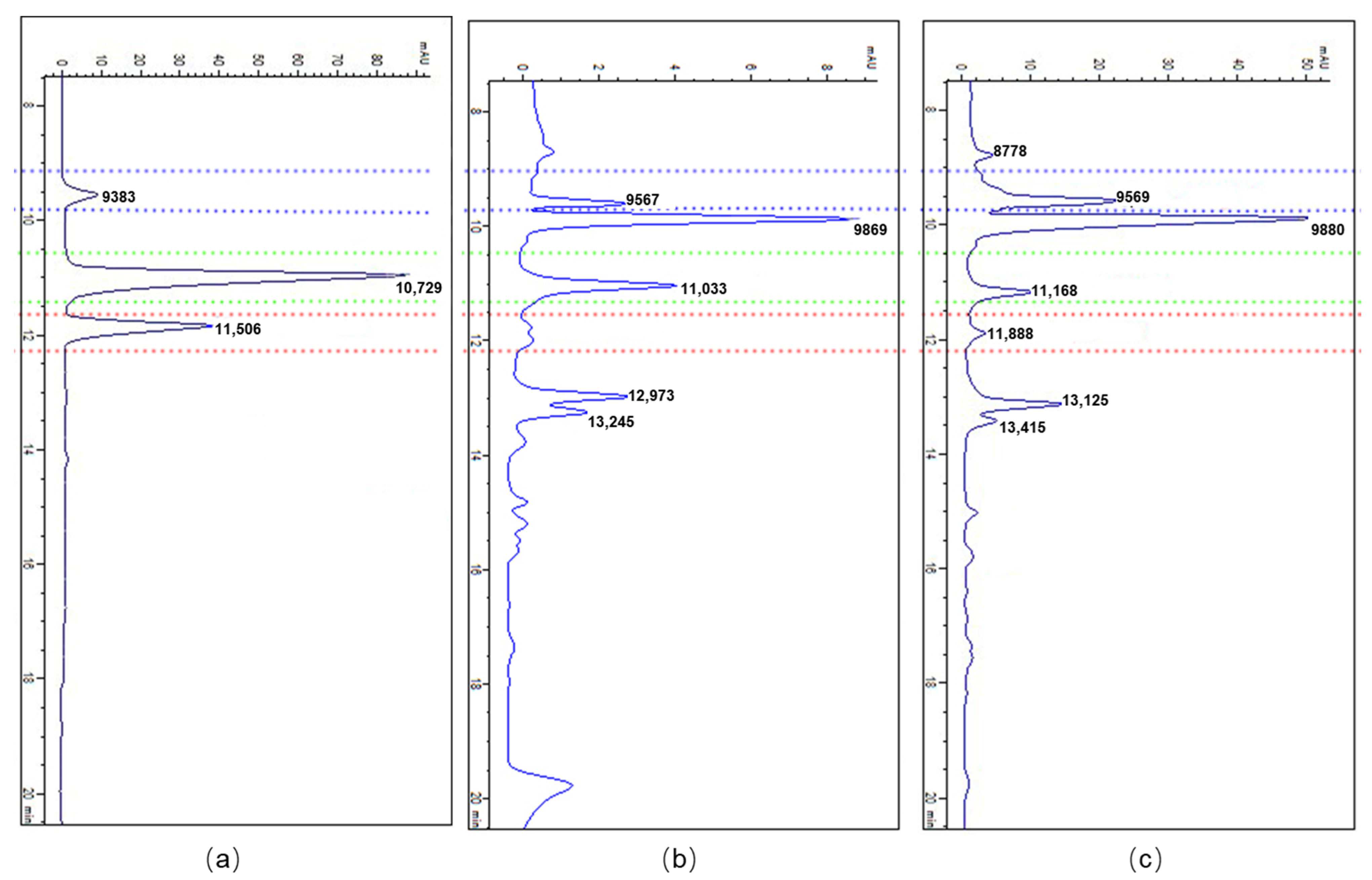
2.3. Accumulation of BPAs
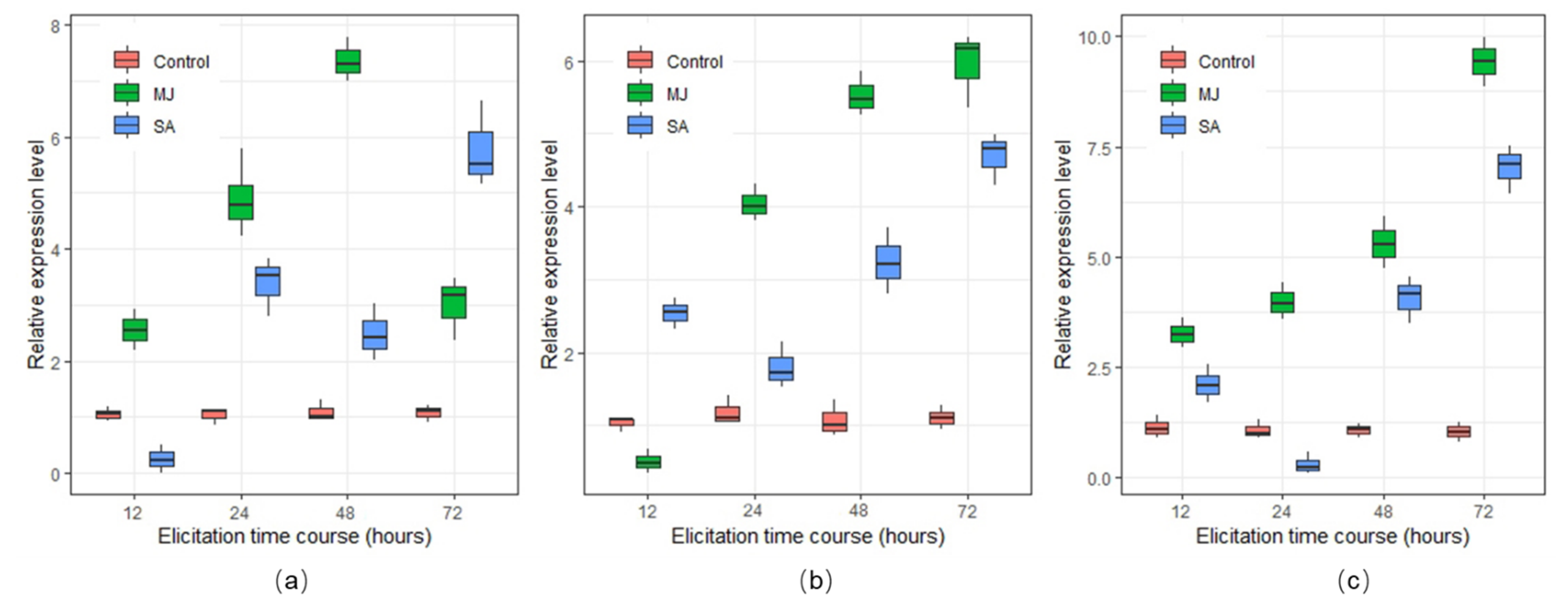
2.4. Expression of BAPs Biosynthetic Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Seed Germination
4.2. Callus and Cell Suspension Culture
4.3. Elicitation Treatments
4.4. Measurement of Cell Growth
4.5. Metabolite Extraction and Evaluation
4.6. RNA Extraction and Transcription Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Funayama, S.; Cordell, G.A. Alkaloids: Treasury of Poisons and Medicines, 1st ed.; Academic Press: London, UK, 2014; ISBN 9780124173026. [Google Scholar]
- Herrmann, R.; Roller, J.; Polednik, C.; Schmidt, M. Effect of chelidonine on growth, invasion, angiogenesis and gene expression in head and neck cancer cell lines. Oncol. Lett. 2018, 16, 3108–3116. [Google Scholar] [CrossRef]
- Wang, X.; Feng, S.; Ding, N.; He, Y.; Li, C.; Li, M.; Ding, X.; Ding, H.; Li, J.; Wu, J.; et al. Anti-Inflammatory effects of berberine hydrochloride in an LPS-induced murine model of mastitis. Evid. Based Complement. Altern. Med. 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Zielinska, S.; Wójciak-Kosior, M.; Dziagwa-Becker, M.; Glensk, M.; Sowa, I.; Fijałkowski, K.; Ruranska-Smutnicka, D.; Matkowski, A.; Junka, A. The activity of isoquinoline alkaloids and extracts from Chelidonium majus against pathogenic bacteria. Toxins 2019, 11, 406. [Google Scholar] [CrossRef] [PubMed]
- El-Readi, M.Z.; Eid, S.; Ashour, M.L.; Tahrani, A.; Wink, M. Modulation of multidrug resistance in cancer cells by chelidonine and Chelidonium majus alkaloids. Phytomedicine 2013, 20, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Ernst, E.; Schmidt, K. Ukrain—a new cancer cure? A systematic review of randomised clinical trials. BMC Cancer 2005, 5, 69. [Google Scholar] [CrossRef] [PubMed]
- Moscatiello, R.; Baldan, B.; Navazio, L. Plant Cell Suspension Cultures. In Methods in Molecular Biology; Springer: Cham, Switzerland, 2013; Volume 953, p. 297. ISBN 9781627031516. [Google Scholar]
- Gabotti, D.; Locatelli, F.; Cusano, E.; Baldoni, E.; Genga, A.; Pucci, L.; Consonni, R.; Mattana, M. Cell suspensions of cannabis sativa (var. futura): Effect of elicitation on metabolite content and antioxidant activity. Molecules 2019, 24, 4056. [Google Scholar] [CrossRef]
- Skalicky, M.; Kubes, J.; Hejnak, V.; Tumova, L.; Martinkova, J.; Martin, J.; Hnilickova, H. Isoflavones production and possible mechanism of their exudation in genista tinctoria L. suspension culture after treatment with vanadium compounds. Molecules 2018, 23, 1619. [Google Scholar] [CrossRef]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Narayani, M.; Srivastava, S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochem. Rev. 2017, 16, 1227–1252. [Google Scholar] [CrossRef]
- Yang, D.; Fang, Y.; Xia, P.; Zhang, X.; Liang, Z. Diverse responses of tanshinone biosynthesis to biotic and abiotic elicitors in hairy root cultures of Salvia miltiorrhiza and Salvia castanea Diels f. tomentosa. Gene 2018, 643, 61–67. [Google Scholar] [CrossRef]
- Ochoa-Villarreal, M.; Howat, S.; Hong, S.; Jang, M.O.; Jin, Y.-W.; Lee, E.-K.; Loake, G.J. Plant cell culture strategies for the production of natural products. BMB Rep. 2016, 49, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Strnad, M. Jasmonates: News on Occurrence, Biosynthesis, Metabolism and Action of an Ancient Group of Signaling Compounds. Int. J. Mol. Sci. 2018, 19, 2539. [Google Scholar] [CrossRef]
- Gundlach, H.; Muller, M.J.; Kutchan, T.M.; Zenk, M.H. Jasmonic acid is a signal transducer in elicitor-induced plant cell cultures. Proc. Natl. Acad. Sci. USA 1992, 89, 2389–2393. [Google Scholar] [CrossRef] [PubMed]
- Wiesel, L.; Davis, J.L.; Milne, L.; Fernandez, V.R.; Herold, M.B.; Williams, J.M.; Morris, J.; Hedley, P.E.; Harrower, B.; Newton, A.C.; et al. A transcriptional reference map of defence hormone responses in potato. Sci. Rep. 2015, 5, 15229. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.B.; Jayakodi, M.; Lee, Y.S.; Nguyen, V.B.; Park, H.; Koo, H.J.; Choi, I.Y.; Kim, D.H.; Chung, Y.J.; Ryu, B.; et al. Identification of candidate UDP-glycosyltransferases involved in protopanaxadiol-type ginsenoside biosynthesis in Panax ginseng. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Kutchan, T. Alkaloid Biosynthesis-The Basis for Metabolic Engineering of Medicinal Plants. Plant Cell 1995, 7, 1059–1070. [Google Scholar] [CrossRef]
- Beaudoin, G.A.W.; Facchini, P.J. Benzylisoquinoline alkaloid biosynthesis in opium poppy. Planta 2014, 240, 19–32. [Google Scholar] [CrossRef]
- Verma, P.; Khan, S.A.; Mathur, A.K.; Ghosh, S.; Shanker, K.; Kalra, A. Improved sanguinarine production via biotic and abiotic elicitations and precursor feeding in cell suspensions of latex-less variety of Papaver somniferum with their gene expression studies and upscaling in bioreactor. Protoplasma 2014, 251, 1359–1371. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-wenzig, E.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-H. Anticancer drug design based on plant-derived natural products. Biomed. Sci. 1999, 7360, 236–250. [Google Scholar] [CrossRef]
- Balandrin, M.F.; Klocke, J.A.; Wurtele, E.S.; Hugh, W. Natural plant chemicals: Sources of industrial and medicinal materials. Science 1985, 228, 1154–1160. [Google Scholar] [CrossRef]
- Ramirez-estrada, K.; Vidal-limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.M.; Palazon, J. Elicitation, an Effective Strategy for the Biotechnological Production of Bioactive High-Added Value Compounds in Plant Cell Factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef]
- Kaczmarek, Z. The effects of genotype, inflorescence developmental stage and induction medium on callus induction and plant regeneration in two Miscanthus species. Plant Cell Tissue Organ Cult. 2010, 102, 79–86. [Google Scholar]
- Farhadi, N.; Panahandeh, J.; Azar, A.M.; Salte, S.A. Effects of explant type, growth regulators and light intensity on callus induction and plant regeneration in four ecotypes of Persian shallot (Allium hirtifolium). Sci. Hortic. 2017, 218, 80–86. [Google Scholar] [CrossRef]
- Ahmad, N.; Faisal, M.; Anis, M.; Aref, I.M. In vitro callus induction and plant regeneration from leaf explants of Ruta graveolens L. S. Afr. J. Bot. 2010, 76, 597–600. [Google Scholar] [CrossRef]
- Kai, G.; Dai, L.; Mei, X.; Zheng, J.; Wang, W.; Lu, Y.; Qian, Z.; Zhou, G. In vitro plant regeneration from leaf explants of Ophiorrhiza japonica. Biol. Plant. 2008, 52, 557–560. [Google Scholar] [CrossRef]
- Sitbon, F.; Sundberg, B.; Olsson, O.; Sandberg, G. Free and Conjugated Indoleacetic Acid ( IAA ) Contents in Transgenic Tobacco Plants Expressing the iaaM and iaaH IAA Biosynthesis Genes from Agrobacterium tumefaciens. Plant Physiol. 1991, 95, 480–485. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, A.; Sahin, G.; Uslu, E. Influence of nutrient media on callus induction, somatic embryogenesis and plant regeneration in selected Turkish crocus species. Biotechnol. Rep. 2016, 10, 66–74. [Google Scholar]
- Gadzovska, S.; Maury, S.; Delaunay, A.; Spasenoski, M.; Hagège, D.; Courtois, D.; Joseph, C. The influence of salicylic acid elicitation of shoots, callus, and cell suspension cultures on production of naphtodianthrones and phenylpropanoids in Hypericum perforatum L. Plant Cell Tissue Organ Cult. 2013, 113, 25–39. [Google Scholar] [CrossRef]
- Siddiqi, K.S.; Husen, A. Plant response to jasmonates: Current developments and their role in changing environment. Bull. Natl. Res. Cent. 2019, 6, 153–164. [Google Scholar] [CrossRef]
- Chronopoulou, L.; Livia, D.; Bramosanti, M.; Rosciani, R.; Palocci, C.; Pasqua, G.; Valletta, A. Microfluidic synthesis of methyl jasmonate-loaded PLGA nanocarriers as a new strategy to improve natural defenses in Vitis vinifera. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Ali, M.B.; Yu, K.-W.; Hahn, E.-J.; Paek, K.-Y. Methyl jasmonate and salicylic acid elicitation induces ginsenosides accumulation, enzymatic and non-enzymatic antioxidant in suspension culture Panax ginseng roots in bioreactors. Plant Cell Rep. 2006, 25, 613–620. [Google Scholar] [CrossRef]
- Sathish, S.; Vasudevan, V.; Karthik, S.; Pavan, G.; Manickavasagam, M. Enhanced l-Dopa production from elicited cell suspension culture of Hybanthus enneaspermus (L.) F. Muell. Plant Biotechnol. Rep. 2019, 13, 613–621. [Google Scholar] [CrossRef]
- Chen, C.T.; Kao, C.H. Osmotic stress and water stress have opposite effects on putrescine and proline production in excised rice leaves. Plant Growth Regul. 1993, 13, 197–202. [Google Scholar] [CrossRef]
- Patil, R.A.; Lenka, S.K.; Normanly, J.; Walker, E.L.; Roberts, S.C. Methyl Jasmonate Represses Growth and Affects Cell Cycle Progression in Cultured Taxus Cells. Plant Cell Rep. 2014, 33, 1479–1492. [Google Scholar] [CrossRef]
- Siddaiah, C.N.; Satyanarayana, N.R.; Mudili, V. Elicitation of resistance and associated defense responses in Trichoderma hamatum induced protection against pearl millet downy mildew pathogen. Sci. Rep. 2017, 7, srep43991. [Google Scholar] [CrossRef]
- Shi, M.; Zhou, W.; Zhang, J.; Huang, S.; Wang, H.; Kai, G. Methyl jasmonate induction of tanshinone biosynthesis in Salvia miltiorrhiza hairy roots is mediated by JASMONATE ZIM-DOMAIN repressor proteins. Sci. Rep. 2016, 6, 20919. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, W.; Zhang, Y.; Zhang, X.; Lang, D.; Zhang, X. The roles of methyl jasmonate to stress in plants. Funct. Plant Biol. 2019, 46, 197–212. [Google Scholar] [CrossRef]
- Benevenuto, R.F.; Seldal, T.; Hegland, S.J.; Rodriguez-saona, C.; Kawash, J.; Polashock, J. Transcriptional profiling of methyl jasmonate-induced defense responses in bilberry (Vaccinium myrtillus L.). BMC Plant Biol. 2019, 19, 1–18. [Google Scholar] [CrossRef]
- Mendoza, D.; Cuaspud, O.; Pablo, J.; Ruiz, O.; Arias, M. Effect of salicylic acid and methyl jasmonate in the production of phenolic compounds in plant cell suspension cultures of Thevetia peruviana. Biotechnol. Rep. 2018, 19, e00273. [Google Scholar] [CrossRef]
- Cho, H.; Young, S.; Soon, H.; Yoon, S.H.; Lee-parsons, C.W.T.; Moon, J. Synergistic effects of sequential treatment with methyl jasmonate, salicylic acid and yeast extract on benzophenanthridine alkaloid accumulation and protein expression in Eschscholtzia californica suspension cultures. J. Biotechnol. 2008, 135, 117–122. [Google Scholar] [CrossRef]
- Xu, A.; Huang, J.Z.W. Effects of ultraviolet C, methyl jasmonate and salicylic acid, alone or in combination, on stilbene biosynthesis in cell suspension cultures of Vitis vinifera L. cv. Cabernet Sauvignon. Plant Cell. Tissue Organ Cult. 2015, 122, 197–211. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, R.S.; Kumar, U.; Rekha, K.; Thakur, D.; Pal, A.K.; Singh, P.K. Expression of MYB Transcription Factor Genes in Response to Methyl Jasmonate, Salicylic Acid and Sodium Nitropruside in Selaginella bryopteris (L.) Baker. Int. J. Agric. Environ. Biotechnol. 2018, 11, 30954. [Google Scholar]
- Facchini, P.J. Alkaloid biosynthesis in plants: Biochemistry, cell biology, molecular regulation, and metabolic engineering applications. Annu. Rev. Plant Mol. Biol. 2001, 52, 29–66. [Google Scholar] [CrossRef] [PubMed]
- Gamborg, L.; Miller, R.A.; OJIMAl, K. Nutrient requirements of suspension cultures of soybean root cells. Expr. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Murasnige, T.; Folke, S. A revised medium for rapid growth and bio agsays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- De Mendiburu, F.; Yaseen, M. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.4.0. 2020. Available online: https://cran.r-project.org/package=agricolae (accessed on 24 October 2020).
- Gómez-Rubio, V. ggplot2—Elegant Graphics for Data Analysis (2nd Edition). J. Stat. Softw. 2017, 77, 3–5. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Primer Sequence (5′–3′) | Amplicon Length (bp) |
|---|---|---|---|
| CFS | GU325749.1 | F-GAAAAGGTCTTCAAGGTGTTGC R-GAGACGGTTCGATTACTAAGTCG | 150 |
| STS | GU325750.1 | F-TGGATCGGAAGTTGGAGAC R-GCCACATTTTGAAGACCTTTTCG | 187 |
| TNMT | EU882994.1 | F-GCAGTCGACGAGGATGACTGG R-GTGCATTCCGTTCACAACCCAATGATC | 107 |
| EFT-α | XM026563191.1 | F-AGATGATTCCAACCAAGCCC R-CCTTGATGACACCAACAGCA | 113 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hashemi, S.M.; Naghavi, M.R.; Bakhshandeh, E.; Ghorbani, M.; Priyanatha, C.; Zandi, P. Effects of Abiotic Elicitors on Expression and Accumulation of Three Candidate Benzophenanthridine Alkaloids in Cultured Greater Celandine Cells. Molecules 2021, 26, 1395. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051395
Hashemi SM, Naghavi MR, Bakhshandeh E, Ghorbani M, Priyanatha C, Zandi P. Effects of Abiotic Elicitors on Expression and Accumulation of Three Candidate Benzophenanthridine Alkaloids in Cultured Greater Celandine Cells. Molecules. 2021; 26(5):1395. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051395
Chicago/Turabian StyleHashemi, Seyed Mohammad, Mohammad Reza Naghavi, Esmaeil Bakhshandeh, Mehdi Ghorbani, Chanditha Priyanatha, and Peiman Zandi. 2021. "Effects of Abiotic Elicitors on Expression and Accumulation of Three Candidate Benzophenanthridine Alkaloids in Cultured Greater Celandine Cells" Molecules 26, no. 5: 1395. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051395