
Biochemical Properties of Tyrosinase from Aspergillus terreus and Penicillium copticola; Undecanoic Acid from Aspergillus flavus, an Endophyte of Moringa oleifera, Is a Novel Potent Tyrosinase Inhibitor

Abstract
:1. Introduction
2. Materials and Methods
2.1. Screening for the Potent Tyrosinase Producing Fungal Isolates
2.2. Tyrosinase Activity and Concentration
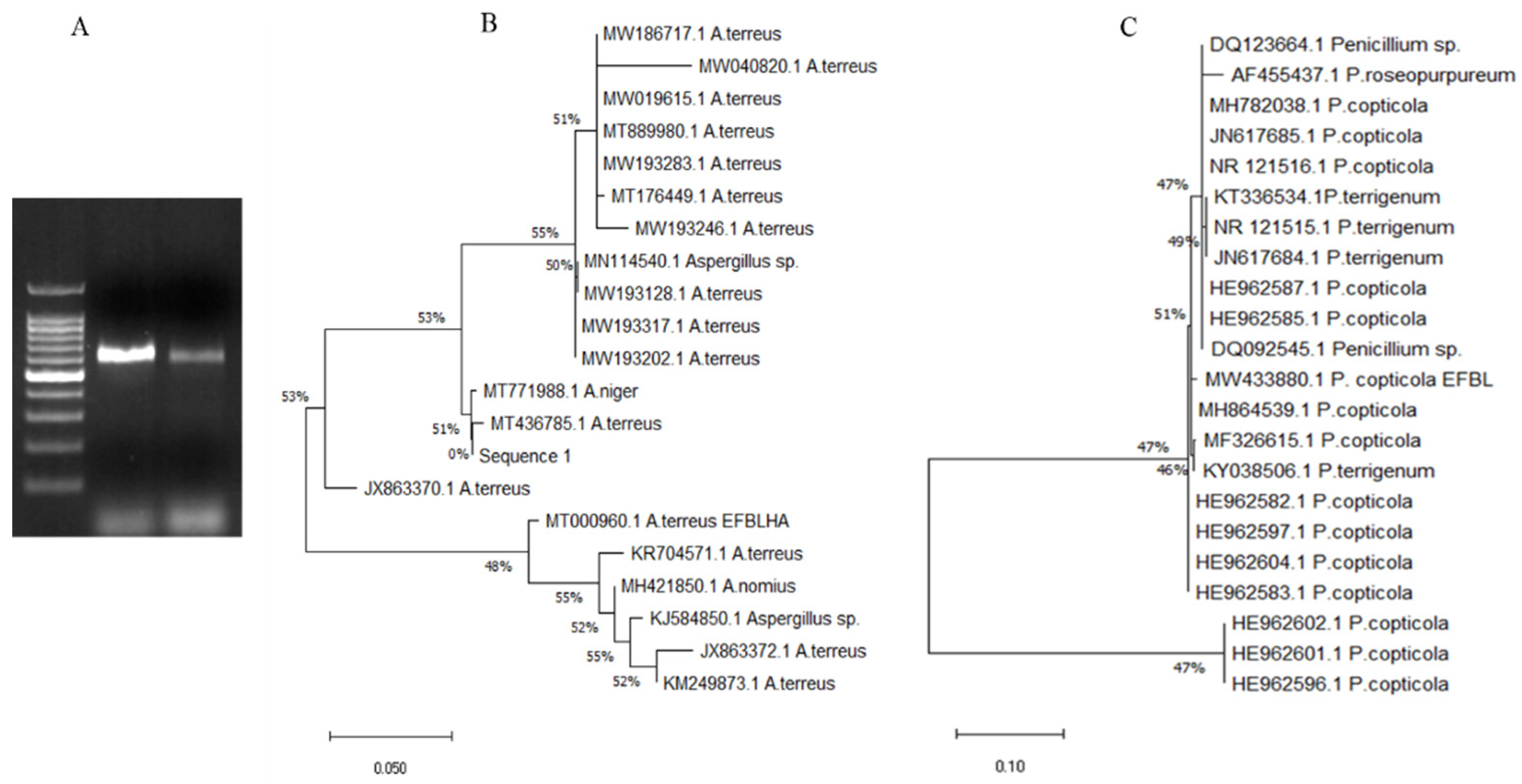
2.3. Morphological and Molecular Identification of the Potent Fungal Isolates
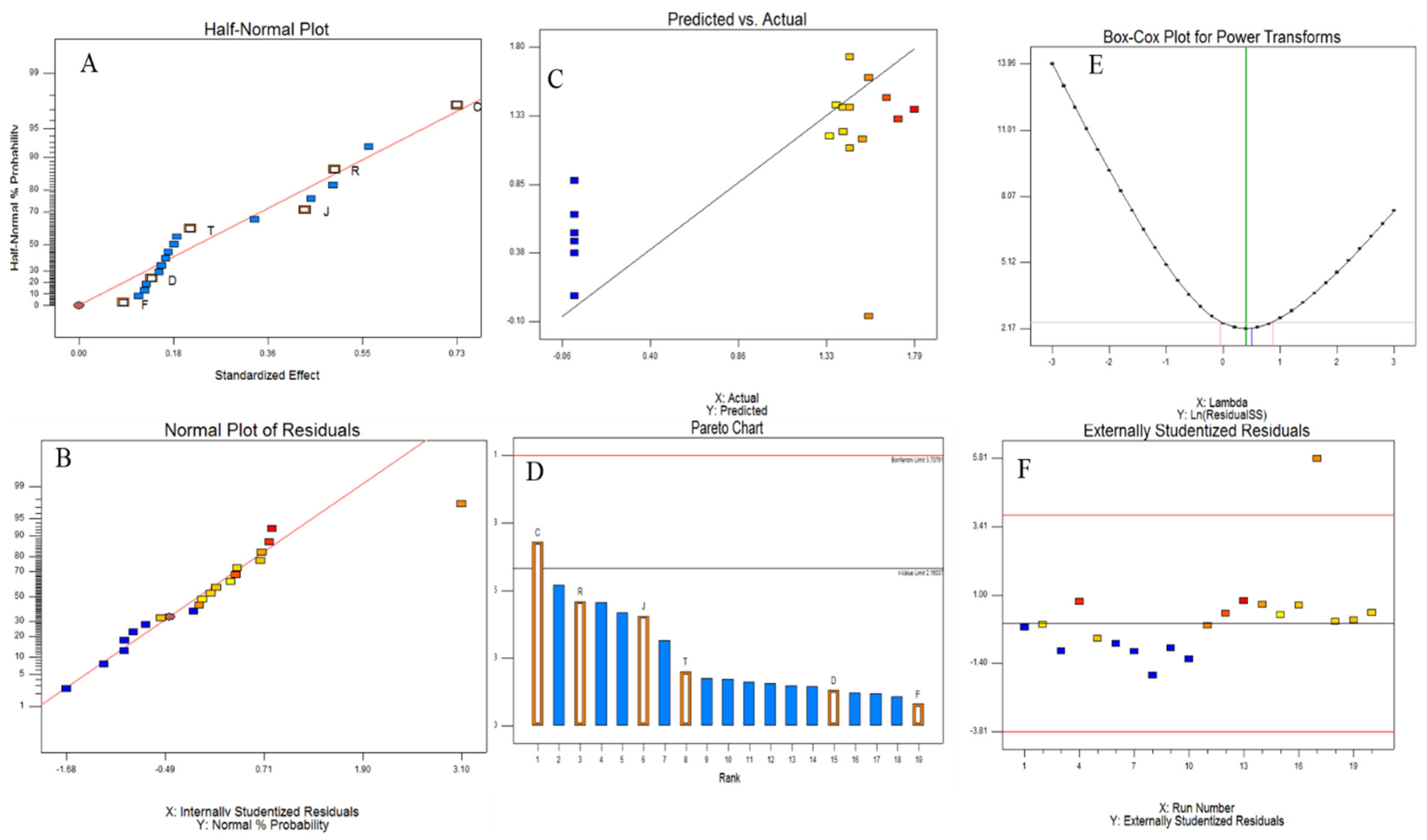
2.4. Bioprocess Optimization of the Potent Fungal Isolate by Two-Factorial Plackett–Burman Design
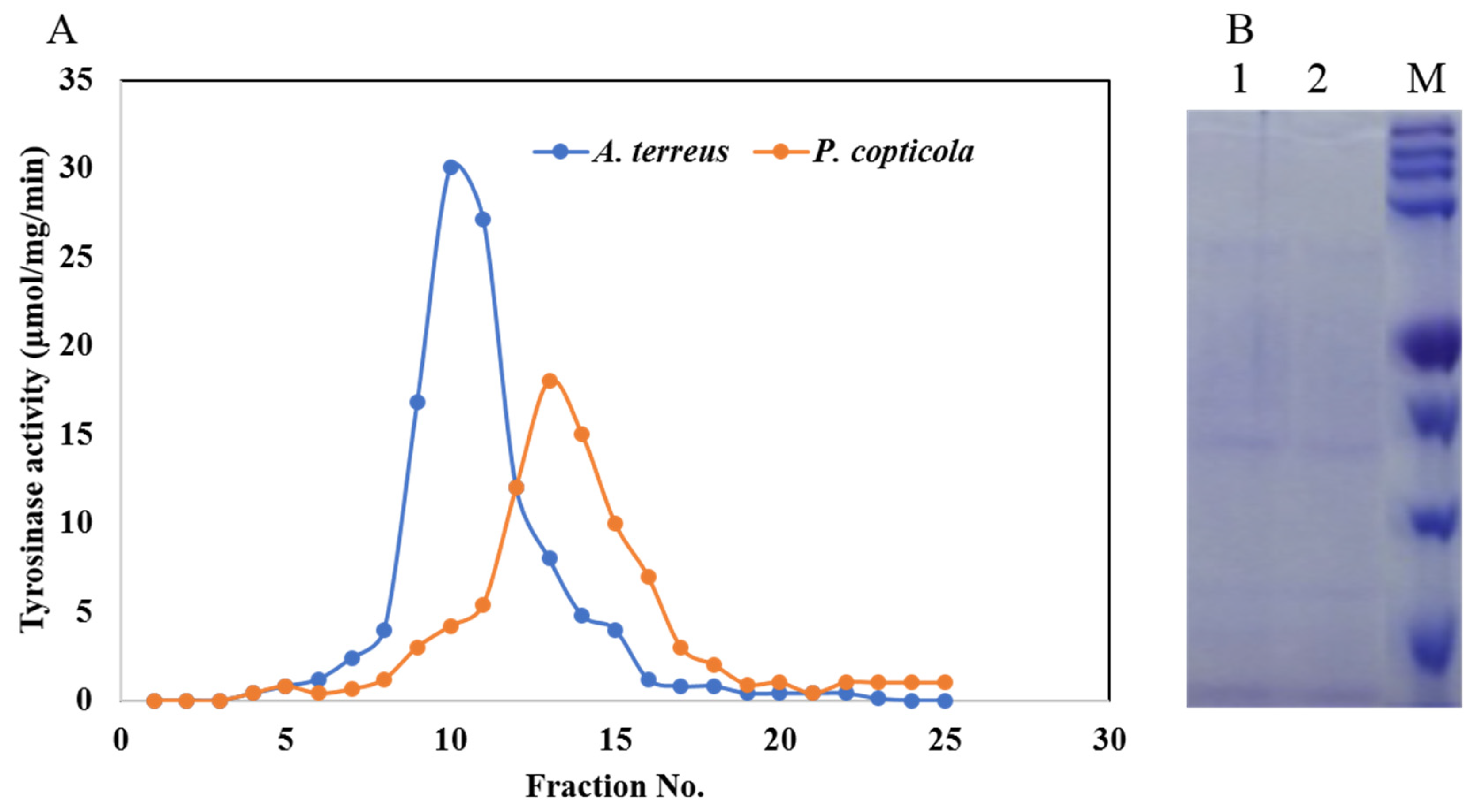
2.5. Purification, Molecular Mass, and Subunit Structure of Tyrosinase
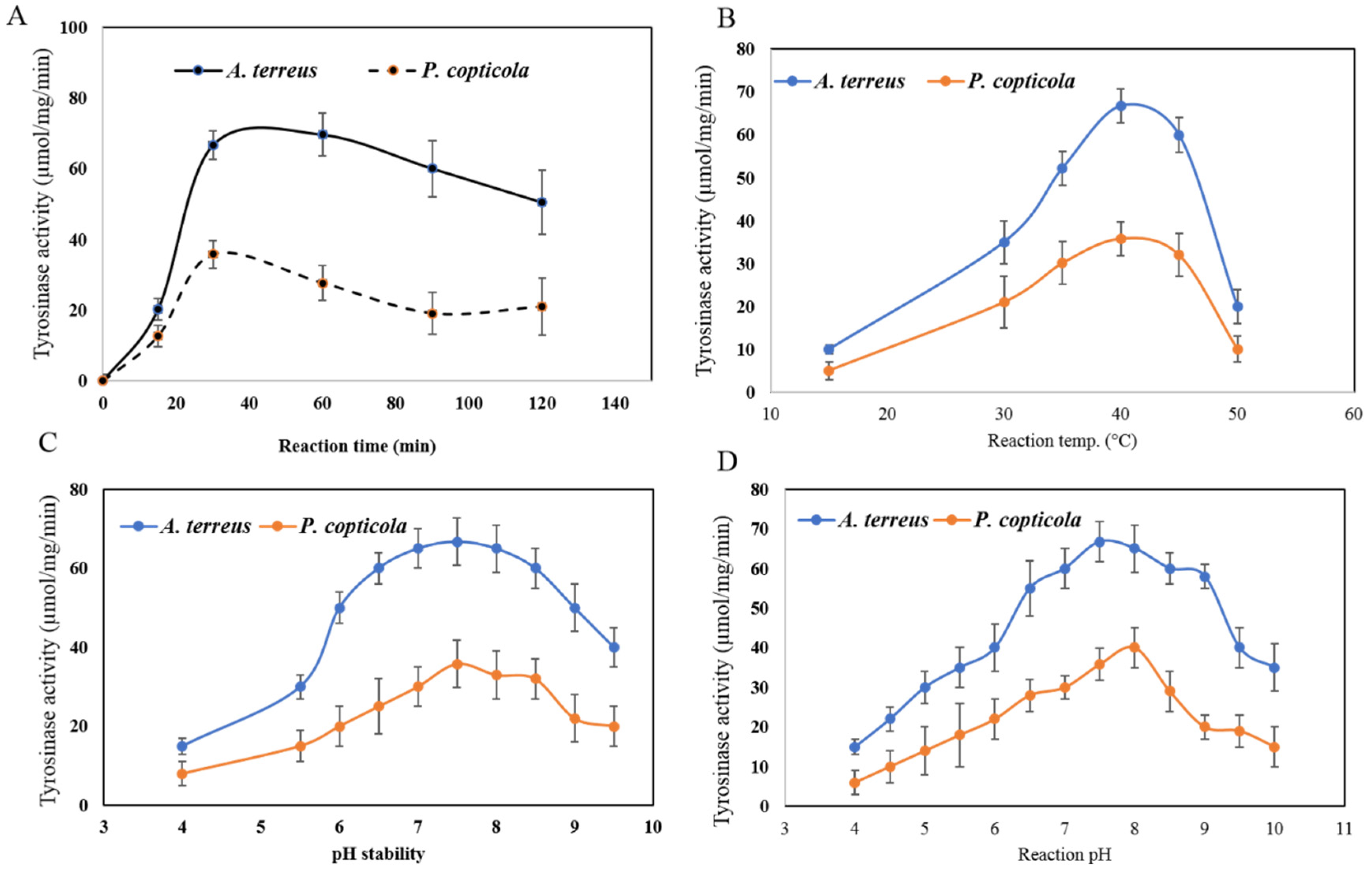
2.6. Biochemical Properties of the Purified Tyrosinase from the Potent Fungal Isolates
2.7. Effect of Various Inhibitors and Fungal Extracts
2.8. Effect of Kojic Acid, and Extracts of Different Endophytic Fungal Isolates
2.9. Fractionation and GC-MS Analysis of the Bioactive Fractions
2.10. Deposition of the Fungal Isolates
2.11. Statistical Analyses
3. Results and Discussion
3.1. Screening for the Potent Tyrosinase Producing Fungi
3.2. Molecular Confirmation of the Identity of the Potent Tyrosinase Producing Fungi
3.3. Purification, Molecular Subunit Structure of Tyrosinase from A. terreus and P. copticola
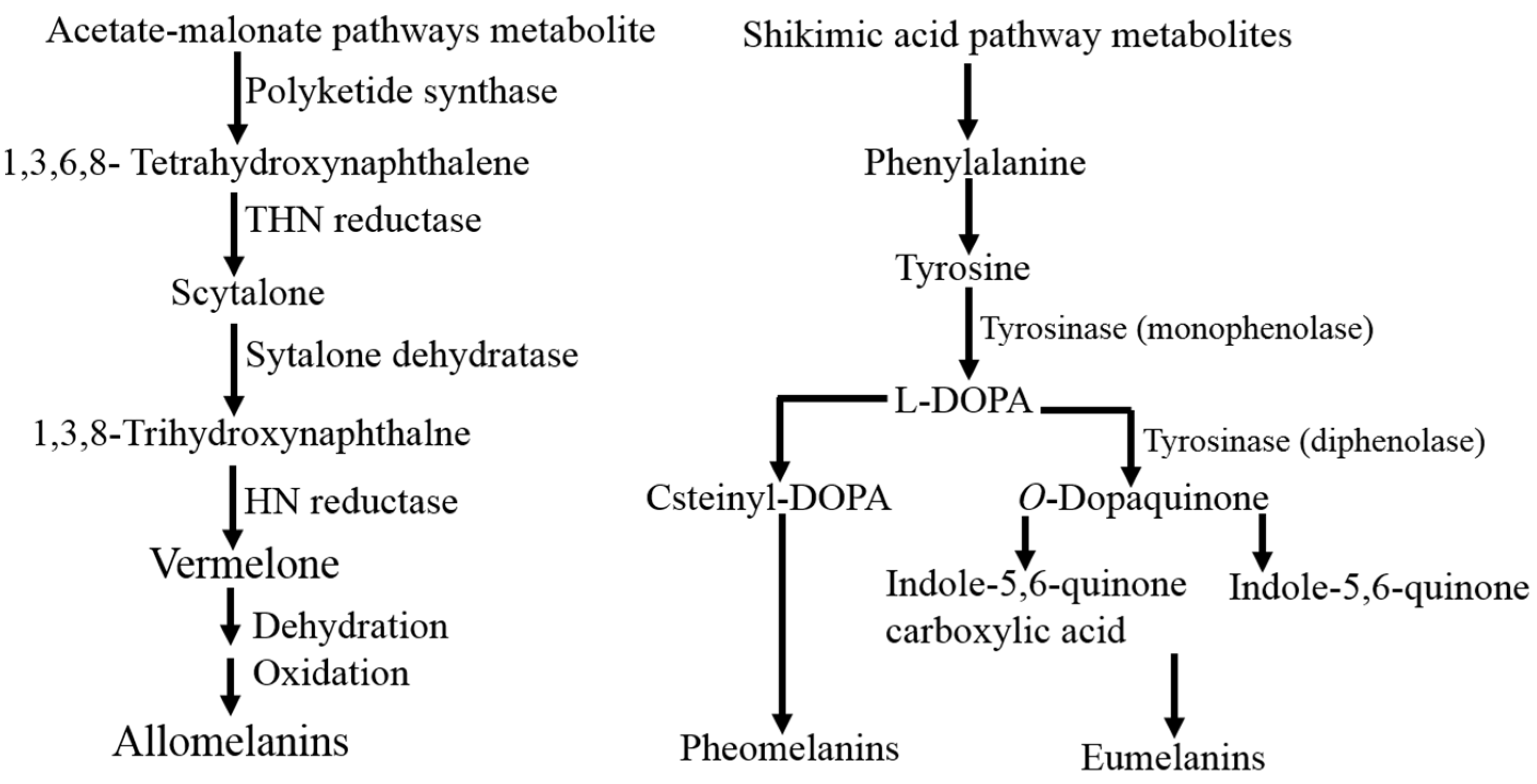
3.4. Biochemical Properties of the Purified A. terreus and P. copticola Tyrosinase
3.4.1. Reaction Time, Reaction Temperature, Thermal Stability, Reaction pH, and pH Stability
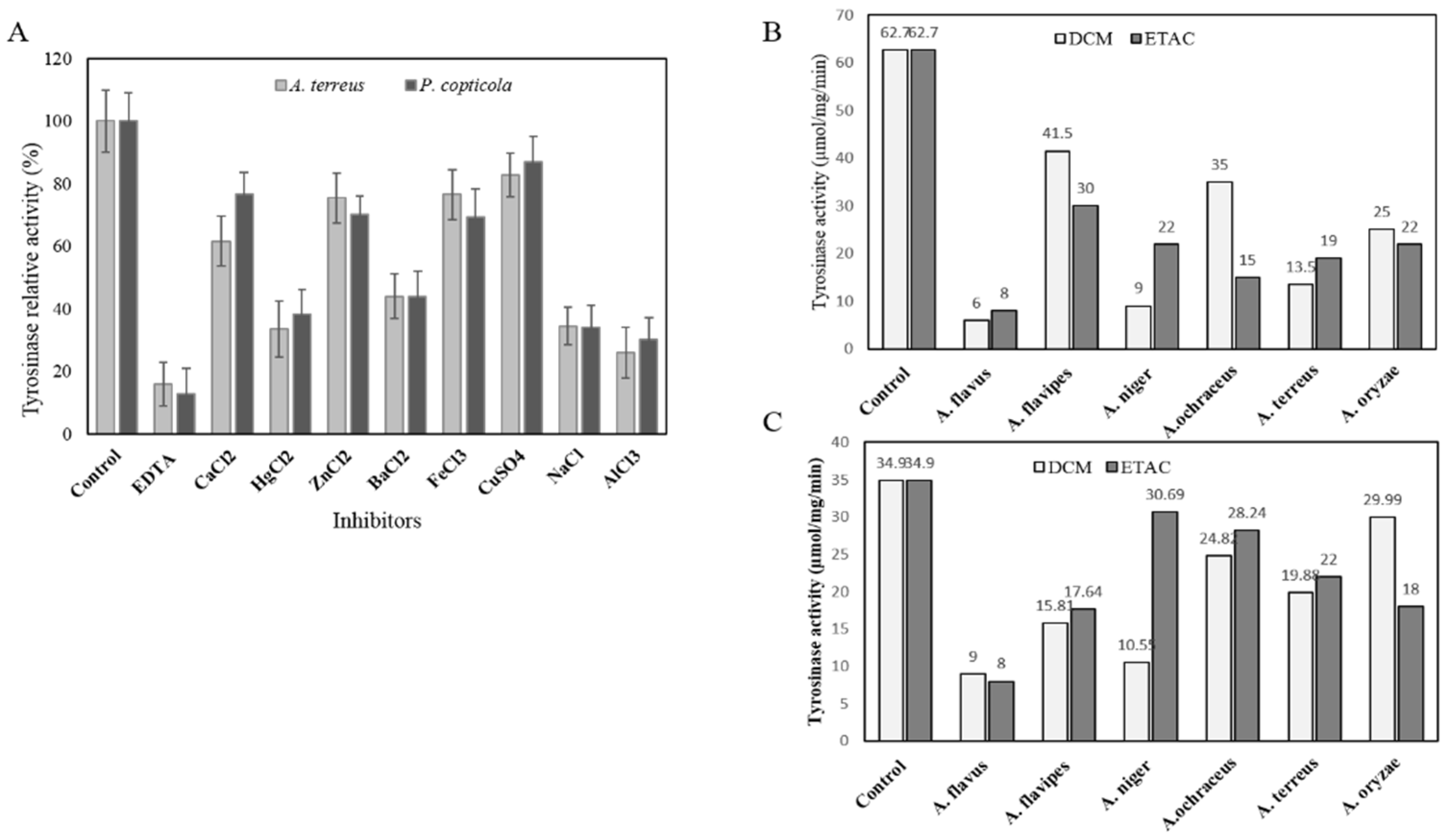
3.4.2. Effect of Various Cations Inhibitors and Extracted Compounds from Various Endophytic Fungi
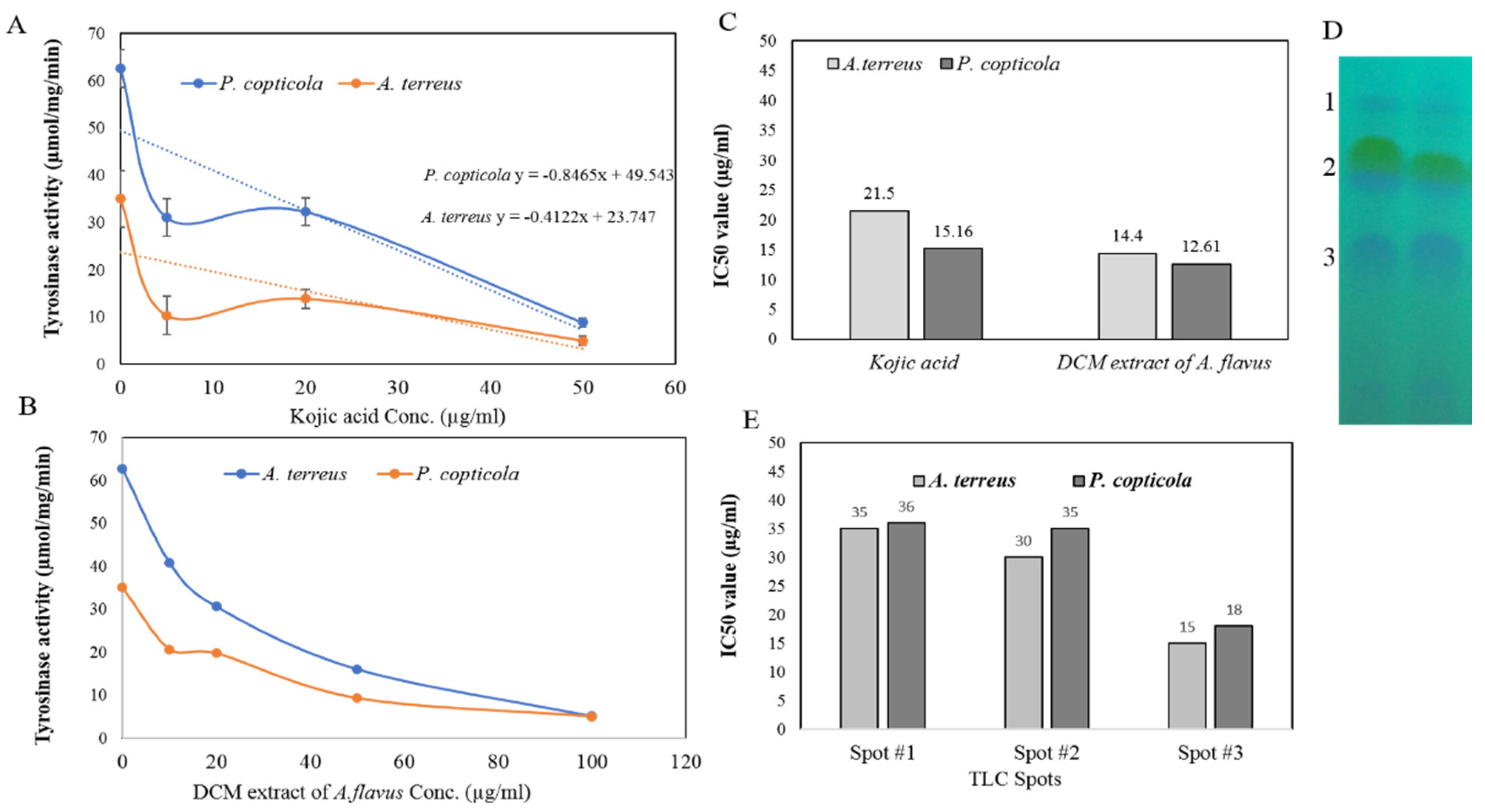
3.4.3. Kinetics of Inhibition of Tyrosinase by Kojic Acid and Dichloromethane Extracts of A. flavus
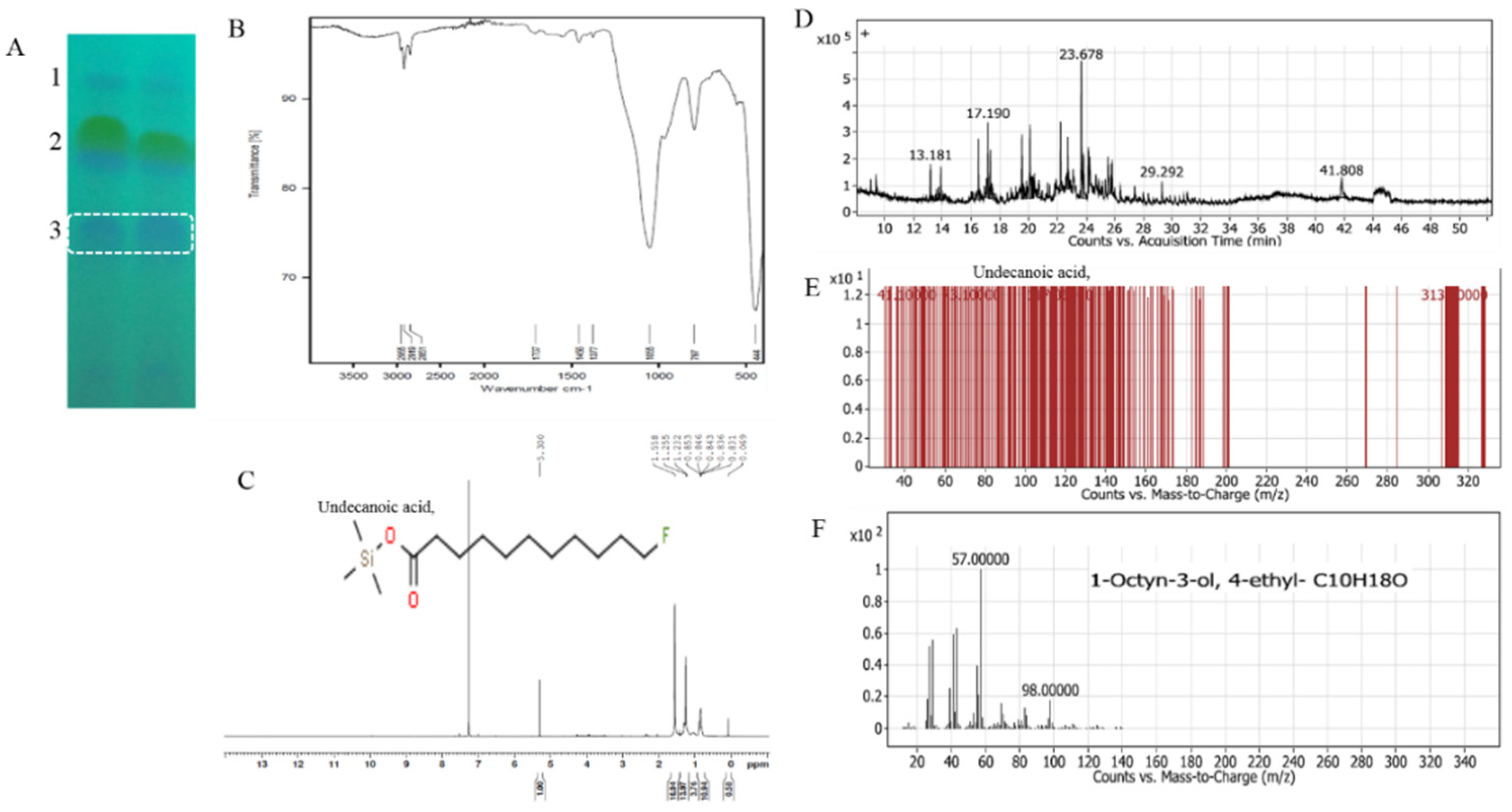
3.4.4. Chemical Identity of the Most Anti-Tyrosinase Active Fraction by FT-IR, H NMR and GC-MS Analyses
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Sánchez-Ferrer, Á.; Neptuno Rodríguez-López, J.; García-Cánovas, F.; García-Carmona, F. Tyrosinase: A comprehensive review of its mechanism. Biochim. Biophys. Acta Protein Struct. Mol. 1995, 1247, 1–11. [Google Scholar] [CrossRef]
- Zaidi, K.U.; Ali, A.S.; Ali, S.A.; Naaz, I. Microbial tyrosinases: Promising enzymes for pharmaceutical, food bioprocessing, and environmental industry. Biochem. Res. Internat. 2014, 2014, 854687. [Google Scholar] [CrossRef] [Green Version]
- Shcherba, V.V.; Babitskaya, V.G.; Kurchenko, V.P.; Ikonnikova, N.V.; Kukulyanskaya, T.A. Antioxidant properties of fungal melanin pigments. Appl. Biochem. Microbiol. 2000, 36, 491–495. [Google Scholar] [CrossRef]
- Plonka, P.M.; Grabacka, M. Melanin synthesis in microorganisms—Biotechnological and medical aspects. Acta Biochim. Pol. 2006, 53, 429–443. [Google Scholar] [CrossRef] [Green Version]
- Nosanchuk, J.D.; Casadevall, A. The contribution of melanin to microbial pathogenesis. Cell. Microbiol. 2003, 5, 203–223. [Google Scholar] [CrossRef]
- Butler, M.J.; Day, A.W. Fungal melanins: A review. Can. J. Microbiol. 1998, 44, 1115–1136. [Google Scholar] [CrossRef]
- Kim, Y.J.; Uyama, H. Tyrosinase inhibitors from natural and synthetic sources: Structure, inhibition mechanism and perspective for the future. Cell. Mol. Life Sci. 2005, 62, 1707–1723. [Google Scholar] [CrossRef] [PubMed]
- Mohania, D.; Chandel, S.; Kumar, P.; Verma, V.; Digvijay, K.; Tripathi, D.; Choudhury, K.; Mitten, S.K.; Shah, D. Ultraviolet Radiations: Skin Defense-Damage Mechanism. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2017; Volume 996, pp. 71–87. [Google Scholar]
- Øyehaug, L.; Plahte, E.; Våge, D.I.; Omholt, S.W. The regulatory basis of melanogenic switching. Journal of Theoretical Biology. J. Theor. Biol 2002, 215, 449–468. [Google Scholar] [CrossRef]
- Jacobson, E.S. Pathogenic roles for fungal melanins. Clin. Microbiol. Rev. 2000, 13, 708–717. [Google Scholar] [CrossRef]
- Meredith, P.; Sarna, T. The physical and chemical properties of eumelanin. Pigment Cell Res. 2006, 19, 572–594. [Google Scholar] [CrossRef]
- Fernandes, M.S.; Kerkar, S. Microorganisms as a source of tyrosinase inhibitors: A review. Ann. Microbiol. 2017, 67, 343–358. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Hassan, M.N.; Nada, H.M.S. Purification, immobilization, and biochemical characterization of l-arginine deiminase from thermophilic Aspergillus fumigatus KJ434941: Anticancer activity in vitro. Biotechnol. Prog. 2015, 31, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [Green Version]
- Lerch, K.; Ettlinger, L. Purification and Characterization of a Tyrosinase from Streptomyces glaucescens. Eur. J. Biochem. 1972, 31, 427–437. [Google Scholar] [CrossRef]
- Kebeish, R.; El-Sayed, A.; Fahmy, H.; Abdel-Ghany, A. Molecular cloning, biochemical characterization, and antitumor properties of a novel l-asparaginase from Synechococcus elongatus PCC6803. Biochemistry 2016, 81, 1173–1181. [Google Scholar] [CrossRef]
- Arafa, A.M.; Abdel-Ghany, A.E.; El-Dahmy, S.I.; Abdelaziz, S.; El-Ayouty, Y.; El-Sayed, A.S.A. Purification and Characterization of Anabaena flos-aquae Phenylalanine Ammonia-Lyase as a Novel Approach for Myristicin Biotransformation. JMB 2020, 30, 622–632. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Ali, D.M.I.; Yassin, M.A.; Zayed, R.A.; Ali, G.S. Sterol inhibitor “Fluconazole” enhance the Taxol yield and molecular expression of its encoding genes cluster from Aspergillus flavipes. Process Biochem. 2019, 76, 55–67. [Google Scholar] [CrossRef]
- El-Sayed, A.S.; Khalaf, S.A.; Aziz, H.A. Characterization of homocysteine γ-lyase from submerged and solid cultures of Aspergillus fumigatus ASH (JX006238). JMB 2013, 23, 499–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-sayed, A.S.A.; Shad, G. Aspergillus flavipes is a novel e fficient biocontrol agent of Phytophthora parasitica. Biol. Control 2020, 140, 104072. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Akbar, A.; Iqrar, I.; Ali, R.; Norman, D.; Brennan, M.; Ali, G.S. A glucanolytic Pseudomonas sp. associated with Smilax bona-nox L. displays strong activity against Phytophthora parasitica. Microbiol. Res. 2018, 207, 140–152. [Google Scholar] [CrossRef]
- El-Sayed, A.S.; Shindia, A.A.; Zaher, Y. L-Amino acid oxidase from filamentous fungi: Screening and optimization. Ann. Microbiol. 2012, 62, 773–784. [Google Scholar] [CrossRef]
- El-Sayed, A.S.; Shindia, A.A.; Diab, A.A.; Rady, A.M. Purification and immobilization of l-arginase from thermotolerant Penicillium chrysogenum KJ185377.1; with unique kinetic properties as thermostable anticancer enzyme. Arch. Pharm. Res. 2014, 1–10. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; Shindia, A.A.; Ali, G.S.; Yassin, M.A.; Hussein, H.; Awad, S.A.; Ammar, H.A. Production and bioprocess optimization of antitumor Epothilone B analogue from Aspergillus fumigatus, endophyte of Catharanthus roseus, with response surface methodology. Enzyme Microb. Technol. 2021, 143, 109718. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; Moustafa, A.H.; Hussein, H.A.; El-Sheikh, A.A.; El-Shafey, S.N.; Fathy, N.A.M.; Enan, G.A. Potential insecticidal activity of Sarocladium strictum, an endophyte of Cynanchum acutum, against Spodoptera littoralis, a polyphagous insect pest. Biocatal. Agric. Biotechnol. 2020, 24, 101524. [Google Scholar] [CrossRef]
- El-Sayed, A.S.; Shindia, A.A. Characterization and immobilization of purified Aspergillus flavipes l-methioninase: Continuous production of methanethiol. J. Appl. Microbiol. 2011, 111, 54–69. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; George, N.M.; Yassin, M.A.; Alaidaroos, B.A.; Bolbol, A.A.; Mohamed, M.S.; Rady, A.M.; Aziz, S.W.; Zayed, R.A.; Sitohy, M.Z. Purification and Characterization of Ornithine Decarboxylase from Aspergillus terreus; Kinetics of Inhibition by Various Inhibitors. Molecules 2019, 24, 2756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sayed, A.S.A.; Shindia, A.A.; AbouZaid, A.A.; Yassin, A.M.; Ali, G.S.; Sitohy, M.Z. Biochemical characterization of peptidylarginine deiminase-like orthologs from thermotolerant Emericella dentata and Aspergillus nidulans. Enzyme Microb. Technol. 2019, 124. [Google Scholar] [CrossRef]
- Masamoto, Y.; Ando, H.; Murata, Y.; Shimoishi, Y.; Tada, M.; Takahata, K. Mushroom tyrosinase inhibitory activity of esculetin isolated from seeds of Euphorbia lathyris L. Biosci. Biotechnol. Biochem. 2003, 67, 631–634. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; RANDALL, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Raper, K.B.; Fennell, D.I. The Genus Aspergillus; Springer: Boston, MA, USA, 1965. [Google Scholar]
- Pitt, J.I. The genus Penicillium and its teleomorphic states Eupenicillium and Talaromyces; Academic Press: London, UK; New York, NY, USA, 1979. [Google Scholar]
- Booth, C. The Genus Fusarium. Available online: https://books.google.com.eg/books/about (accessed on 27 January 2021).
- El-Sayed, A.S.A.; Shindia, A.A.; Zeid, A.A.A.; Yassin, A.M.; Sitohy, M.Z.; Sitohy, B. Aspergillus nidulans thermostable arginine deiminase-Dextran conjugates with enhanced molecular stability, proteolytic resistance, pharmacokinetic properties and anticancer activity. Enzyme Microb. Technol. 2019, 131, 109432. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Safan, S.; Mohamed, N.Z.; Shaban, L.; Ali, G.S.; Sitohy, M.Z. Induction of Taxol biosynthesis by Aspergillus terreus, endophyte of Podocarpus gracilior Pilger, upon intimate interaction with the plant endogenous microbes. Process Biochem. 2018, 71, 31–40. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Ruff, L.E.; Ghany, S.E.A.; Ali, G.S.; Esener, S. Molecular and Spectroscopic Characterization of Aspergillus flavipes and Pseudomonas putida l-Methionine γ-Lyase in Vitro. Appl. Biochem. Biotechnol. 2017, 181, 1513–1532. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, N.E.A.; Moawad, H.; El-Shweihy, N.M.; El-Ewasy, S.M.; Elsehemy, I.A.; Abdelwahed, N.A.M. Process development for scale-up production of a therapeutic l-asparaginase by Streptomyces brollosae NEAE-115 from shake flasks to bioreactor. Sci. Rep. 2019, 9, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Yassin, M.A.; Ibrahim, H. Coimmobilization of l-methioninase and glutamate dehydrogenase: Novel approach for l-homoalanine synthesis. Biotechnol. Appl. Biochem. 2015, 62. [Google Scholar] [CrossRef]
- El-Sayed, A.S.; Shouman, S.A.; Nassrat, H.M. Pharmacokinetics, immunogenicity and anticancer efficiency of Aspergillus flavipes l-methioninase. Enzyme Microb. Technol. 2012, 51, 200–210. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; Abdel-Azeim, S.; Ibrahim, H.M.; Yassin, M.A.; Abdel-Ghany, S.E.; Esener, S.; Ali, G.S. Biochemical stability and molecular dynamic characterization of Aspergillus fumigatus cystathionine γ-lyase in response to various reaction effectors. Enzyme Microb. Technol. 2015, 81, 31–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sayed, A.S.A. l-glutaminase production by Trichoderma koningii under solid-state fermentation. Indian J. Med. Microbiol. 2009, 49, 243–250. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, A.S.A. Purification and characterization of a new l-methioninase from solid cultures of Aspergillus flavipes. J. Microbiol. 2011, 49, 130–140. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Hassan, A.E.A.; Shindia, A.A.; Mohamed, S.G.; Sitohy, M.Z. Aspergillus flavipes methionine γ-lyase-dextran conjugates with enhanced structural, proteolytic stability and anticancer efficiency. J. Mol. Catal. B Enzym. 2016, 133, S15–S24. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Ibrahim, H.; Sitohy, M.Z. Co-immobilization of PEGylated Aspergillus flavipes l-methioninase with glutamate dehydrogenase: A novel catalytically stable anticancer consortium. Enzyme Microb. Technol. 2014, 54, 59–69. [Google Scholar] [CrossRef]
- Halaouli, S.; Asther, M.; Sigoillot, J.-C.; Hamdi, M.; Lomascolo, A. Fungal tyrosinases: New prospects in molecular characteristics, bioengineering and biotechnological applications. J. Appl. Microbiol. 2006, 100, 219–232. [Google Scholar] [CrossRef]
- Nikodinovic-Runic, J.; Martin, L.B.; Babu, R.; Blau, W.; O’Connor, K.E. Characterization of melanin-overproducing transposon mutants of Pseudomonas putida F6. FEMS Microbiol. Lett. 2009, 298, 174–183. [Google Scholar] [CrossRef] [Green Version]
- McMahon, A.M.; Doyle, E.M.; Brooks, S.; O’Connor, K.E. Biochemical characterisation of the coexisting tyrosinase and laccase in the soil bacterium Pseudomonas putida F6. Enzyme Microb. Technol. 2007, 40, 1435–1441. [Google Scholar] [CrossRef]
- Kupper, U.; Niedermann, D.M.; Travaglini, G.; Lerch, K. Isolation and Characterization of the Tyrosinase Gene from Neurospora crassa. J. Biol. Chem. 1989, 264, 17250–17258. [Google Scholar] [CrossRef]
- Nakamura, M.; Nakajima, T.; Ohba, Y.; Yamauchi, S.; Lee, B.R.; Ichishima, E. Identification of copper ligands in Aspergillus oryzae tyrosinase by site-directed mutagenesis. Biochem. J. 2000, 350, 537–545. [Google Scholar] [CrossRef]
- Jolley, R.L.; Nelson, R.M.; Robb, D.A. The multiple forms of mushroom tyrosinase. Structural studies on the isozymes. J. Biol. Chem. 1969, 244, 3251–3257. [Google Scholar] [CrossRef]
- Alba, L.D.; Shawkey, M.D. Melanosomes: Biogenesis, properties, and evolution of an ancient organelle. Physiol. Rev. 2019, 99, 1–19. [Google Scholar] [PubMed]
- Selinheimo, E.; NiEidhin, D.; Steffensen, C.; Nielsen, J.; Lomascolo, A.; Halaouli, S.; Record, E.; O’Beirne, D.; Buchert, J.; Kruus, K. Comparison of the characteristics of fungal and plant tyrosinases. J. Biotechnol. 2007, 130, 471–480. [Google Scholar] [CrossRef]
- Tripathi, R.K.; Devi, C.C.; Ramaiah, A. pH-dependent interconversion of two forms of tyrosinase in human skin. Biochem. J. 1988, 252, 481–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, T.; Gerwat, W.; Batzer, J.; Eggers, K.; Scherner, C.; Wenck, H.; Stäb, F.; Hearing, V.J.; Röhm, K.H.; Kolbe, L. Inhibition of Human Tyrosinase Requires Molecular Motifs Distinctively Different from Mushroom Tyrosinase. J. Investig. Dermatol. 2018, 138, 1601–1608. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, A.S.A.; Abdel-Ghany, S.E.; Ali, G.S. Genome editing approaches: Manipulating of lovastatin and taxol synthesis of filamentous fungi by CRISPR/Cas9 system. Appl. Microbiol. Biotechnol. 2017, 101, 3953–3976. [Google Scholar] [CrossRef] [PubMed]
- KAHN, V. Effect of Kojic Acid on the Oxidation of DL-DOPA, Norepinephrine, and Dopamine by Mushroom Tyrosinase. Pigment Cell Res. 1995, 8, 234–240. [Google Scholar] [CrossRef]
- Chen, M.J.; Hung, C.C.; Chen, Y.R.; Lai, S.T.; Chan, C.F. Novel synthetic kojic acid-methimazole derivatives inhibit mushroom tyrosinase and melanogenesis. J. Biosci. Bioeng. 2016, 122, 666–672. [Google Scholar] [CrossRef]
- Rahman, V.P.M.; Mukhtar, S.; Ansari, W.H.; Lemiere, G. Synthesis, stereochemistry and biological activity of some novel long alkyl chain substituted thiazolidin-4-ones and thiazan-4-one from 10-undecenoic acid hydrazide. Eur. J. Med. Chem. 2005, 40, 173–184. [Google Scholar] [CrossRef]
- Narra, N.; Kaki, S.S.; Prasad, R.B.N.; Misra, S.; Dhevendar, K.; Kontham, V.; Korlipara, P.V. Synthesis and evaluation of anti-oxidant and cytotoxic activities of novel 10-undecenoic acid methyl ester based lipoconjugates of phenolic acids. Beilstein J. Org. Chem. 2017, 13, 26–32. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Fungal Isolate | Specific Activity (μmol/mg/min) |
|---|---|---|
| 1 | Trichoderma sp. | 0.405 |
| 2 | Aspergillus terreus | 0.422 |
| 3 | Penicillium lilacinum | 0.23 |
| 4 | Pestalotia sp. | 0.24 |
| 5 | Alternaria alternata 1 | 0.17 |
| 6 | Alternaria alternata 2 | 0.177 |
| 7 | penicillium monoverticillate | 0.152 |
| 8 | Penicillium citrinum 1 | 0.112 |
| 9 | penicillium asymmetrica | 0.325 |
| 10 | Chaetomium sp. | 0.295 |
| 11 | penicillium divericata | 0.245 |
| 12 | Aspergillus ornatus | 0.312 |
| 13 | Humicola sp. | 0.185 |
| 14 | Aspergillus awamori | 0.322 |
| 15 | Penicillium citrinum 2 | 0.232 |
| 16 | Fusarium sp. | 0.26 |
| 17 | Aspergillus glaucus | 0.24 |
| 18 | Aspergillus ustus | 0.36 |
| 19 | Penicillium copticola | 0.507 |
| Variables | Level | ||
|---|---|---|---|
| −1 | 1 | ||
| X1 | Peptone (g/L) | 2 | 5 |
| X2 | Casein (g/L) | 2 | 5 |
| X3 | Ammonium nitrate (g/L) | 2 | 5 |
| X4 | Beef extract (g/L) | 2 | 5 |
| X5 | Phenylalanine (g/L) | 2 | 5 |
| X6 | Yeast extract (g/L) | 2 | 5 |
| X7 | Urea (g/L) | 3 | 6 |
| X8 | Tyrosine (g/L) | 2 | 5 |
| X9 | Asparagine (g/L) | 2 | 5 |
| X10 | Glycine (g/L) | 2 | 5 |
| X11 | Valine (g/L) | 2 | 5 |
| X12 | Cysteine (g/L) | 2 | 5 |
| X13 | Methionine (g/L) | 2 | 5 |
| X14 | Glutamic acid (g/L) | 2 | 5 |
| X15 | Sucrose (g/L) | 2 | 6 |
| X16 | Starch (g/L) | 1 | 4 |
| X17 | Glucose (g/L) | 2 | 8 |
| X18 | pH | 5 | 8 |
| X19 | Incubation time (day) | 5 | 10 |
| Run | X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | X10 | X11 | X12 | X13 | X14 | X15 | X16 | X17 | X18 | X19 | Actual Activity | Predicted Activity | Residuals |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | 0.008 | 0.075 | −0.075 |
| 2 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 1 | 1.9 | 1.39 | 0.51 |
| 3 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 0.07 | 0.9 | 0 |
| 4 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 2.9 | 1.302 | 1.598 |
| 5 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | 2.1 | 1.73 | 0.37 |
| 6 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 0.09 | 0.375 | −0.375 |
| 7 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 1 | −1 | 0.03 | 0.513 | −0.513 |
| 8 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 0.09 | 0.87 | −0.87 |
| 9 | 1 | −1 | 1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 0.1 | 0.455 | −0.455 |
| 10 | − 1 | 1 | −1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | 0.1 | 0.641 | −0.641 |
| 11 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 2.4 | 1.59 | 0.81 |
| 12 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 2.7 | 1.451 | 1.249 |
| 13 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 3.2 | 1.369 | 1.831 |
| 14 | −1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | 2.3 | 1.162 | 1.138 |
| 15 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | 1.8 | 1.18 | 0.62 |
| 16 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | 2.1 | 1.103 | 0.997 |
| 17 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | 2.4 | 0.065 | 2.335 |
| 18 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | 2 | 1.38 | 0.62 |
| 19 | 1 | 1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 2.1 | 1.381 | 0.719 |
| 20 | −1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 1 | 2 | 1.21 | 0.79 |
| Step | Aspergillus terreus | Penicillium copticola | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Total Activity (µmol/min) | Total Protein (mg) | Specific Activity (µmol/mg/min) | Fold | Yield (%) | Total Activity (µmol/min) | Total Protein (mg) | Specific Activity (µmol/mg/min) | Fold | Yield (%) | |
| Crude enzyme | 188.44 | 92.79 | 2.03 | 1 | 100 | 178.69 | 105.69 | 2.09 | 1 | 100 |
| Acetone precipitate | 147.96 | 8.94 | 16.7 | 8.19 | 83.83 | 152 | 14.16 | 10.73 | 5.14 | 85.1 |
| Sephadex G200 | 120.46 | 4.32 | 28.1 | 13.8 | 63.93 | 100.7 | 4.31 | 23.37 | 11.2 | 56.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salah Maamoun, H.; Rabie, G.H.; Shaker, I.; A. Alaidaroos, B.; El-Sayed, A.S.A. Biochemical Properties of Tyrosinase from Aspergillus terreus and Penicillium copticola; Undecanoic Acid from Aspergillus flavus, an Endophyte of Moringa oleifera, Is a Novel Potent Tyrosinase Inhibitor. Molecules 2021, 26, 1309. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051309
Salah Maamoun H, Rabie GH, Shaker I, A. Alaidaroos B, El-Sayed ASA. Biochemical Properties of Tyrosinase from Aspergillus terreus and Penicillium copticola; Undecanoic Acid from Aspergillus flavus, an Endophyte of Moringa oleifera, Is a Novel Potent Tyrosinase Inhibitor. Molecules. 2021; 26(5):1309. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051309
Chicago/Turabian StyleSalah Maamoun, Hanaa, Gamal H. Rabie, Ibrahim Shaker, Bothaina A. Alaidaroos, and Ashraf S. A. El-Sayed. 2021. "Biochemical Properties of Tyrosinase from Aspergillus terreus and Penicillium copticola; Undecanoic Acid from Aspergillus flavus, an Endophyte of Moringa oleifera, Is a Novel Potent Tyrosinase Inhibitor" Molecules 26, no. 5: 1309. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051309