
The Biological Effects of Forsythia Leaves Containing the Cyclic AMP Phosphodiesterase 4 Inhibitor Phillyrin

Abstract
:1. Introduction
2. Structures of Phillyrin and Forsythiaside
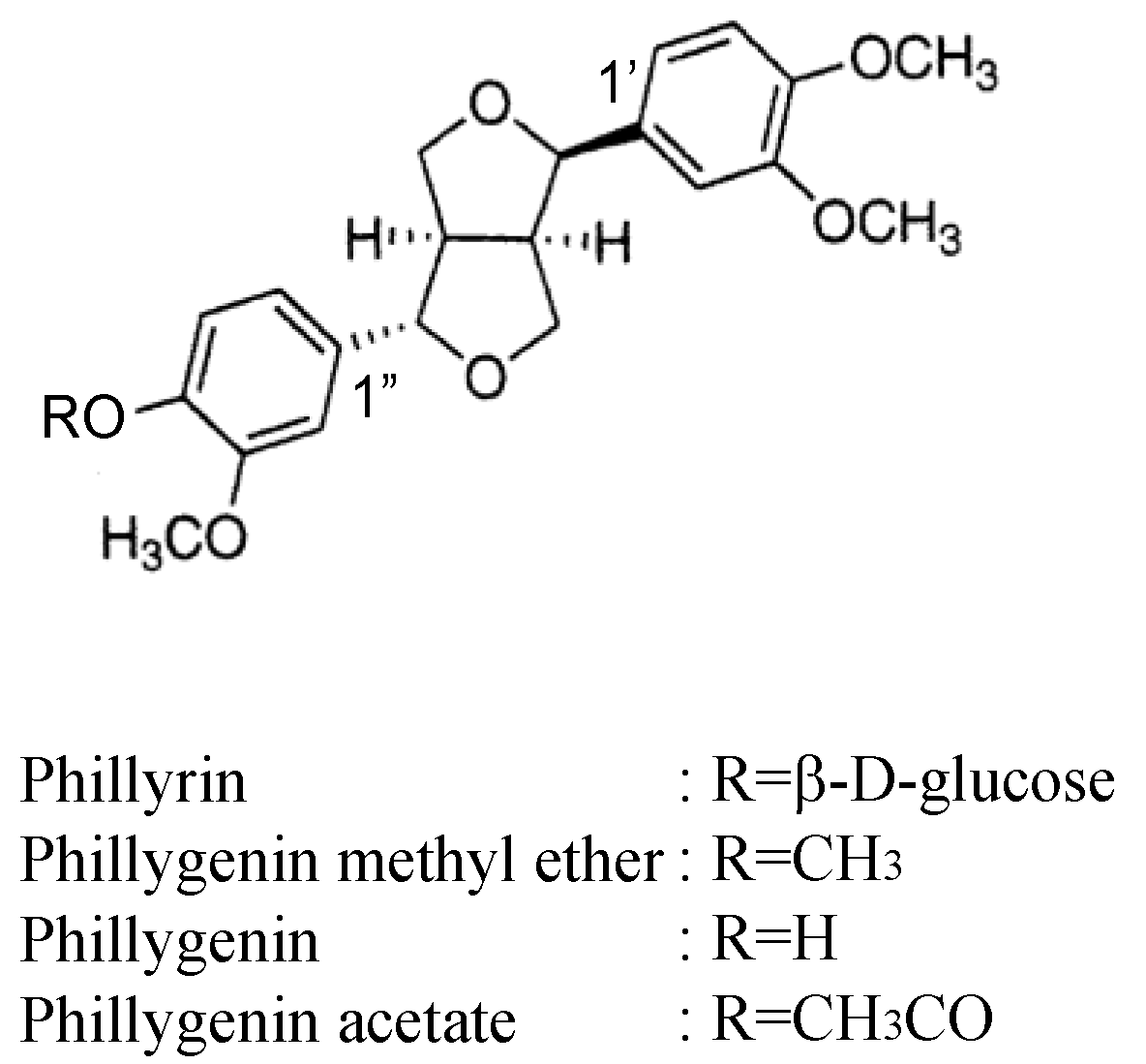
2.1. Phillyrin
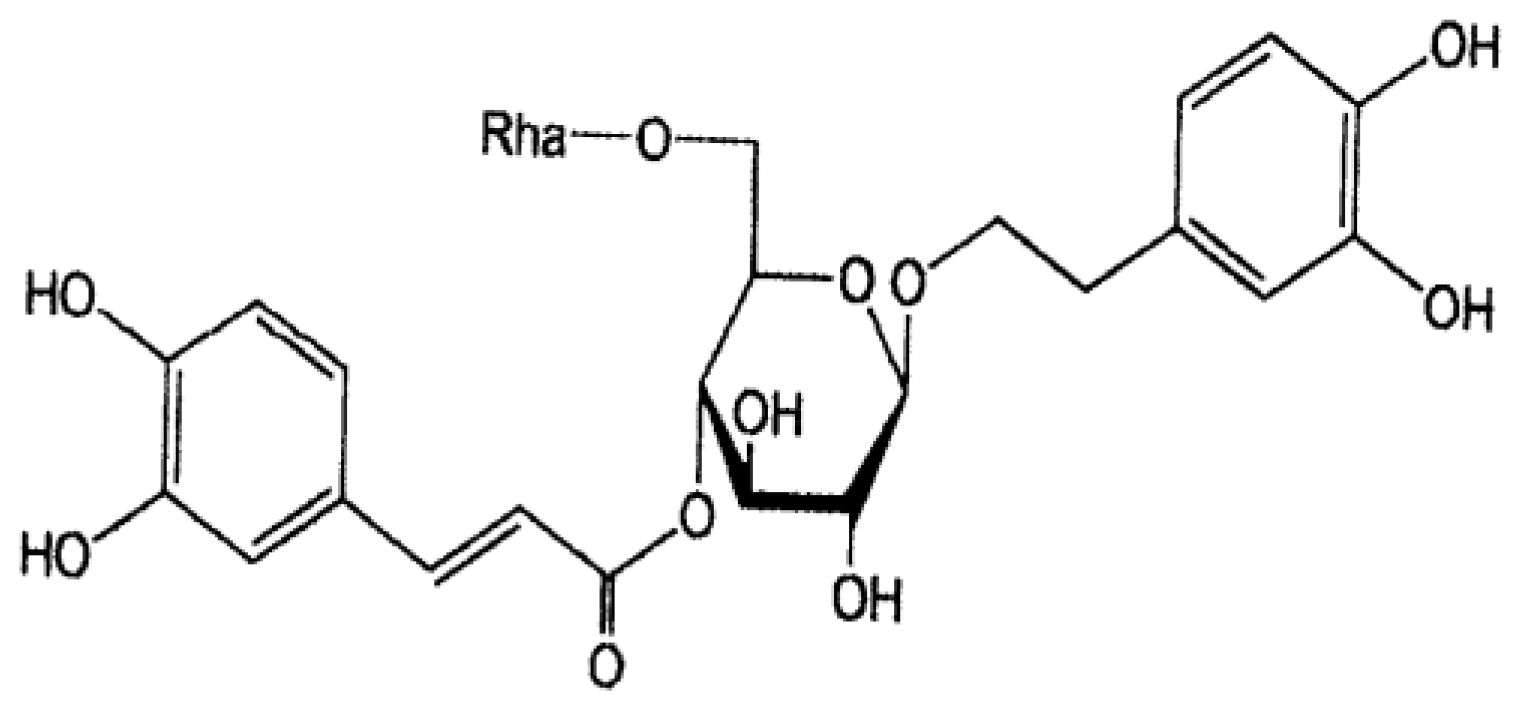
2.2. Forsythiaside
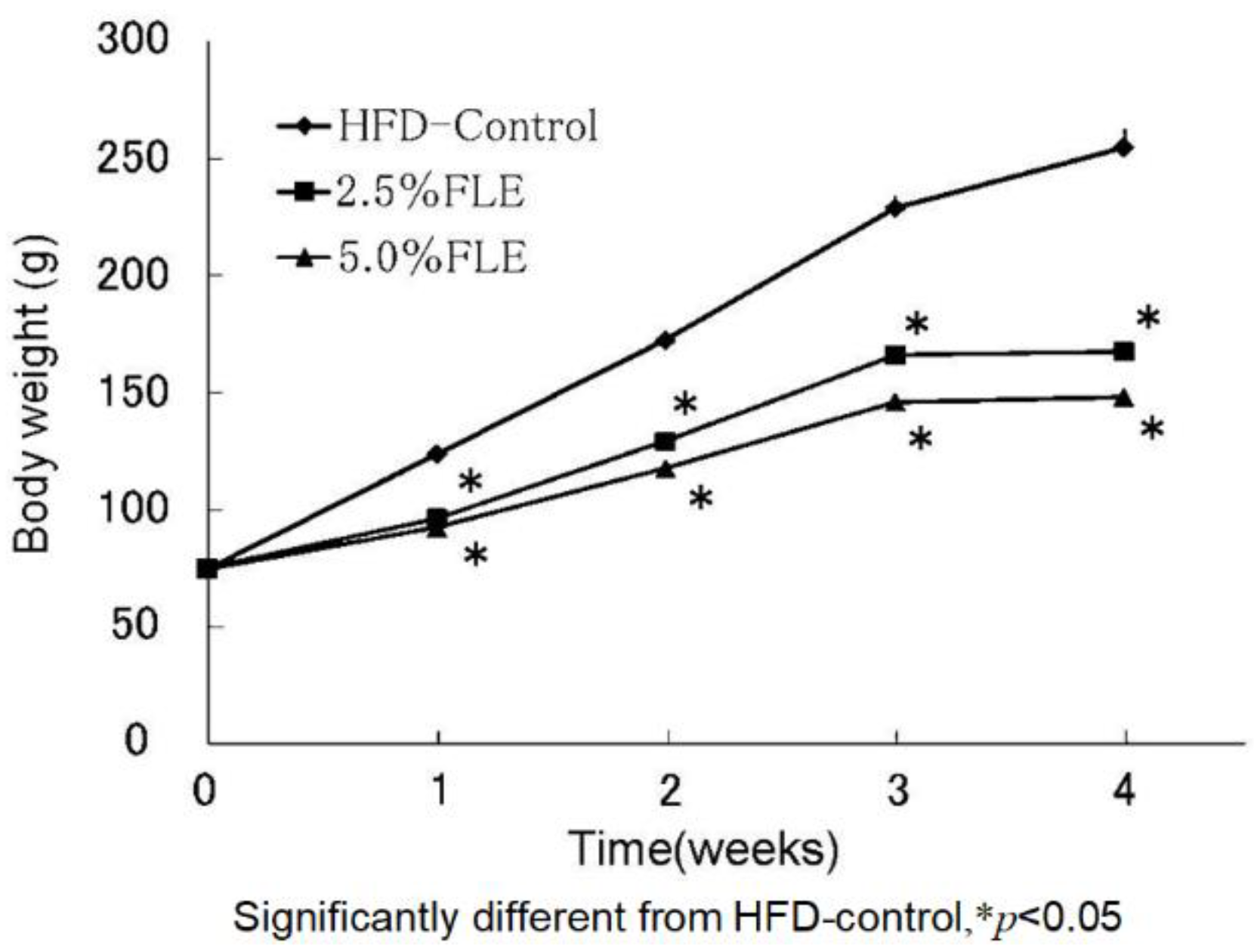
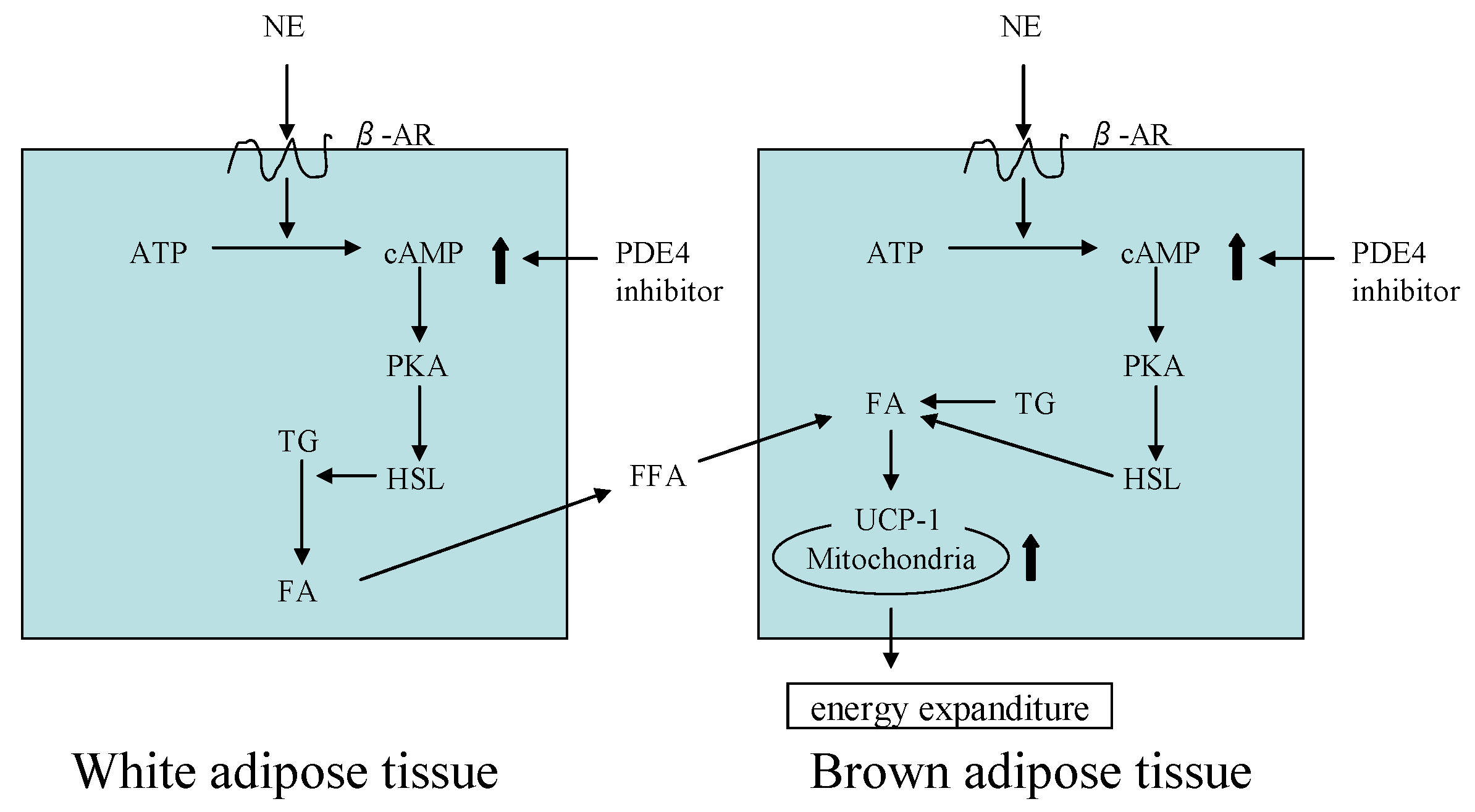
3. The Anti-Obesity Effects of Forsythia Leaves
4. Combinations of Forsythia Leaves and Other Materials
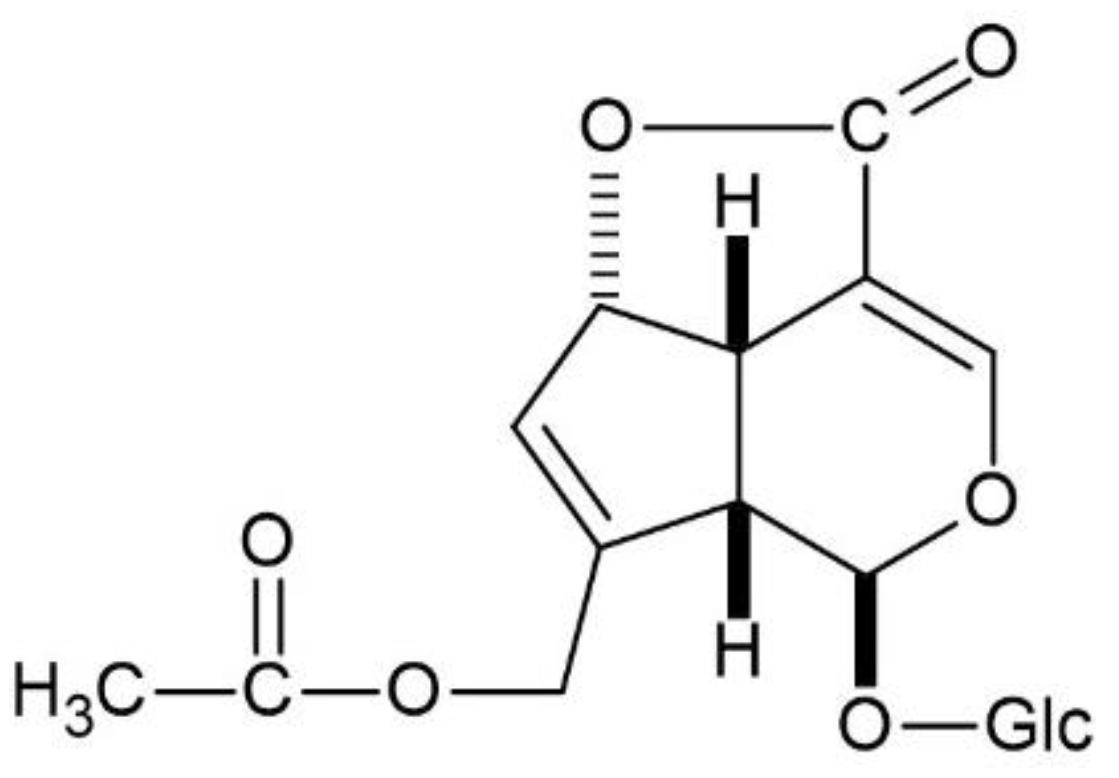
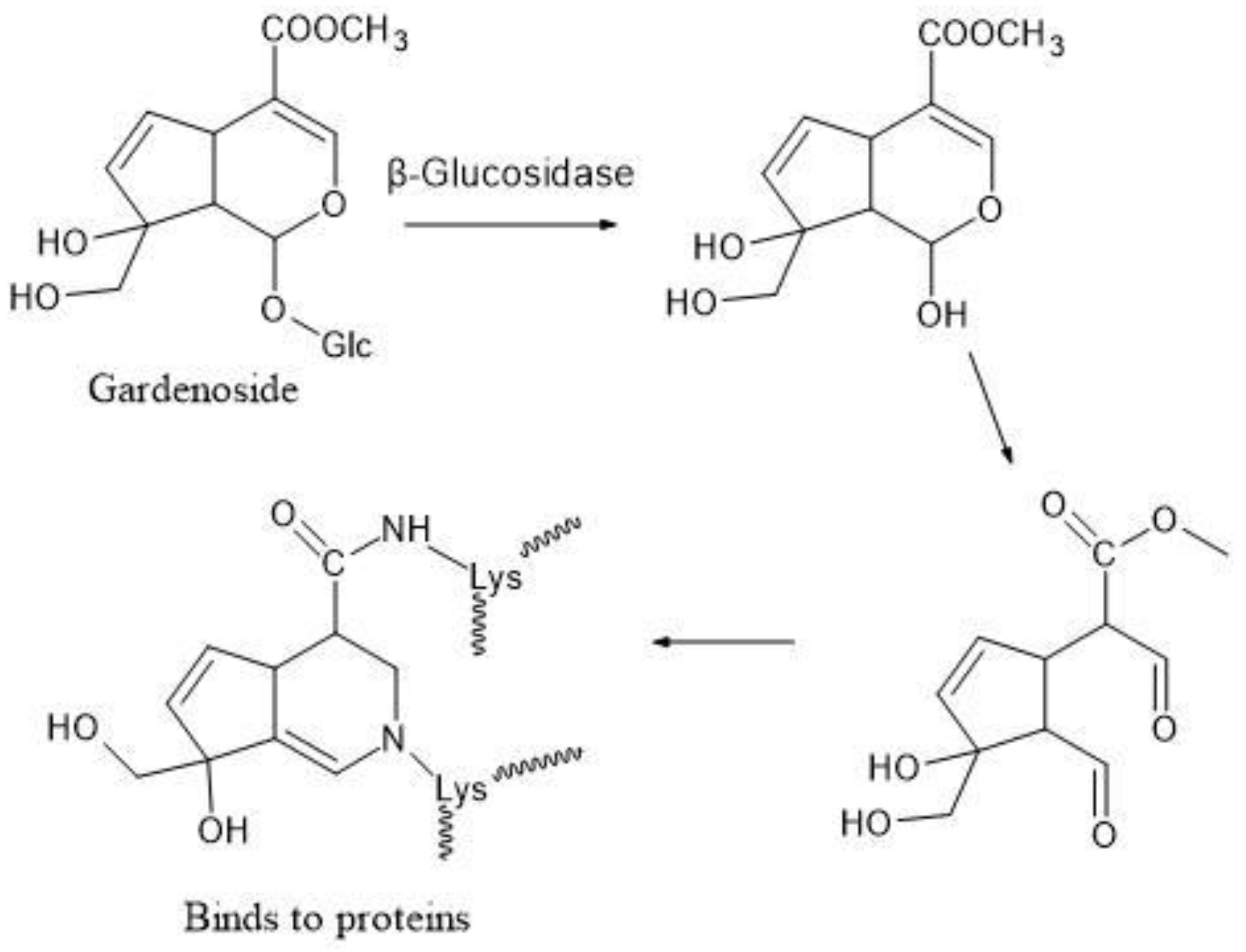
4.1. Gardenia Fruit, Glycyrrhiza, and Immature Orange
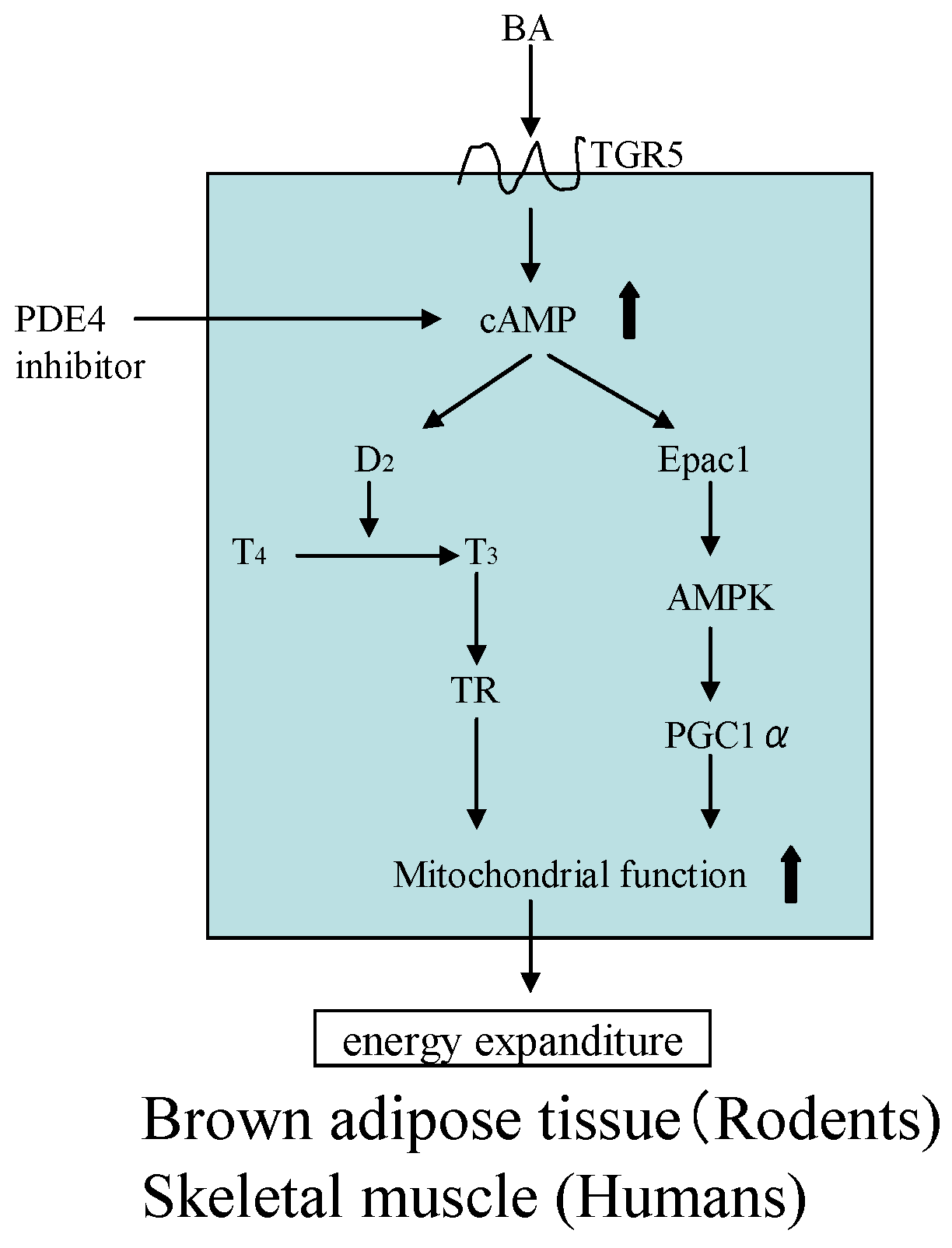
4.2. Eucommia Leaves
5. Other Biological Activities
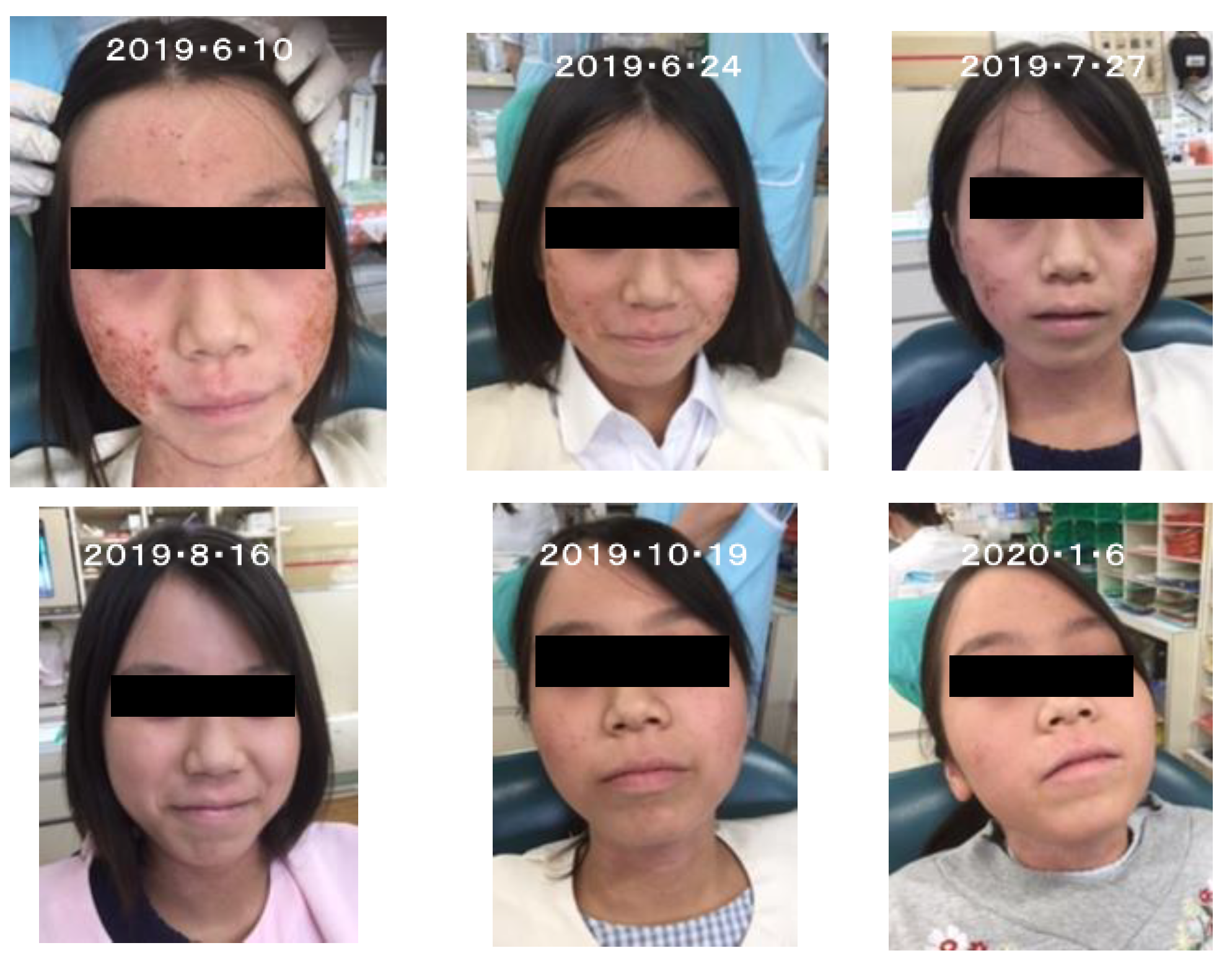
5.1. Effects for Forsythia Leaves on Atopic Dermatitis
5.2. Effects of Forsythia Leaves against Influenza A Virus Infection
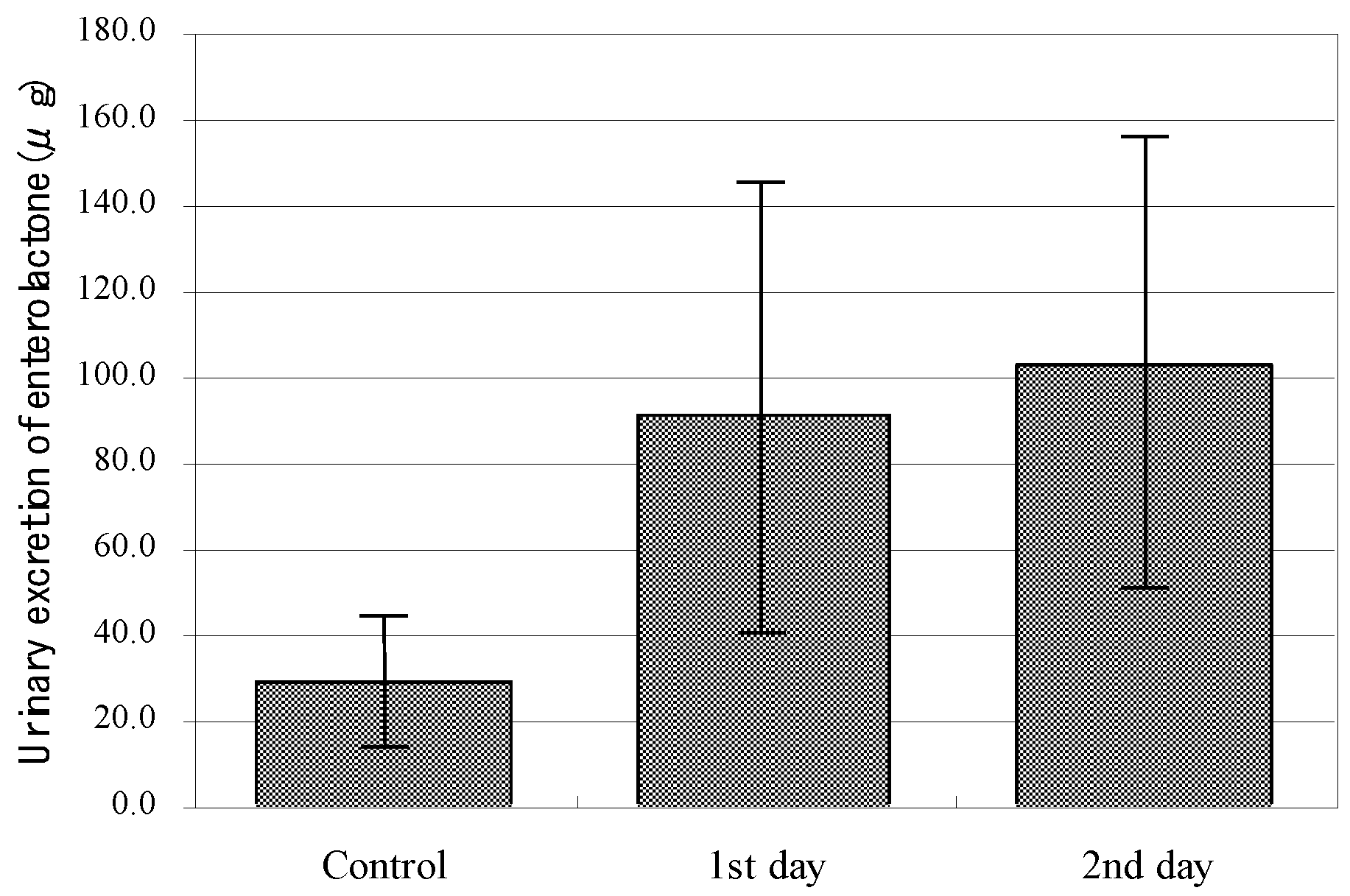
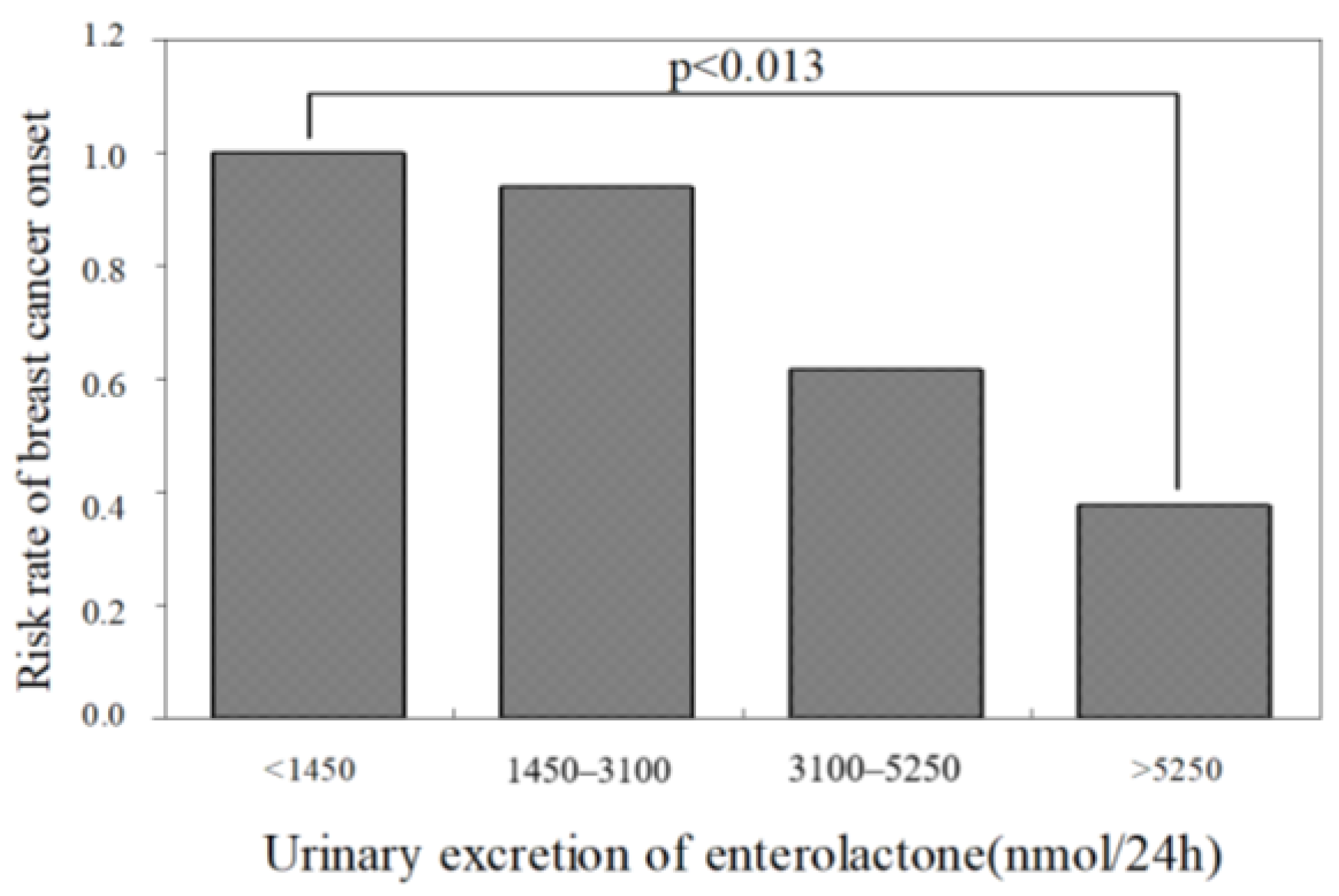
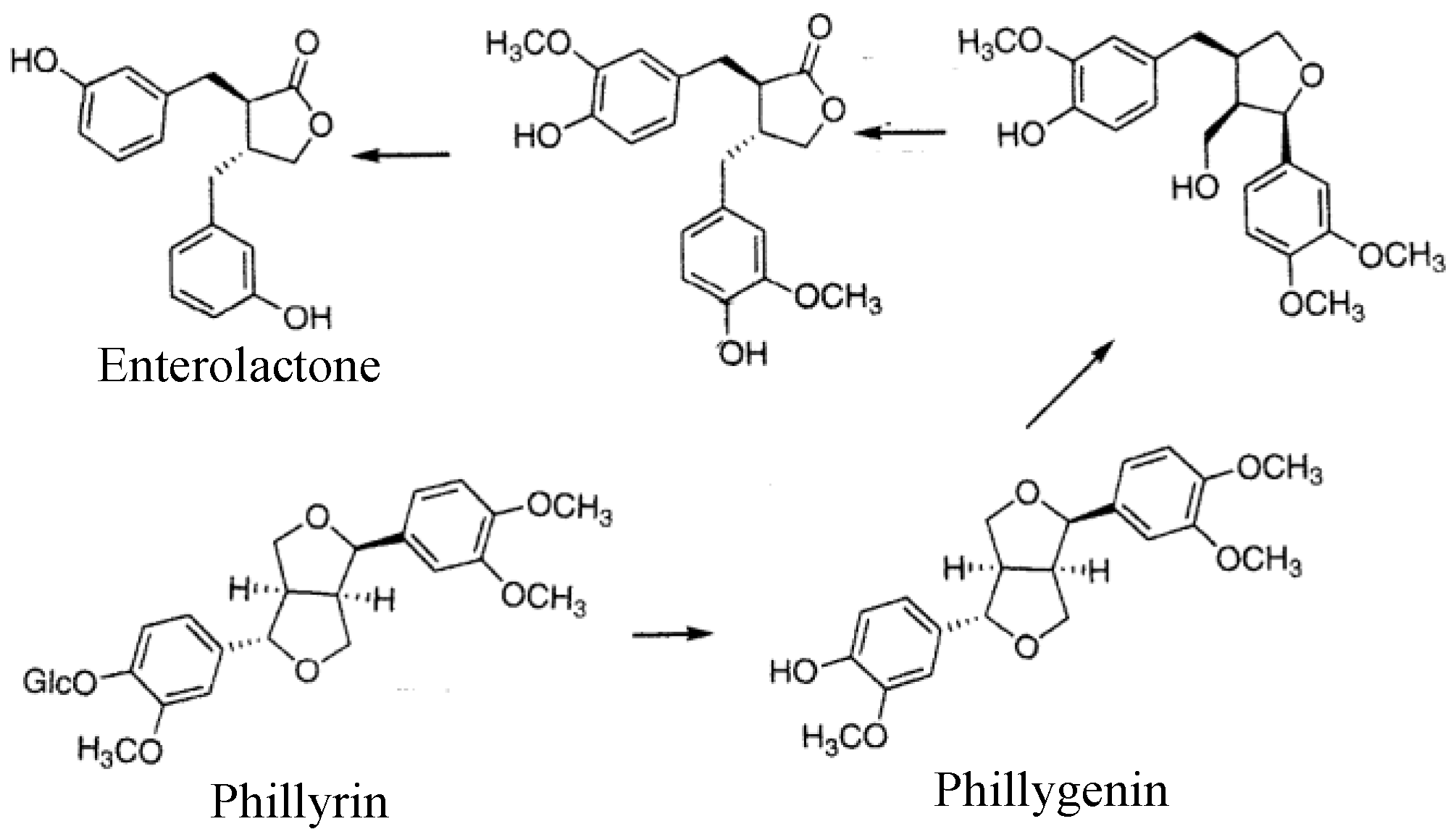
5.3. Function of Phillyrin as a Phytoestrogen
6. Toxicity and Safety of Forsythia Leaves
7. Limitations of the Study
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Urek, R.; Crncevic-Urek, M.; Cubrilo-Turek, M. Obesity—A global public health problem. Acta Med. Croat. Cas. Hravatske Akad. Med. Znan. 2007, 61, 161–1642. [Google Scholar]
- The Japanese Pharmacopoeia, 17th ed.; Minister of Health, Labour and Welfare: Tokyo, Japan, 2016; p. 1934. (In Japanese)
- Yoshida, T.; Sakane, N.; Wakabayashi, Y.; Umekawa, T.; Kondo, M. Thermogenic, anti-obesity effects of bofu-tsusho-san in MSG-obese mice. Int. J. Obes. Relat. Metab. Disord. J. Int. Assoc. Study Obes. 1995, 19, 717–722. [Google Scholar]
- Wang, Z.; Xia, Q.; Liu, X.; Liu, W.; Huang, W.; Mei, X.; Luo, J.; Shan, M.; Lin, R.; Zou, D.; et al. Phytochemistry, pharmacology, quality control and future research of Forsythia suspensa (Thunb.) Vahl: A review. J. Ethnopharmacol. 2018, 210, 318–339. [Google Scholar] [CrossRef]
- Noro, Y.; Hisata, Y.; Okuda, K.; Kawamura, T.; Tanaka, T.; Nishibe, S. Phenylethanoid glycosides in the leaves of Forsythia spp. Shoyakugaku Zasshi 1992, 46, 254–256. (In Japanese) [Google Scholar]
- Nishibe, S.; Kawamura, T.; Tanaka, T.; Aldercreutz, H. On the contents of lignans in Forsythia Fruit and in Forsythia leaf. Nat. Med. 2001, 55, 300–303. (In Japanese) [Google Scholar]
- Murase, T.; Nagasawa, A.; Suzuki, J.; Hase, T.; Tokimitsu, I. Beneficial effects of tea catechins on diet-induced obesity: Stimulation of lipid catabolism in the liver. Int. J. Obes. Relat. Metab. Disord. 2002, 26, 1459–1464. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yamasaki, M.; Katsube, T.; Sun, X.; Yamasaki, Y.; Shiwaku, K. Antiobesity effect of polyphenolic compounds from molokheiya (Corchorus olitorius L.) leaves in LDL receptor-deficient mice. Eur. J. Nutr. 2011, 50, 127–133. [Google Scholar] [CrossRef]
- Nikaido, T.; Ohmoto, T.; Kinoshita, T.; Sankawa, U.; Nishibe, S.; Hisada, S. Inhibition of cyclic AMP phosphodiesterase by lignans. Chem. Pharm. Bull. 1981, 29, 3586–3592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coon, T.A.; Mckelvey, A.C.; Weathington, N.M.; Birru, R.L.; Lear, T.; Leikauf, G.D.; Chen, B.B. Novel PDE4 inhibitors derived from Chinese medicine Forsythia. PLoS ONE 2014, 9, e115937. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Rajagopalan, S. Phosphodiesterase-4 inhibition as a therapeutic strategy for metabolic disorders. Obes. Rev. 2016, 17, 429–441. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Yoon, T.; Jang, S.; Kim, H.K. Forsythia suspensa suppresses house dust mite extract-induced atopic dermatitis in NC/Nga mice. PLoS ONE 2016, 11, e0167687. [Google Scholar]
- Qu, X.Y.; Li, Q.J.; Zhang, H.M.; Zang, X.J.; Shi, P.H.; Zhang, X.J.; Yang, J.; Zhou, Z.; Wang, S.Q. Protective effects of phillyrin against influenza A virus in vivo. Arch. Pharm. Res. 2016, 39, 998–1005. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Pang, P.; Zheng, K.; Nie, J.; Xu, H.; Wu, S.; Chen, J.; Chen, X. Forsythoside A controls influenza A virus infection and improves the prognosis by inhibiting virus replication in mice. Molecules 2016, 21, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitagawa, S.; Hisada, S.; Nishibe, S. Phenolic compounds from Forsythia leaves. Phytochemistry 1984, 23, 1635–1636. [Google Scholar] [CrossRef]
- Chiba, M.; Hisada, S.; Nishibe, S. The position of glucose linkage in phillyrin. Chem. Pharm. Bull. 1977, 25, 3435–3436. [Google Scholar] [CrossRef] [Green Version]
- Pelter, A.; Ward, R.S.; Rao, V.; Sastry, K.V. Revised structures for pluviatilol, methyl pluviatilol and xanthoxylol: General methods for the assignment of stereochemistry to 2,6-diaryl- 3,7-dioxabicyclo[3.3.0]octane lignans. Tetrahedron 1976, 32, 2783–2788. [Google Scholar] [CrossRef]
- Nishibe, S.; Okabe, K.; Tsukamoto, H.; Sakushima, A.; Hisada, S. The structure of Forsythiaside isolated from Forsythia suspensa. Chem. Pharm. Bull. 1982, 30, 1048–1050. [Google Scholar] [CrossRef] [Green Version]
- Nishibe, S.; Yamaguchi, S.; Hasegawa, M.; Oba, K.; Fujikawa, T. Anti-obesity effect of Forsythia leaf extract containing polyphenolic compounds and its mechanism. J. Trad. Med. 2012, 29, 149–155. [Google Scholar]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar]
- Ricquier, D.; Thibault, J.; Bouillaud, F.; Kuster, Y. Molecular approach to thermogenesis in brown dipose tissue. J. Biol. Chem. 1983, 258, 6675–6677. [Google Scholar] [CrossRef]
- Musovic, S.; Olofsson, C.S. Adrenergic stimulation of adiponectin secretion in visceral mouse adipocytes is blunted in high-fat diet induced obesity. Sci. Rep. 2019, 9, 10680. [Google Scholar] [CrossRef] [Green Version]
- Han, L.K.; Sumiyoshi, M.; Zhang, J.; Liu, M.X.; Zhang, X.F.; Zheng, Y.N.; Okuda, H.; Kimura, Y. Anti-obesity action of Salix matsudana leaves (Part 1). Anti-obesity action by polyphenols of Salix matsudana in high fat-diet trated rodent animals. Phytother. Res. 2003, 17, 1188–1194. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, F.Y.; An, X.; Zhou, M. Effect of phillyrin on the anti-obesiy in nutritive obesity mice. Zhong Yao Cai. 2005, 28, 123–124. (In Chinese) [Google Scholar]
- Kraynik, S.M.; Miyaoka, R.S.; Beavo, J.A. PDE3 and PDE4 isozyme-selective inhibitors are both required for synergistic activation of brown adipose tissue. Mol. Pharmacol. 2013, 83, 1155–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishibe, S.; Yamaguchi, S.; Hasegawa, M.; Oba, K.; Fujikawa, T. Anti-obesity effect of the mixture of herbal extracts containing a new material, Forsythia leaf extract. J. Trad. Med. 2013, 30, 82–90. [Google Scholar]
- Kusano, A.; Nikaido, T.; Kuge, T.; Ohmoto, T.; Monache, G.D.; Botta, B.; Botta, M.; Saitoh, T. Inhibition of adenosine 3′,5′-cyclic monophosphate phosphodiesterase by flavonoids from Licorice roots and 4-arylcoumarins. Chem. Pharm. Bull. 1991, 39, 930–933. [Google Scholar] [CrossRef] [Green Version]
- Kubo, K.; Kiyose, C.; Ogino, S.; Saito, M. Suppressive effect of Citrus aurantium against body fat accumulation and its safety. J. Clin. Biochem. Nutr. 2005, 36, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Kojima, K.; Shimada, T.; Nagareda, Y.; Watanabe, M.; Ishizaki, J.; Sai, Y.; Miyamoto, K.; Aburada, M. Preventive effect of geniposide on metabolic disease status in spontaneously obese type 2 diabetic mice and free fatty acid-treated HepG2 cells. Biol. Pharm. Bull. 2011, 34, 1613–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, A.; Sakata, M.; Morimoto, Y.; Tajima, S. Effects of Bofu-tsusho-san (Fang-Feng-Tong- Sheng-San) on body weight and fat content in normal mice. J. Trad. Med. 2001, 18, 33–38. (In Japanese) [Google Scholar]
- Akagiri, S.; Naito, Y.; Ichikawa, H.; Mizushima, K.; Takagi, T.; Handa, O.; Kokura, S.; Yoshikawa, T. Bofutsushosan, an oriental herbal medicine, attenuates the weight gain of white adipose tissue and the increased size of adipocytes associated with the increase in their expression of uncoupling protein 1 in high-fat diet-fed male KK/Ta mice. J. Clin. Biochem. Nutr. 2008, 42, 158–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, T.; Suzuki, S.; Kudo, H.; Sassa, S.; Nomura, M.; Sakamoto, S. Effects of three Chinese herbal medicines on plasma and liver lipids in mice fed a high-fat diet. J. Ethnopharmacol. 2007, 109, 236–240. [Google Scholar] [CrossRef]
- Hioki, C.; Yoshimoto, K.; Yoshida, T. Efficacy of bofu-tsusho-san, an oriental herbal medicine, in obese Japanese women with impaired glucose tolerance. Clin. Exp. Pharmacol. Physiol. 2004, 31, 614–619. [Google Scholar] [CrossRef]
- Hu, E.; Liang, P.; Spiegelman, B.M. AdipoQ is a novel adipose-specific gene dysregulated in obesity. J. Biol. Chem. 1996, 271, 10697–10703. [Google Scholar] [CrossRef] [Green Version]
- Fujikawa, T.; Hirata, T.; Wada, A.; Kawamura, N.; Yamaguchi, Y.; Fujimura, K.; Ueda, T.; Yurugi, Y.; Soya, H.; Nishibe, S. Chronic administration of eucommia leaf stimulates metabolic function of rats across several organs. Br. J. Nutr. 2010, 104, 1868–1877. [Google Scholar] [CrossRef] [Green Version]
- Matsuzawa, Y.; Funahashi, T.; Kihara, S.; Shimomura, I. Adiponectin and metabolic syndrome. Arter. Thromb. Vasc. Biol. 2004, 24, 29–33. [Google Scholar] [CrossRef] [Green Version]
- Lowell, B.B.; Susulic, V.S.; Hamann, A.; Lawitts, J.A.; Hagen, J.H.; Boyer, B.B.; Kozak, L.P.; Flier, J.S. Development of obesity in transgenic mice after genetic ablation of brown adipose tissue. Nature 1993, 366, 740–742. [Google Scholar] [CrossRef]
- Hirata, T.; Ikeda, T.; Fujikawa, T.; Nishibe, S. The chemistry and boactivity of Eucommia ulmoides Oliver leaves. In Studies in Natural Products Chemistry, 1st ed.; Atta-ur-Rahman, F.R.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 41, pp. 235–240, 256–257. [Google Scholar]
- Hosoo, S.; Koyama, M.; Kato, M.; Hirata, T.; Yamaguchi, Y.; Yamasaki, H.; Wada, A.; Wada, K.; Nishibe, S.; Nakamura, K. The restorative effects of Eucommia ulmoides Oliver leaf extract on vascular function in spontaneously hypertensive rats. Molecules 2015, 20, 21971–21981. [Google Scholar] [CrossRef] [Green Version]
- Fujikawa, T.; Hirata, T.; Hosoo, S.; Nakajima, K.; Wada, A.; Yurugi, Y.; Soya, H.; Matsui, T.; Yamaguchi, A.; Ogata, M.; et al. Asperuloside stimulates metabolic function in rats across several organs under high-fat diet conditions, acting like the major ingredient of Eucommia leaves with anti-obesity activity. J. Nutr. Sci. 2012, 1, e10. [Google Scholar] [CrossRef] [Green Version]
- Yoshinaga, N.; Mori, N. Diverse insect adaptation strategies for the defensive substance iridoid individual strategy and evolutionary direction. Kagaku Seibutsu 2018, 56, 454–456. (In Japanese) [Google Scholar] [CrossRef]
- Trim, A.R.; Hill, R. The preparation and properties of aucubin, asperuloside and some related glycosides. Biochem. J. 1952, 50, 310–319. [Google Scholar] [CrossRef] [Green Version]
- McCreight, L.J.; Bailey, C.J.; Pearson, E.R. Metformin and the gastrointestinal tract. Diabetologia 2016, 59, 426–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Horai, Y.; Houten, S.M.; Morimoto, K.; Sugizaki, T.; Arita, E.; Mataki, C.; Sato, H.; Tanigawara, Y.; Schoonjans, K.; et al. Lowering bile acid pool size with a synthetic farnesoid X receptor (FXR) agonist induces obesity and diabetes through reduced energy expenditure. J. Biol. Chem. 2011, 286, 26913–26920. [Google Scholar] [CrossRef] [Green Version]
- Kajiyama, G. The effects of bile acid on serum cholesterol. J. Jpn. Atheroscler. Soc. 1998, 26, 47–51. (In Japanese) [Google Scholar] [CrossRef] [Green Version]
- Zietak, M.; Kozak, L.P. Bile acids induce uncoupling protein 1-dependent thermogenesis and stimulate energy expenditure at thermoneutrality in mice. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E346–E354. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, A.; Yokoyama, Y.; Tanaka, K.; Benegiamo, G.; Hirayama, A.; Zhu, Q.; Kitamura, N.; Sugizaki, T.; Morimoto, K.; Itoh, H.; et al. Asperuloside improves obesity and type 2 diabetes through modulation of gut microbiota and metabolic signaling. iScience 2020, 23, 101522. [Google Scholar] [CrossRef]
- Fujisaka, S.; Usui, I.; Nawaz, A.; Igarashi, Y.; Okabe, K.; Furusawa, Y.; Watanabe, S.; Yamamoto, S.; Sasahara, M.; Watanabe, Y.; et al. Bofutsushosan improves gut barrier function with a bloom of Akkermansia muciniphila and improves glucose metabolism in mice with diet-induced obesity. Sci. Rep. 2020, 10, 5544. [Google Scholar] [CrossRef]
- Cani, P.; Vos, W.M. Next-generation beneficial microbes: The case of Akkermansia muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef]
- Imai, K.; Kobayashi, Y.; Hirata, T.; Yoshikawa, S.; Ueda, T.; Nishibe, S. Effects of a food containing Eucommia leaf extract on body fat reduction. Jpn. Pharmacol. Ther. 2017, 45, 93–102. (In Japanese) [Google Scholar]
- Brufau, G.; Bahr, M.J.; Staels, B.; Claudel, T.; Ockenga, J.; Boker, K.H.; Murphy, E.J.; Prado, K.; Stellaard, F.; Manns, M.P.; et al. Plasma bile acids are not associated with energy metabolism in humans. Nutr. Metab. 2010, 7, 73. [Google Scholar] [CrossRef] [Green Version]
- Kawada, T. Food function study of obesity and fat energy metabolism. Nippon Eiyo Shokuryo Gakkaishi 2014, 67, 119–125. (In Japanese) [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Ahmad, F.; Philp, A.; Baar, K.; Williams, T.; Luo, H.; Ke, H.; Rehmann, H.; Taussig, R.; Brown, A.L.; et al. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell 2012, 148, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Glatz, M.; Horiuchi, K.; Kawasaki, H.; Akiyama, H.; Kaplan, D.H.; Kong, H.H.; Amagai, M.; Nagao, K. Dysbiosis and Staphylococcus aureus colonization drives inflammation in atopic dermatitis. Immunity 2015, 42, 756–766. [Google Scholar] [CrossRef] [Green Version]
- Nishibe, S.; Okabe, K.; Tsukamoto, H.; Sakushima, A.; Hisada, S.; Baba, H.; Akisada, T. Studies on the Chinese crude drug “Forsythiae Fructus” VI. The structure and antibacterial activity of suspensaside isolated from Forsythia suspensa. Chem. Pharm. Bull. 1982, 30, 4548–4553. [Google Scholar] [CrossRef] [Green Version]
- Kitagawa, S.; Nishibe, S.; Baba, H. Studies on the Chinese crude drug “Forsythiae Fructus”. VIII. On isolation of phenylpropanoid glycosides from fruits of Forsythia koreana and their antibacterial activity. Yakugaku Zasshi. 1987, 107, 274–278. (In Japanese) [Google Scholar] [CrossRef]
- Qu, H.; Zhang, Y.; Wang, Y.; Li, B.; Sun, W. Antioxidant and antibacterial activity of two compounds (forsythiaside and forsythin) isolated from Forsythia suspensa. J. Pharm. Pharmacol. 2008, 60, 261–266. [Google Scholar] [CrossRef]
- Ahluwalia, J.; Udkoff, J.; Waldman, A.; Borok, J.; Eichenfield, L.F. Phosphodiesterase 4 inhibitor therapies for atopic dermatitis: Progress and outlook. Drugs 2017, 77, 1389–1397. [Google Scholar] [CrossRef]
- Hanifin, J.M.; Chan, S.S.; Cheng, J.B.; Tofte, S.J.; Henderson, W.R., Jr.; Kirby, D.S.; Weiner, E.S. Type 4 phosphodiesterase inhibitors have clinical and in vitro anti-inflammatory effects in atopic dermatitis. J. Investig. Dermatol. 1996, 107, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Grewe, S.R.; Chan, S.C.; Hanifin, J.M. Elevated leukocyte cyclic AMP-phosphodiesterase in atopic disease: A possible mechanism for cyclic AMP-agonist hyporesponsiveness. J. Allergy Clin. Immunol. 1982, 70, 452–457. [Google Scholar] [CrossRef]
- Butler, J.M.; Chan, S.C.; Stevens, S.; Hanifin, J.M. Increased leukocyte histamine release with elevated cyclic AMP-phosphodiesterase activity in atopic dermatitis. J. Allergy Clin. Immunol. 1983, 71, 490–497. [Google Scholar] [CrossRef]
- Heskel, N.S.; Chan, S.C.; Thiel, M.L.; Stevens, S.R.; Casperson, L.S.; Hanifin, J.M. Elevated umbilical cord blood leukocyte cyclic adenosine monophophate-phosphodiesterase activity in children with atopic parents. J. Am. Acad. Dermatol. 1984, 11, 422–426. [Google Scholar] [CrossRef]
- Nishibe, S. Bioactive phenolic compounds for cancer prevention from herbal medicines. In Food Factors for Cancer Prevention; Ohigashi, H., Osawa, T., Terao, J., Watanabe, S., Yoshikawa, T., Eds.; Springer: Tokyo, Japan, 1997; pp. 276–279. [Google Scholar]
- Heinonen, S.; Nurmi, T.; Liukkonen, K.; Poutanen, K.; Wahala, K.; Deyama, T.; Nishibe, S.; Adlercreutz, H. In vitro metabolism of plant lignans: New precursors of mammalian lignans enterolactone and enterodiol. J. Agric. Food Chem. 2001, 49, 3178–3186. [Google Scholar] [CrossRef]
- Nishibe, S. On the utilization of Forsythia Leaf. Medical Herb. 2015, 34, 30–34. (In Japanese) [Google Scholar]
- Nishibe, S.; Kawamura, T.; Fujikawa, T. The toxicity test of Forsythia leaf extract by single and repeated oral administration to mice. In Proceedings of the 135th Annual Meeting of the Pharmaceutical Society of Japan, Kobe, Japan, 25–27 March 2015. 27PB-am198 (In Japanese). [Google Scholar]
- Ai, X.; Dong, X.T.; Wang, I.J.; Han, X.; Luo, C.R. Acute toxicity test and micronucleus test of Forsythia suspena leaves in mice. J. Anhui Agri. Sci. 2011, 11, 6397–6398. [Google Scholar]
- Wu, J.F.; Zhang, Z.W.; Kuang, L.; Shen, H.; Lu, Y.; Wu, G.J. Study on the toxicity of forsythiaside on cell culture in vitro. Chin. J. Vet. Drug. 2009, 6, 4–8. [Google Scholar]
- Mao, D.Y.; Zhang, Z.W.; Yang, M.; Wang, H.C.; Wu, G.J. Acte toxicity and the induction on IFN-á of forthythiaside in mice. Prog. Vet. Med. 2009, 6, 15–17. [Google Scholar]
- Han, Z.; Lei, X.L.; Zhang, H.; Liu, L.; Chen, Z.S.; Yang, W.; Lun, Z.R. Evaluating the safety of forsythin from Forsythia suspensa leaves by acute and sub-chronic oral administration in rodent models. Asian Pac. J. Trop. Med. 2017, 10, 47–51. [Google Scholar] [CrossRef]
- Dong, Z.; Lu, X.; Tong, X.; Dong, Y.; Tang, L.; Liu, M. Forsythiae Fructus: A review on its phytochemistry, quality control, pharmacology and pharmacokinetics. Molecules 2017, 22, 1466. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C-1′ | C-1″ | |
|---|---|---|
| Phillyrin | 131.0 | 135.4 |
| Phillygenin methyl ether | 131.0 | 133.8 |
| Phillygenin | 131.0 | 133.1 |
| Phillygenin acetate | 131.0 | 140.3 |
| Diet | |||
|---|---|---|---|
| Control (n = 9) | 2.5% FLE (n = 8) | 5.0% FLE (n = 8) | |
| Food intake (g/rat/day) | |||
| The first week | 13.3 ± 0.25 | 12.6 ± 0.61 | 12.9 ± 0.26 |
| The fourth week | 15.6 ± 0.71 | 14.9 ± 0.88 | 14.4 ± 073 |
| Body weight and organ weight (g/rat) | |||
| Initial body weight | 74.7 ± 0.58 | 75.3 ± 1.40 | 74.8 ± 0.91 |
| Final body weight | 255.1 ± 7.41 | 167.8 ± 1.77 * | 148.2 ± 3.74 * |
| Perirenal white adipose tissue | 1.44 ± 0.22 | 0.47 ± 0.10 * | 0.33 ± 0.07 * |
| Epididymal white adipose tissue | 3.14 ± 0.21 | 1.22 ± 0.07 * | 1.02 ± 0.09 * |
| Brown adipose tissue | 1.02 ± 0.04 | 0.61 ± 0.02 * | 0.59 ± 0.03 * |
| Liver | 2.05 ± 0.05 | 1.55 ± 0.09 * | 1.51 ± 0.06 * |
| Plasma parameters | |||
| Triglyceride (mg/dL) | 124.4 ± 15.1 | 65.5 ± 11.4 * | 47.4 ± 4.7 * |
| Free fatty acid (μEq/L) | 474.3 ± 29.5 | 318.8 ± 35.4 * | 280 ± 18.6 * |
| HDL-cholesterol (mg/dL) | 38.0 ± 1.3 | 46.1 ± 2.4 | 47.8 ± 1.6 |
| LDL-cholesterol (mg/dL) | 14.0 ± 0.8 | 17.0 ± 0.9 | 15.4 ± 0.7 |
| Total cholesterol (mg/dL) | 102.6 ± 3.7 | 121.9 ± 5.0 | 115 ± 3.9 |
| Glucose (mg/dL) | 168.4 ± 4.3 | 174.8 ± 16.6 | 168.1 ± 4.9 |
| Fold Change to Control | ||
|---|---|---|
| Gene Name (Accession No) | 2.5% FLE | 5.0% FLE |
| Glycolitic system | ||
| Gck (NM012565) | 1.66 ± 0.14 * | 1.53 ± 0.05 |
| TCA cycle | ||
| Cs (NM130755) | 1.03 ± 0.07 | 1.21 ± 0.10 |
| Ogdh (AI412142) | 0.89 ± 0.05 | 0.96 ± 0.04 |
| Electron transfer system | ||
| Comp. I (CB5449004) | 0.99 ± 0.02 | 1.03 ± 0.07 |
| Comp. IV (NM017202) | 1.84 ± 0.07 * | 1.98 ± 0.05 * |
| Fatty acid synthesis | ||
| Fasn (NM017332) | 1.19 ± 0.13 | 1.24 ± 0.08 |
| Fatty acid transporter | ||
| Fatp (NM053580) | 1.91 ± 0.11 * | 1.92 ± 0.04 * |
| Fatty acid β-oxidation | ||
| Cpt1o (NM031559) | 1.84 ± 0.03 * | 1.81 ± 0.15 * |
| Acadvl (NM012891) | 1.94 ± 0.67 * | 1.91 ± 0.10 * |
| Fold Change to Control | ||
|---|---|---|
| Gene Name (Accession No) | 2.5% FLE | 5.0% FLE |
| Perirenal white adipose tissue | ||
| Fatty acid receptor and adipocytokine | ||
| PPARγ (NM013124) | 2.74 ± 0.04 ** | 2.85 ± 0.13 ** |
| Adiponectin (NM144744) | 1.93 ± 0.13 * | 1.97 ± 0.03 * |
| Resistin (AJ555618) | 1.56 ± 0.10 | 1.33 ± 0.13 |
| Brown adipose tissue | ||
| Uncopling ATP synthesis from oxidative metabolism | ||
| UCP1 (NM012682) | 1.88 ± 0.02 * | 1.92 ± 0.11 * |
| Diet (ND) | ||||
|---|---|---|---|---|
| Control (n = 4) | 0.2% MHE (n = 4) | 1% MHE (n = 4) | 5% MHE (n = 4) | |
| Food intake (g/rat) | ||||
| Final day | 43.2 ± 1.2 | 44.1 ± 0.3 | 40.2 ± 0.8 | 44.2 ± 0.1 |
| Body weight (g/rat) | ||||
| Final body weight | 491 ± 13 | 445 ± 14 | 454 ± 16 | 416 ± 15 |
| Organ weight/(g/100 g body weight) | ||||
| Perirenal white adipose tissue | 0.67 ± 0.16 | 0.36 ± 0.06 | 0.36 ± 0.06 | 0.31 ± 0.05 * |
| Epididymal white adipose tissue | 1.71 ± 0.21 | 1.14 ± 0.14 * | 1.29 ± 0.13 | 1.03 ± 0.06 * |
| Brown adipose tissue | 0.13 ± 0.01 | 0.15 ± 0.01 | 0.18 ± 0.01 | 0.19 ± 0.02 |
| Plasma parameters | ||||
| Triglyceride (mg/dL) | 232 ± 38 | 128 ± 19 * | 89.0 ± 3.8 * | 81.5 ± 4.0 * |
| Free fatty acid (μEq/L) | 355 ± 46 | 362 ± 30 | 262 ± 22 | 313 ± 68 |
| HDL-cholesterol (mg/dL) | 32.0 ± 1.7 | 27.3 ± 1.1 | 25.8 ± 1.2 * | 31.5 ± 2.2 |
| LDL-cholesterol (mg/dL) | 11.0 ± 0.7 | 8.50 ± 0.29 | 8.50 ± 0.87 | 8.25 ± 0.25 |
| Total cholesterol (mg/dL) | 93.5 ± 7.7 | 72.0 ± 2.3 | 68.0 ± 4.2 * | 81.5 ± 5.8 |
| Glucose (mg/dL) | 150 ± 7 | 151 ± 3 | 138 ± 5 | 154 ± 4 |
| Insulin (ng/mL) | 1.27 ± 0.31 | 1.49 ± 0.06 | 1.49 ± 0.23 | 1.90 ± 0.15 |
| Adiponectin (ng/mL) | 3881 ± 461 | 4749 ± 1111 | 3223 ± 360 | 3717 ± 544 |
| Diet (HFD) | ||||
|---|---|---|---|---|
| Control (n = 6) | 0.2% MHE (n = 6) | 1% MHE (n = 6) | 5% MHE (n = 6) | |
| Food intake (g/rat) | ||||
| Final day | 35.5 ± 0.4 | 31.8 ± 0.8 | 32.9 ± 0.5 | 39.3 ± 3.7 |
| Body weight (g/rat) | ||||
| Final body weight | 588 ± 6 † | 561 ± 23 | 539 ± 13 | 410 ± 5 * |
| Organ weight (g/100g body weight) | ||||
| Perirenal white adipose tissue | 1.64 ± 0.06 † | 1.42 ± 0.11 | 1.40 ± 0.15 | 0.74 ± 0.11 ** |
| Epididymal white adipose tissue | 3.41 ± 0.07 † | 3.03 ± 0.22 | 2.53 ± 0.10 * | 1.53 ± 0.17 ** |
| Brown adipose tissue | 0.20 ± 0.01 † | 0.21 ± 0.02 | 0.28 ± 0.02 | 0.24 ± 0.02 |
| Plasma parameters | ||||
| Triglyceride (mg/dL) | 229 ± 6 | 164 ± 36 | 207 ± 47 | 65.3 ± 16.4 ** |
| Free fatty acid (μEq/L) | 530 ± 25 † | 387 ± 5 * | 396 ± 26 * | 454 ± 51 |
| HDL-cholesterol (mg/dL) | 32.2 ± 2.5 | 28.7 ± 1.2 | 34.0 ± 2.1 | 40.7 ± 1.3 * |
| LDL-cholesterol (mg/dL) | 15.7 ± 1.4 † | 11.3 ± 1.0 * | 14.3 ± 1.2 | 13.7 ± 1.7 |
| Total cholesterol (mg/dL) | 125 ± 7 † | 83.0 ± 2.5 * | 101 ± 10 * | 110 ± 7 |
| Glucose (mg/dL) | 177 ± 5 † | 160 ± 6 * | 162 ± 3 * | 153 ± 3 * |
| Insulin (ng/mL) | 2.62 ± 0.07 † | 1.00 ± 0.13 ** | 1.49 ± 0.30 * | 1.31 ± 0.28 * |
| Adiponectin (ng/mL) | 3332 ± 200 | 6342 ± 998 * | 4079 ± 237 | 5194 ± 757 * |
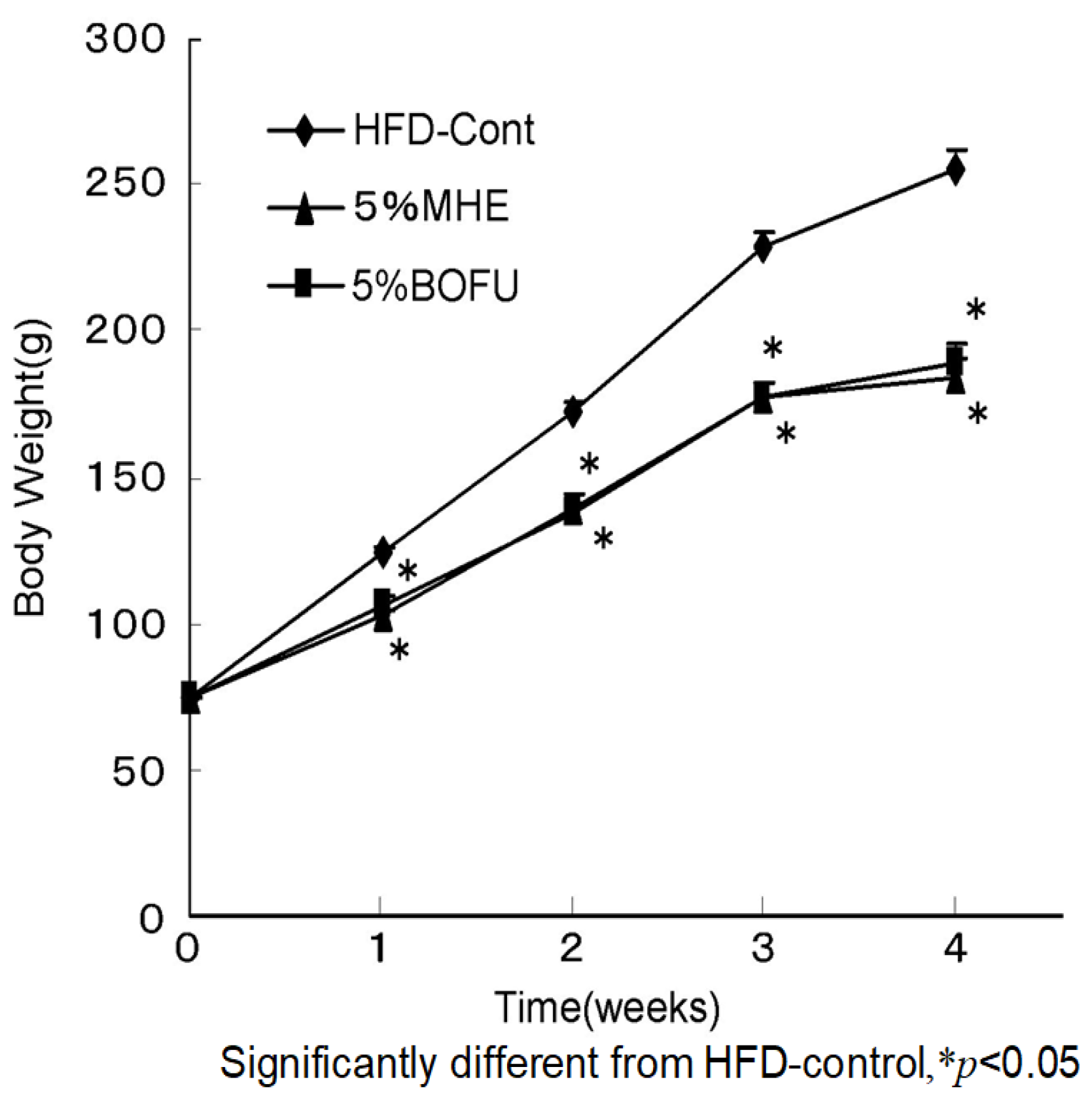
| Diet (HFD) | |||
|---|---|---|---|
| Control (n = 9) | 5% MHE (n = 8) | 5% BOFU (n = 8) | |
| Body weight (g/rat) | |||
| Initial body weight | 74.7 ± 0.6 | 74.8 ± 0.9 | 74.2 ± 0.9 |
| Final body weight | 255.1 ± 7.4 | 183.3 ± 7.8 * | 189.6 ± 5.9 * |
| Organ weight (g/100 g body weight) | |||
| Perirenal white adipose tissue | 0.53 ± 0.08 | 0.31 ± 0.08 * | 0.31 ± 0.07 * |
| Epididymal white adipose tissue | 1.2 ± 0.06 | 0.63 ± 0.09 * | 0.69 ± 0.05 * |
| Brown adipose tissue | 0.38 ± 0.01 | 0.35 ± 0.02 | 0.36 ± 0.02 |
| Fold Change to Control | ||
|---|---|---|
| Gene Name (Accession No) | 5% MHE | 5% BOFU |
| Glycolytic system | ||
| Gck (NM012565) | 0.54 ± 0.05 *** | 0.45 ± 0.09 ** |
| TCA cycle | ||
| Cs (NM130755) | 1.08 ± 0.12 | 0.97 ± 0.09 |
| Ogdh (AI412142) | 1.16 ± 0.06 | 1.18 ± 0.06 |
| Electron transfer system | ||
| Comp. I (CB5449004) | 1.49 ± 0.06 | 0.89 ± 0.06 |
| Comp. IV (NM017202) | 1.89 ± 0.04 * | 1.84 ± 0.07 * |
| Fatty acid synthesis | ||
| Fasn (NM017332) | 0.46 ± 0.03 *** | 0.80 ± 0.11 |
| Fatty acid transporter | ||
| Fatp (NM053580) | 1.87 ± 0.20 * | 1.82 ± 0.17 * |
| Fatty acid β-oxidation | ||
| Cpt1α (NM031559) | 1.94 ± 0.06 * | 1.74 ± 0.06 * |
| ACADVL (NM012891) | 1.87 ± 0.11 * | 1.87 ± 0.09 * |
| Fold Change to Control | ||
|---|---|---|
| Gene Name (Accession No) | 5% MHE | 5% BOFU |
| Perirenal white adipose tissue | ||
| Fatty acid receptor and adipocytokine | ||
| PPARγ (NM013124) | 2.22 ± 0.09 * | 1.89 ± 0.25 * |
| Adiponectin (NM144744) | 2.42 ± 0.05 ** | 1.92 ± 0.19 * |
| Resistin (AJ555618) | 1.30 ± 0.07 | 1.60 ± 0.17 |
| Brown adipose tissue | ||
| Uncoupling ATP synthesis from oxidative metabolism | ||
| UCP1 (NM012682) | 1.86 ± 0.09 * | 1.32 ± 0.05 |
| Diet (HFD) | |||
|---|---|---|---|
| Control (n = 8) | 3% ELE (n = 8) | 9% ELE (n = 8) | |
| Final body weight (g/rat) | 548.6 ± 16.8 | 485.3 ± 13.6 * | 422.2 ± 17.7 * |
| Food intake (g/day/rat) | 25.3 ± 3.0 | 19.7 ± 3.6 | 15.2 ± 1.7 * |
| WAT weight (g/rat) | |||
| Perirenal white adipose tissue | 10.0 ± 0.8 | 5.6 ± 0.4 *** | 3.5 ± 0.6 *** |
| Epididymal white adipose tissue | 18.3 ± 0.7 | 13.9 ± 0.6 *** | 5.8 ± 0.4 *** |
| Plasma parameters | |||
| Glucose (mg/L) | 1520 ± 17 | 1458 ± 5 * | 1433 ± 17 * |
| Insulin (ng/mL) | 6.6 ± 0.5 | 4.2 ± 0.5 ** | 2.4 ± 0.3 *** |
| Free fatty acid (µEq/L) | 610.4 ± 78.8 | 450.8 ± 33.8 | 493.4 ± 26.2 |
| Total cholesterol (mg/L) | 780 ± 27 | 655 ± 28 ** | 725 ± 15 |
| Adiponectin (µg/L) | 27 ± 3 | 42 ± 4 | 53 ± 4 ** |
| TNF-α (pg/mL) | 178.5 ± 22.6 | 137.1 ± 15.1 | 63.5 ± 8.3 * |
| Resistin (ng/mL) | 187.6 ± 15.9 | 175.9 ± 15.9 | 111.4 ± 11.0 ** |
| Leptin (ng/mL) | 6.8 ± 0.4 | 5.9 ± 0.7 | 6.7 ± 0.8 |
| Diet (HFD) | ||||
|---|---|---|---|---|
| Control (n = 6) | 0.03% ASP (n = 6) | 0.1% ASP (n = 6) | 0.3% ASP (n = 6) | |
| Initial body weight (g/rat) | 71 ± 1.0 | 71.2 ± 1.5 | 72.5 ± 0.5 | 71 ± 0.6 |
| Food intake (g/day/rat) | 27.8 ± 2.2 | 21.3 ± 3.2 * | 17.7 ± 2.7 * | 14.9 ± 2.0 * |
| Final body weight (g/rat) | 564 ± 9 | 516 ± 19 * | 465 ± 8 * | 461 ± 7 * |
| Body weight gain (g/rat) | 493 ± 10 | 445 ± 18 * | 393 ± 8 * | 390 ± 7 * |
| Relative WAT weight (%) | ||||
| Perirenal white adipose tissue | 2.7 ± 0.3 | 1.5 ± 0.2 * | 1.4 ± 0.1 * | 1.3 ± 0.1 * |
| Epididymal white adipose tissue | 2.6 ± 0.2 | 2.5 ± 0.2 | 2.2 ± 0.1 | 2.0 ± 0.1 |
| Relative BAT weight (%) | 0.24 ± 0.02 | 0.31 ± 0.01 * | 0.33 ± 0.02 * | 0.37 ± 0.02 * |
| Relative Sol. M. weight (%) | 0.07 ± 0.01 | 0.07 ± 0.01 | 0.07 ± 0.01 | 0.07 ± 0.01 |
| Plasma parameters | ||||
| Glucose (mg/L) | 1621 ± 71 | 1501 ± 37 * | 1394 ± 42 * | 1338 ± 55 * |
| Insulin (ng/mL) | 7.7 ± 0.6 | 5.2 ± 1.1 * | 3.9 ± 0.8 * | 3.3 ± 0.6 * |
| Free fatty acid (µEq/L) | 639.1 ± 33.7 | 449 ± 56.0 * | 402.7 ± 21.6 * | 397.3 ± 20.9 * |
| Total cholesterol (mg/L) | 880 ± 34 | 721 ± 25 * | 708 ± 24 * | 664 ± 26 * |
| Adiponectin (µg/L) | 29 ± 5 | 39 ± 6 * | 48 ± 4 * | 53 ± 3 * |
| TNF-α (pg/mL) | 198.3 ± 18.2 | 136.5 ± 13.1 * | 98.7 ± 9.2 * | 70.6 ± 8.9 * |
| Non-Intake Number (%) | Intake Number (%) | |
|---|---|---|
| Infected | 15 (12.2) | 6 (7.9) |
| Non-infected | 108 (87.8) | 70 (92.1) |
| Totals | 123 | 76 |
| Days | 0 | 1 | 2 | 3 | 7 | 14 |
|---|---|---|---|---|---|---|
| Cont (5) | 32.9 ± 3.15 | 29.1 ± 3.59 | 33.0 ± 3.26 | 33.4 ± 2.80 | 35.1 ± 1.88 | 37.0 ± 1.58 |
| FLE (5) | 33.5 ± 3.35 | 29.1 ± 3.15 | 34.1 ± 2.97 | 33.7 ± 3.13 | 36.1 ± 1.89 | 37.6 ± 1.67 |
| (n) | BW (g) | Spleen (g) | Liver (g) | Renal (g) | Lung (g) | Heart (g) | Adrenal (mg) | Thymus (mg) | Testis (g) | Brain (g) | Hypophysis (mg) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cont (5) | 37.1 ± 1.00 | 0.12 ± 0.019 | 1.8 ± 0.12 | 0.63 ± 0.023 | 0.18 ± 0.012 | 0.17 ± 0.012 | 5.3 ± 0.87 | 50.1 ± 2.23 | 0.23 ± 0.013 | 0.48 ± 0.025 | 1.2 ± 0.42 |
| FLE (5) | 38.2 ± 1.31 | 0.12 ± 0.026 | 1.9 ± 0.13 | 0.59 ± 0.043 | 0.19 ± 0.018 | 0.17 ± 0.008 | 5.3 ± 0.88 | 51.9 ± 14.55 | 0.24 ± 0.027 | 0.48 ± 0.021 | 1.3 ± 0.56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishibe, S.; Mitsui-Saitoh, K.; Sakai, J.; Fujikawa, T. The Biological Effects of Forsythia Leaves Containing the Cyclic AMP Phosphodiesterase 4 Inhibitor Phillyrin. Molecules 2021, 26, 2362. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26082362
Nishibe S, Mitsui-Saitoh K, Sakai J, Fujikawa T. The Biological Effects of Forsythia Leaves Containing the Cyclic AMP Phosphodiesterase 4 Inhibitor Phillyrin. Molecules. 2021; 26(8):2362. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26082362
Chicago/Turabian StyleNishibe, Sansei, Kumiko Mitsui-Saitoh, Junichi Sakai, and Takahiko Fujikawa. 2021. "The Biological Effects of Forsythia Leaves Containing the Cyclic AMP Phosphodiesterase 4 Inhibitor Phillyrin" Molecules 26, no. 8: 2362. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26082362