
Analysis of Chemical Composition and In Vitro and In Vivo Antifungal Activity of Raphanus raphanistrum Extracts against Fusarium and Pythiaceae, Affecting Apple and Peach Seedlings

 ,
,
Abstract
:1. Introduction
2. Results
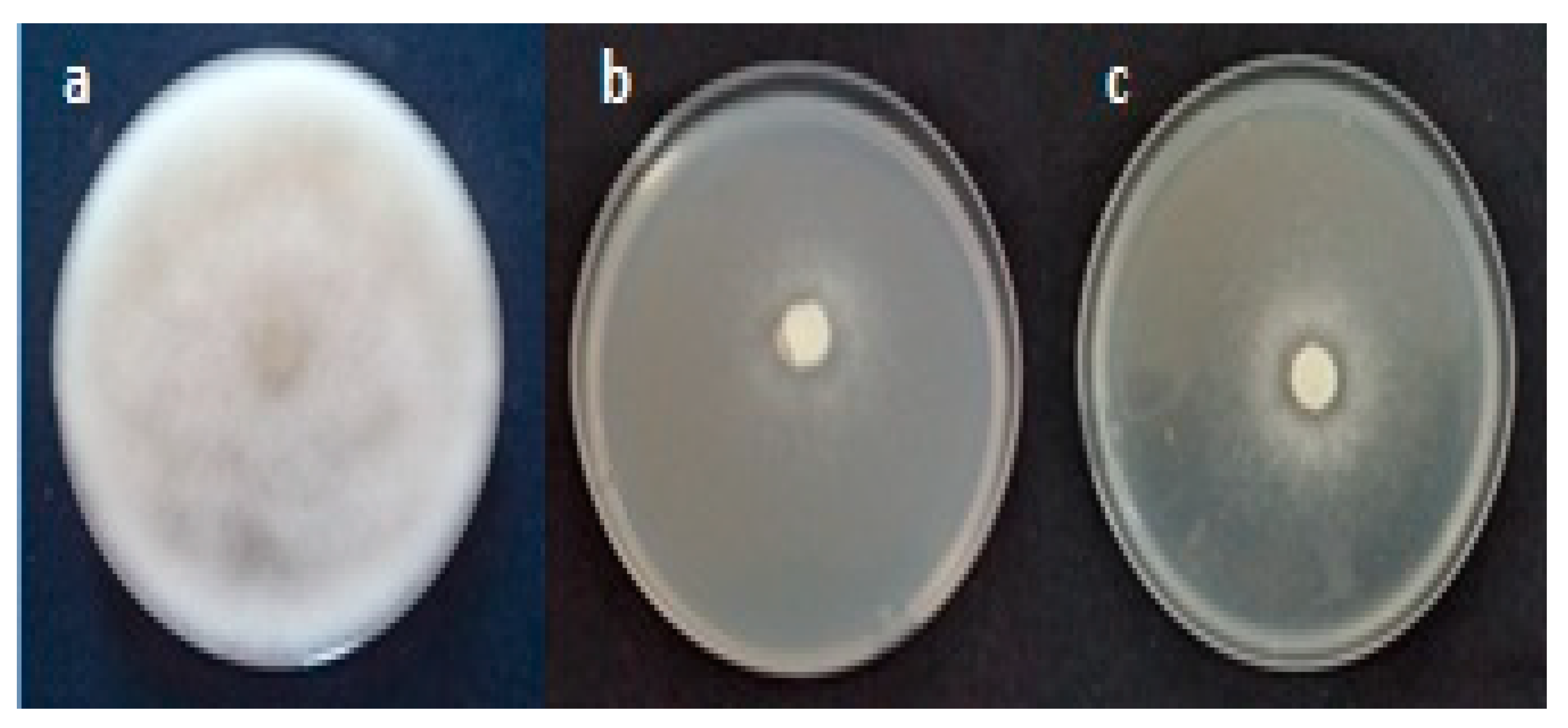
2.1. In Vitro Evaluation of Aqueous Extract Against Fusarium and Oomycete Species
2.2. Effect of Raphanus raphanistrum Plant Extracts on Peach Seedling Decline Severity
2.3. Effect of Raphanus raphanistrum Plant Extracts on Apple Seedling Decline Severity
2.4. Chemical Composition of Raphanus raphanistrum
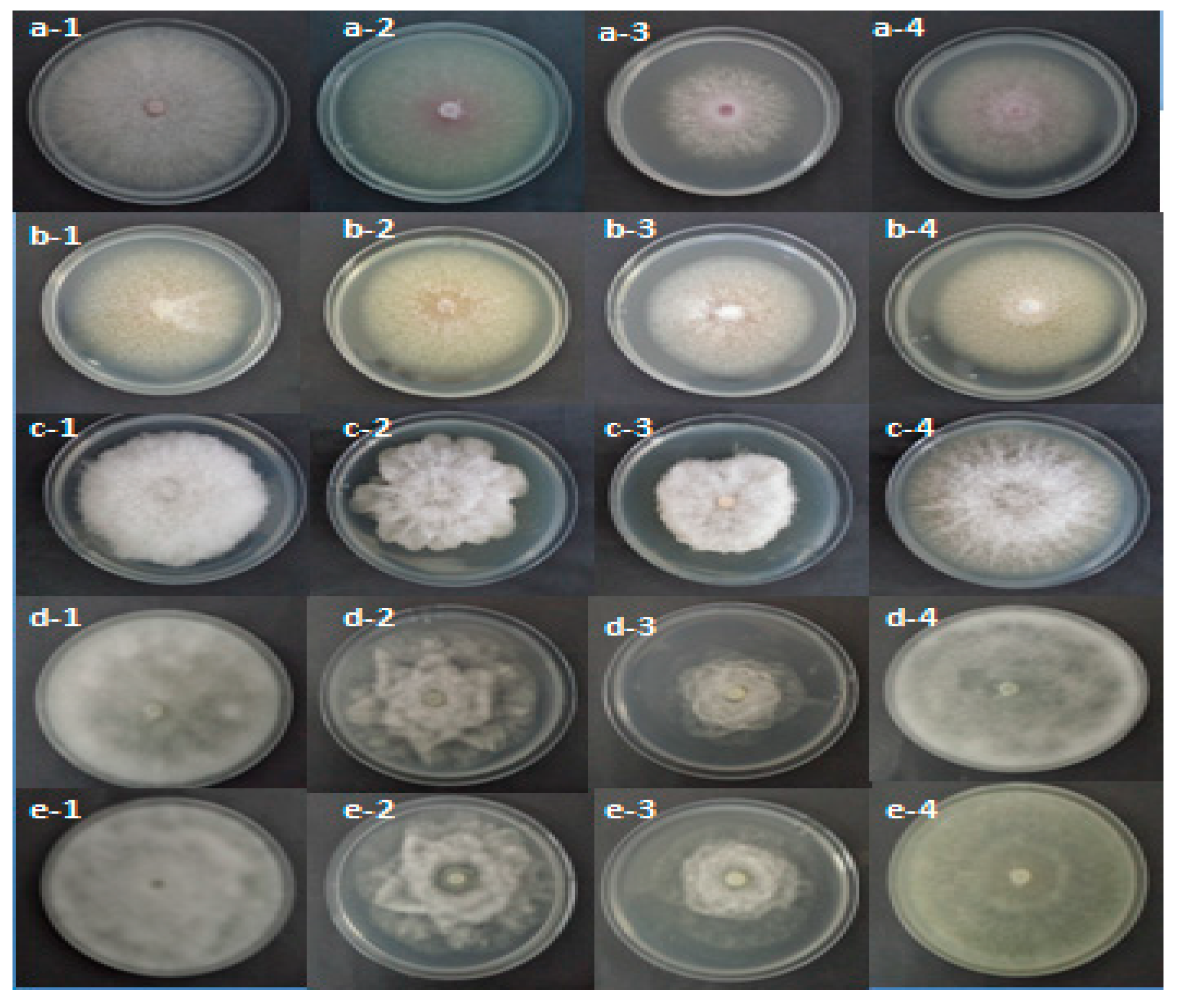
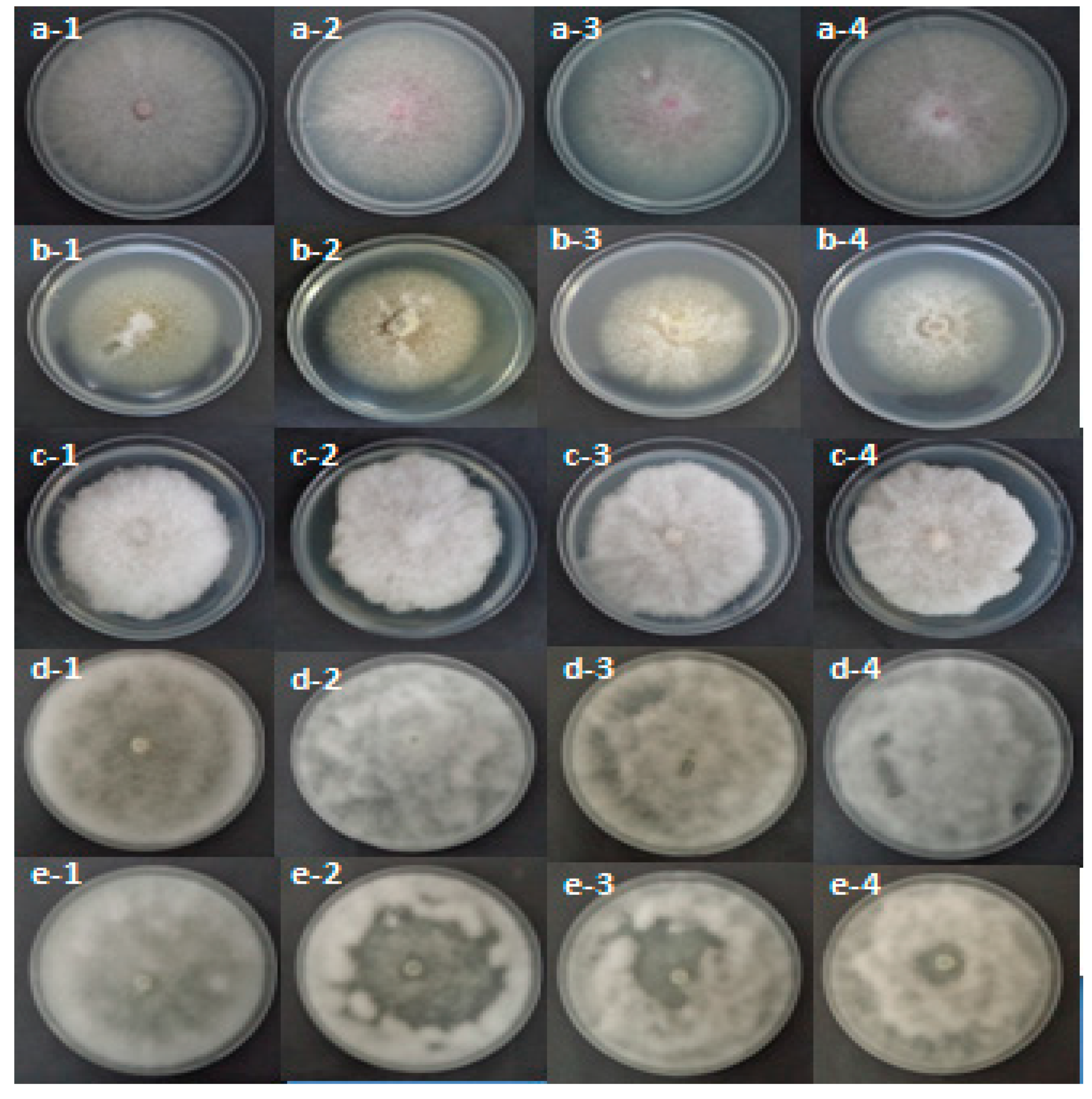
2.4.1. Effect of R. raphanistrum Organic Extracts on Mycelial Growth of Pathogens Associated with Apple and Peach Decline in Tunisian Nurseries
2.4.2. Liquid Chromatography (LC) and Thin Layer Chromatography Analysis
2.4.3. Effect of Raphanus raphanistrum Ethyl Acetate Extract Fractions on Mycelial Growth of Pathogens Associated with Apple and Peach Seedlings Decline in Nurseries
2.4.4. Effect of Raphanus raphanistrum Dichloromethane Extract Fractions on Mycelial Growth of Pathogens Associated with Apple and Peach Seedlings Decline in Nurseries
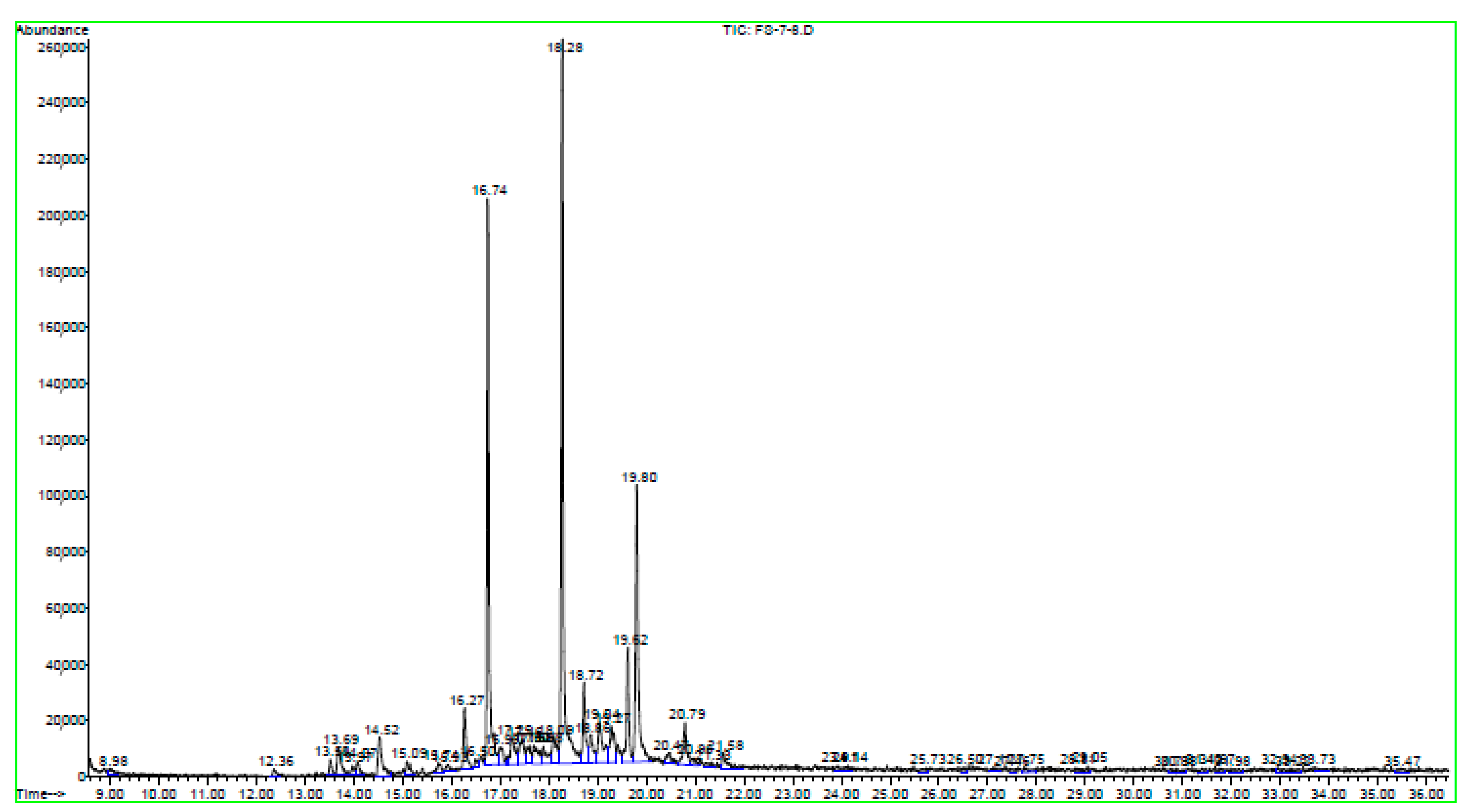
2.4.5. Gas Chromatography Analysis (GCA)
3. Discussion
4. Materials and Methods
4.1. Used Pathogens
4.2. Preparation of Raphanus raphanistrum Extracts
4.3. Effect of Raphanus raphanistrum Aqueous Extract on the Mycelial Growth of Agents Associated with Apple and Peach Seedlings Decline
4.4. Effect of Raphanus raphanistrum Powder on Apple and Peach Seedling Decline Severity
4.5. Chemical Composition of Raphanus raphanistrum
4.5.1. Preparation of Raphanus raphanistrum Organic Extracts
4.5.2. Liquid Chromatography (LC) and Thin Layer Chromatography Analysis
4.5.3. Gas Chromatography Analysis (GCA)
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Yuan, B.; Zhan, J.; Chen, C. Evolution of a Development Model for Fruit Industry against Background of an Aging Population: Intensive or Extensive Adjustment? Sustainability 2018, 10, 49. [Google Scholar] [CrossRef] [Green Version]
- Bent, E.; Loffredo, A.; Yang, J.I.; McKenry, M.V.; Becker, J.O.; Borneman, J. Investigations into peach seedling stunting caused by a replant soil. FEMS Microbiol. Ecol. 2009, 68, 192–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzola, M.; Manici, M. Apple replant disease: Role of microbial ecology in cause and control. Annu. Rev. Phytopathol. 2012, 50, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Boughalleb, N.; Moulahi, A.; El Mahjoub, M. Variability in Pathogenicity among Tunisian Isolates of Phytophthora cactorum as Measured by Their Ability to Cause Crown Rot on Four Apple Cultivars and MM106 Roostock. J Agron. 2006, 5, 321–325. [Google Scholar]
- Souli, M.; Boughalleb, N.; Abad-Campos, P.; Álvarez, L.A.; Pérez-Sierra, A.; Armengol, J.; García-Jiménez, J. First Report of Pythium indigoferae and P. irregulare Associated to Apple Trees Decline in Tunisia. J. Phytopathol. 2011, 159, 352–357. [Google Scholar] [CrossRef]
- Souli, M.; Boughalleb, N.; Abad-Campos, P.; Álvarez, L.A.; Pérez-Sierra, A.; Armengol, J.; García-Jiménez, J.; Romdhani, M.S. Diversity of the Pythium community infecting crown and roots apple in Tunisia. Res. Plant Biol. 2011, 1, 16–22. [Google Scholar]
- Mannai, S.; Benfradj, N.; Horrigue-Raouani, N.; Boughalleb-M’Hamdi, N. Prevalence of Fusarium Species Associated with Peach Decline in Tunisian Nurseries. Microbiol. Res. J. Int. 2018, 23, 1–16. [Google Scholar] [CrossRef]
- Mannai, S.; Horrigue-Raouani, N.; Boughalleb-M’Hamdi, N. Characterization of Fusarium species associated with apple decline in Tunisian nurseries. J. Biol. Stud. 2018, 1, 14–34. [Google Scholar]
- Mannai, S.; Najwa, B.; Boughalleb-M’Hamdi, N. Occurrence of Pythium and Phytopythium species associated with apple and peach seedlings decline in Tunisian nurseries. Manuscript in preparation.
- Menge, J.A. Use of new systemic fungicides on citrus. Citrograph 1986, 71, 245–250. [Google Scholar]
- Utkhede, R.S.; Smith, E.M. Phytophthora and Pythium species associated with root rot of young apple trees and their control. Soil Biol. Biochem. 1991, 23, 1059–1063. [Google Scholar] [CrossRef]
- Thomidis, T.; Elena, K. Effects of metalaxyl, fosetyl-Al, dimethomorph and cymoxanil on Phytophthora cactorum of peach tree. J. Phytopathol. 2001, 149, 97–101. [Google Scholar] [CrossRef]
- Harman, G.E.; Kubicek, C.P. Trichoderma and Gliocladium; Taylor & Francis: London, UK, 1998; p. 278. [Google Scholar]
- Maria, D.; Cerqueira, S.; Helber, B.C.; Patrícia, M.; Bueno, F.; José, A.V.; Debora, D.M. Antifungal activity of plant extracts with potential to control plant pathogens in Pine apple. Asian Pac. J. Trop. Biomed. 2016, 6, 26–31. [Google Scholar]
- Whittaker, R.H.; Feeney, P.P. Allelochemics: Chemical Interactions between Species. Science 1971, 171, 757–770. [Google Scholar] [CrossRef]
- Cohen, M.F.; Mazzola, M. Impact of resident bacteria, nitric oxide emission and particle size on root infection by Pythium spp. and R. solani AG-5 in Brassica napus seed meal amended soils. Plant Soil. 2006, 286, 75–86. [Google Scholar] [CrossRef]
- Motisi, N.; Montfort, F.; Doré, T.; Romillac, N.; Lucas, P. Duration of control of two soilborne pathogens following incorporation of above-and below-ground residues of Brassica juncea into soil. Plant Pathol. 2009, 58, 470–478. [Google Scholar] [CrossRef]
- Weerakoon, M.; Izzo, A.; Mazzola, M. Brassica juncea seed meal amendment induces long-term suppressiveness to Pythiumabappressorium under enclosed and open soil incubation conditions. Phytopathology 2010, 100, S134. [Google Scholar]
- Weerakoon, D.M.N.; Reardon, C.L.; Paulitz, T.C.; Antonio, D.; Izzo, A.D.; Mazzola, M. Long-term suppression of Pythiumabappressorium induced by Brassica juncea seed meal amendment is biologically mediated. Soil Biol. Biochem. 2012, 51, 44–52. [Google Scholar] [CrossRef]
- Mazzola, M.; Zhao, X. Brassica juncea seed meal particle size influences chemistry but not soil biology-based suppression of individual agents inciting apple replant disease. Plant Soil. 2010, 337, 313–324. [Google Scholar] [CrossRef]
- Mazzola, M.; Mullinix, K. Comparative field efficacy of management strategies containing Brassica napus seed meal or green manure for the control of apple replant disease. Plant Dis. 2005, 89, 1207–1213. [Google Scholar] [CrossRef] [Green Version]
- Kjaer, A. Glucosinolates in the Cruciferae. In The Biology and Chemistry of the Cruciferae; Vaughan, J.G., Macleod, A.J., Jones, B.M.G., Eds.; Academic Press: London, UK, 1976; pp. 207–219. [Google Scholar]
- Brown, P.D.; Morra, M.J. Control of soil-borne plant pests using glucosinolate-containing plants. Adv. Agron. 1997, 61, 167–231. [Google Scholar]
- Fenwick, G.R.; Heaney, R.K. Glucosinolates and their Breakdown Products in Cruciferous Crops, Foods and Feedingstuffs. Food Chem. 1983, 11, 249–271. [Google Scholar] [CrossRef]
- Angus, J.F.; Gardner, P.A.; Kirkegaard, J.A.; Desmarchelier, J.M. Biofumigation: Isothiocyanates released from Brassica roots inhibit growth of the take-all fungus. Plant Soil. 1994, 162, 107–112. [Google Scholar] [CrossRef]
- Ramirez-Villapudua, J.; Munnecke, D.E. Effect of solar heating and soil amendments of cruciferous residues on Fusariumoxysporum f. sp. conglutinans and other organisms. Phytopathology 1988, 78, 289–295. [Google Scholar]
- Smolinska, U.; Morra, M.J.; Knudsen, G.R.; Brown, P.D. Toxicity of glucosinolate degradation products from Brassica napus seed meal toward Aphanomyces euteiches f. sp. pisi. Phytopathology 1997, 87, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Mazzola, M.; Granatstein, D.M.; Elfving, D.C.; Mullinix, K. Suppression of specific apple root by Brassica napus seed meal amendment regardless of glucosinolate content. Phytopathology 2001, 91, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.A.; Papavizas, G.C. Effect of volatiles from decomposing plant tissues on pigmentation, growth and survival of Rhizoctonia solani. Soil Sci. 1974, 118, 156–163. [Google Scholar] [CrossRef]
- Santini, A.; Biancalani, F.; Biancalani, F.; Barzanti, G.P.; Capretti, P. Pathogenicity of four Phytophthora species on wild cherry and Italian alder seedlings. J. Phytopathol. 2006, 154, 163–167. [Google Scholar] [CrossRef]
- Tewoldemedhin, Y.T.; Mazzola, M.; Spies, C.F.J.; McLeod, A. Characterization of fungi (Fusarium and Rhizoctonia) and oomycetes (Phytophthora and Pythium) associated with apple orchards in South Africa. Eur. J. Plant Pathol. 2011, 130, 215–229. [Google Scholar] [CrossRef]
- Van Den Dool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Shibamoto, T. Retention indices in essential oil analysis. In Capillary Gas Chromatography in Essential Oil Analysis; Sandra, P., Bicchi, C., Eds.; Huethig Verlag: New York, NY, USA, 1987; pp. 259–274. [Google Scholar]
- Longevialle, P. Spectrometrie de Masse des Substances Organiques; Masson: Paris, France, 1981. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (%) | Py. ultimum | Ph. mercuriale | P. citrophthora | F. solani | F. oxysporum | p-Value *** |
|---|---|---|---|---|---|---|
| 5 | 76.42 ± 1.09a * C ** | 71.15 ± 1.57aB | 55.68 ± 1.25aA | 55.19 ± 1.26aA | 100.00 ± 0.00aD | ≤0.001 |
| 10 | 100.00 ± 0.00bB | 75.96 ± 1.11bA | 100.00 ± 0.00bB | 74.50 ± 3.34bA | 100.00 ± 0.00aB | ≤0.001 |
| 15 | 100.00 ± 0.00bB | 77.88 ± 2.48bA | 100.00 ± 0.00bB | 100.00 ± 0.00cB | 100.00 ± 0.00aB | ≤0.001 |
| p-value | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 | nd |
| Treatment | F. solani | F. oxysporum | Py. ultimum | P. citrophthora | p-Value *** | |
|---|---|---|---|---|---|---|
| Root Browning Index | 1W | 2.33 ± 0.58a * A ** | 1.41 ± 0.00Aa | 2.33 ± 0.58Aa | 3.67 ± 0.58Bb | 0.015 |
| 8W | 2.33 ± 0.58Aab | 1.67 ± 0.58Aa | 1.67 ± 0.58Aa | 3.33 ± 0.58Bb | 0.023 | |
| NIC | 1.33 ± 0.58a | 2.00 ± 0.00a | 2.00 ± 0.00a | 2.00 ± 0.00a | nd | |
| IC | 2.00 ± 0.58a | 1.33 ± 0.43a | 2.00 ± 0.00a | 2.33 ± 0.58a | nd | |
| p-value | 0.119 | 0.219 | 0.330 | ≤0.001 | ||
| Sanitary State Index | 1W | 3.00 ± 1.00Aa | 2.01 ± 0.00Aa | 3.00 ± 0.00Aa | 2.33 ± 0.58Aa | 0.160 |
| 8W | 1.67 ± 0.58Aa | 1.67 ± 0.58Aa | 2.33 ± 0.58Aa | 2.00 ± 0.00Aa | 0.363 | |
| NIC | 1.67 ± 0.58a | 2.33 ± 0.58a | 2.33 ± 0.58a | 2.33 ± 0.58a | nd | |
| IC | 2.33 ± 0.58a | 2.67 ± 0.83a | 1.67 ± 0.58a | 2.33 ± 0.58a | nd | |
| p-value | 0.139 | 0.163 | 0.067 | 0.576 | ||
| Height(cm) | 1W | 53.83 ± 0.29Aa | 56.13 ± 9.00Aa | 64.67 ± 9.07Aa | 56.17 ± 5.51Aa | 0.285 |
| 8W | 58.67 ± 20.0Aa | 82.17 ± 8.01Aa | 58.00 ± 17.00Aa | 64.67 ± 2.52Aa | 0.192 | |
| NIC | 62.83 ± 6.79a | 62.83 ± 6.79a | 62.83 ± 6.79a | 62.83 ± 6.79a | nd | |
| IC | 48.00 ± 4.58a | 59.17 ± 13.83a | 61.17 ± 4.75a | 52.50 ± 7.26a | nd | |
| p-value | 0.423 | 0.135 | 0.881 | 0.09 | ||
| Root Weight (g) | 1W | 3.98 ± 0.21Aa | 3.91 ± 0.26abA | 7.18 ± 1.93abA | 6.63 ± 2.21abA | 0.117 |
| 8W | 4.18 ± 1.56Aa | 8.85 ± 1.57Bb | 7.33 ± 1.33Bb | 6.75 ± 0.33abB | 0.014 | |
| NIC | 7.38 ± 2.09b | 7.38 ± 2.09b | 7.38 ± 2.09b | 7.38 ± 2.09b | nd | |
| IC | 2.61 ± 0.78a | 2.88 ± 1.52a | 3.71 ± 0.26a | 3.25 ± 0.36a | nd | |
| p-value | 0.015 | 0.010 | 0.055 | 0.018 |
| Treatment | Py. ultimum | Ph. mercuriale | |
|---|---|---|---|
| Root Browning Index | 1W | 5.00 ± 0.00d * | 1.67 ± 0.58a |
| 8W | 0.33 ± 0.58a | 1.00 ± 0.00a | |
| NIC | 1.33 ± 0.58b | 1.33 ± 0.58a | |
| IC | 2.67 ± 0.58c | 2.67 ± 0.58b | |
| p-value ** | ≤0.001 | 0.017 | |
| Sanitary State Index | 1W | 5.00 ± 0.00b | 2.33 ± 0.58b |
| 8W | 1.00 ± 0.00a | 1.00 ± 0.00a | |
| NIC | 1.33 ± 0.58a | 1.33 ± 0.58ab | |
| IC | 1.67 ± 0.58a | 1.33± 0.58ab | |
| p-value | ≤0.001 | 0.052 | |
| Height (cm) | 1W | 12.00 ± 2.00a | 52.33 ± 5.01a |
| 8W | 63.43 ± 3.10b | 54.50 ± 4.77a | |
| NIC | 90.33 ± 11.02c | 90.33 ± 11.02b | |
| IC | 62.00 ± 6.00b | 80.67 ± 2.08b | |
| p-value | ≤0.001 | ≤0.001 | |
| Root Weight (g) | 1W | 0.34 ± 0.13a | 4.87 ± 0.34a |
| 8W | 5.53 ± 0.11b | 4.36 ± 1.24a | |
| NIC | 11.81 ± 1.32d | 11.81 ± 1.32b | |
| IC | 7.05 ± 0.06c | 11.64 ± 0.45b | |
| p-value | ≤0.001 | ≤0.001 |
| F. oxysporum | F. solani | P. citrophthora | Py. ultimum | Ph. mercuriale | p-Value *** | |
|---|---|---|---|---|---|---|
| EM | 5.41 ± 1.32a * A ** | 6.90 ± 1.33aA | 27.52 ± 7.16bC | 41.57 ± 4.29bD | 15.45 ± 2.43bB | ≤0.001 |
| EA | 15.95 ± 2.35bC | 18.68 ± 1.72bC | 1.22 ± 1.04aA | 29.36 ± 5.06aD | 6.12 ± 2.23aB | ≤0.001 |
| ED | 25.93 ± 1.32cB | 27.30 ± 1.10cB | 67.58 ± 5.14cC | 75.29 ± 5.57cD | 17.78 ± 2.02bA | ≤0.001 |
| p-value | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 |
| Fractions | F. oxysporum | F. solani | P. citrophthora | Py. ultimum | Ph. mercuriale | p-Value *** |
|---|---|---|---|---|---|---|
| Control | 8.03a * | 8.70a | 8.18a | 8.60a | 9.00a | nd |
| F1 | 7.98aB ** | 7.28eA | 8.13aB | 7.58aA | 9.00aC | ≤0.001 |
| F2 | 8.00aD | 6.78fC | 5.33c-fA | 5.90bcdB | 8.90aE | ≤0.001 |
| F3 | 7.95aB | 8.63aC | 8.08aB | 6.55bA | 9.00aD | ≤0.001 |
| F4 | 8.03aC | 7.45deB | 6.80abcB | 3.78fA | 7.00bB | ≤0.001 |
| F5 | 8.03aB | 8.30abB | 5.98bcdA | 5.65bcdA | 8.90aC | ≤0.001 |
| F6 | 7.88aB | 7.73cdB | 8.13aB | 6.23bcA | 8.88aC | ≤0.001 |
| F7 | 8.00aC | 7.20eBC | 8.08aC | 5.25cdeA | 6.38cAB | ≤0.001 |
| F8 | 7.78aC | 7.40deC | 4.15efA | 3.90fA | 6.25cB | ≤0.001 |
| F9 | 7.68aC | 7.23eBC | 3.98fA | 4.15efA | 6.20cB | ≤0.001 |
| F10 | 7.85aD | 7.98bcD | 5.55cdeB | 4.43efA | 6.45cC | ≤0.001 |
| F11 | 7.73aC | 8.55aD | 2.75gA | 3.90fB | 7.18bC | ≤0.001 |
| F12 | 7.93aD | 7.48deC | 3.95fA | 4.83defB | 8.90aE | ≤0.001 |
| F13 | 8.03aC | 8.55aCD | 7.18abB | 6.40bcA | 9.00aD | ≤0.001 |
| F14 | 8.03aB | 8.65aB | 4.55defA | 8.65aB | 9.00aB | ≤0.001 |
| F15 | 8.00aB | 8.73aC | 6.80abcA | 8.28aB | 8.98aC | ≤0.001 |
| F16 | 8.00aB | 7.78cdB | 6.30bcA | 8.23aB | 9.00aC | ≤0.001 |
| F17 | 7.95aB | 8.50aB | 5.75bcdA | 6.28bcA | 9.00aB | ≤0.001 |
| F18 | 7.88aB | 8.03bcB | 4.23efA | 4.13efA | 8.90aC | ≤0.001 |
| p-value | =0.08 | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 |
| Constituents (%) | F7–8 | F9 | F10 | F11 | F12 | F17 | F18 |
|---|---|---|---|---|---|---|---|
| Carboxylic acids | 0.00 | 0.69 | 0.00 | 0.00 | 0.00 | 11.81 | 0.00 |
| Alcohols | 5.73 | 7.63 | 21.79 | 10.98 | 8.19 | 9.61 | 6.49 |
| Aldehydes | 0.63 | 6.32 | 1.12 | 1.65 | 4 | 0.00 | 0.29 |
| Amines | 0.00 | 13.23 | 3.20 | 1.23 | 0.00 | 0.00 | 0.35 |
| ketones | 7.44 | 1.77 | 1.08 | 0.00 | 1.9 | 9.68 | 9.47 |
| Esters | 4.32 | 7.9 | 3.93 | 3.96 | 0.00 | 8.46 | 6.01 |
| Unsaturated hydrocarbons | 43.16 | 26.44 | 25 | 63.43 | 3.19 | 37.93 | 0.00 |
| Saturated hydrocarbons | 1.15 | 1.60 | 1.62 | 2.44 | 0.00 | 0.00 | 10.24 |
| Ethers | 0.00 | 0.00 | 0.00 | 0.00 | 8.21 | 1.27 | 0.33 |
| Dimethyl sulfoxide | 0.00 | 5.68 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Nitriles | 0.82 | 0.00 | 2.7 | 0.52 | 0.00 | 0.00 | 0.00 |
| Amides | 0.00 | 0.00 | 0.00 | 2.19 | 0.00 | 0.00 | 0.00 |
| Indoles | 0.00 | 11.55 | 0.00 | 0.00 | 3.81 | 0.00 | 0.00 |
| Thiocyanates | 0.00 | 3.93 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Pathogens | Fractions | ||||||
|---|---|---|---|---|---|---|---|
| F7–8 | F9 | F10 | F11 | F12 | F17 | F18 | |
| F. oxysporum | 1.56 ± 1.76 a * | 4.36 ± 3.43 ab | 2.41 ± 2.66 a | 17.67 ± 2.34 d | 7.44 ± 2.34 b | 12.56 ± 2.15 c | 1.87 ± 1.87 a |
| F. solani | 14.94 ± 1.63 ab | 16.95 ± 5.26 b | 8.33 ± 0.57 a | 13.58 ± 8.55 ab | 12.35 ± 1.43 ab | 19.75 ± 1.43 b | 7.76 ± 1.45 a |
| P. citrophthora | 25.06 ± 3.56 a | 51.38 ± 15.99 b | 32.11 ± 5.04 a | 50.00 ± 0.76 b | 44.66 ± 5.20 b | 48.09 ± 2.79 b | 48.32 ± 7.23 b |
| Ph. mercuriale | 38.95 ± 3.65 c | 51.74 ± 8.08 d | 48.55 ± 3.34 d | 20.12 ± 0.68 b | 3.99 ± 3.29 a | 13.31 ± 7.57b | 52.03 ± 2.58 d |
| Py. ultimum | 29.10 ± 1.14 b | 31.11 ± 3.27 b | 28.33 ± 4.93 b | 0.00 ± 0.00 a | 45.99 ± 0.00 c | 0.00 ± 0.00 a | 1.11 ± 1.57 a |
| Fractions | Family | Noun | Percentage | Structure |
|---|---|---|---|---|
| F7–8 | Nitrile | 4-Hydroxy-3-(4-methylphenylthio) butanenitrile | 0.82 |  |
| F9 | Thiocyanate | Thiocyanic acid, ethyle | 3.93 |  |
| F10 | Nitrile | Benzeneacetonitrile, 4-fluoro | 2.7 |  |
| F11 | Nitrile | Butanenitrile, 4-hydroxy-3-[(4-methylphenyl)thio]nitrile | 0.52 |  |
| Species | Isolates | Origins | Sampling Years | GenBank Accession Number |
|---|---|---|---|---|
| F. oxysporum | Fo22 | peach | 2013 | MF993097 |
| F. solani | F48 | peach | 2012 | MF993094 |
| Pythium ultimum | P42 | peach | 2013 | MF993110 |
| Po2 | apple | 2012 | MH260594 | |
| Phytopythium mercuriale | Po26 | apple | 2013 | MF993112 |
| Phytophthora citrophthora | P39 | peach | 2013 | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannai, S.; Benfradj, N.; Karoui, A.; Salem, I.B.; Fathallah, A.; M’Hamdi, M.; Boughalleb-M’Hamdi, N. Analysis of Chemical Composition and In Vitro and In Vivo Antifungal Activity of Raphanus raphanistrum Extracts against Fusarium and Pythiaceae, Affecting Apple and Peach Seedlings. Molecules 2021, 26, 2479. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092479
Mannai S, Benfradj N, Karoui A, Salem IB, Fathallah A, M’Hamdi M, Boughalleb-M’Hamdi N. Analysis of Chemical Composition and In Vitro and In Vivo Antifungal Activity of Raphanus raphanistrum Extracts against Fusarium and Pythiaceae, Affecting Apple and Peach Seedlings. Molecules. 2021; 26(9):2479. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092479
Chicago/Turabian StyleMannai, Sabrine, Najwa Benfradj, Ahlem Karoui, Ibtissem Ben Salem, Amel Fathallah, Mahmoud M’Hamdi, and Naima Boughalleb-M’Hamdi. 2021. "Analysis of Chemical Composition and In Vitro and In Vivo Antifungal Activity of Raphanus raphanistrum Extracts against Fusarium and Pythiaceae, Affecting Apple and Peach Seedlings" Molecules 26, no. 9: 2479. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092479