
Rosa platyacantha Schrenk from Kazakhstan—Natural Source of Bioactive Compounds with Cosmetic Significance

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Phytochemical Content and Antioxidant Extracts from Various Parts of R. platyacantha
2.2. Chromatographic Analysis of R. platyacantha Extracts
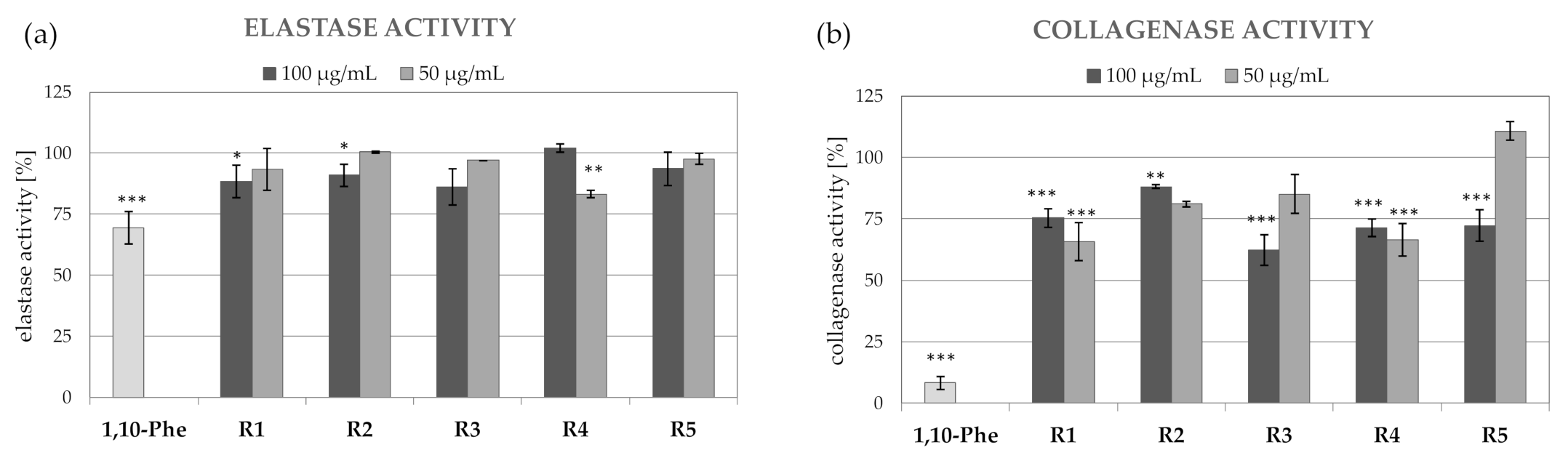
2.3. Anti-Collagenase and Anti-Elastase Activity of R. platyacantha Extracts
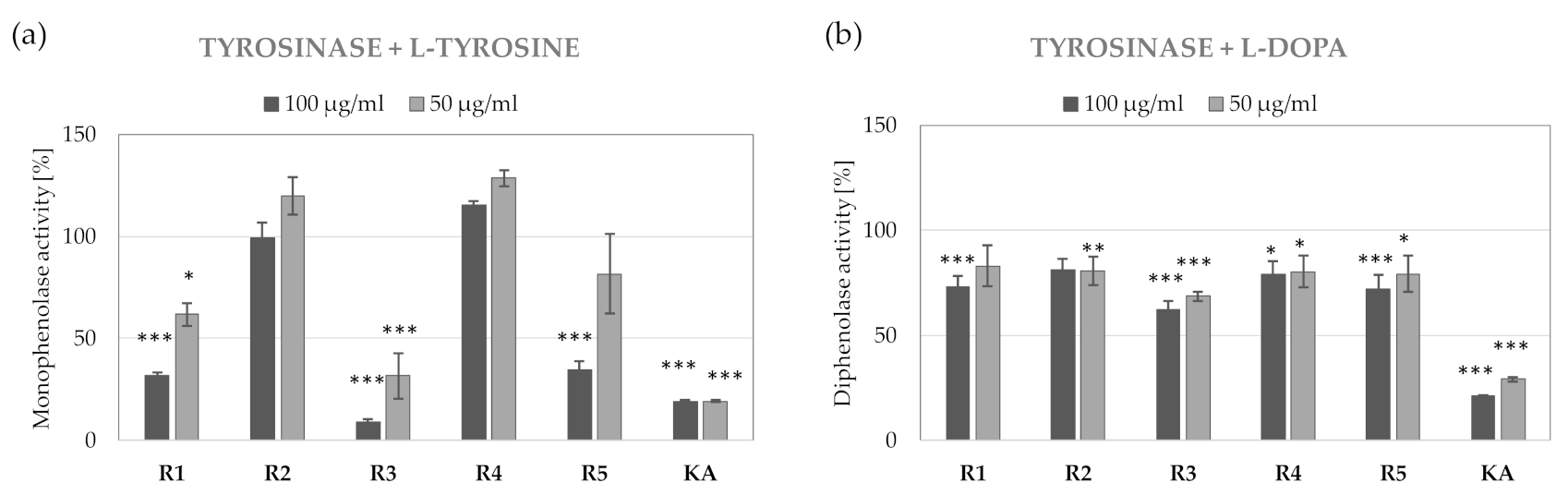
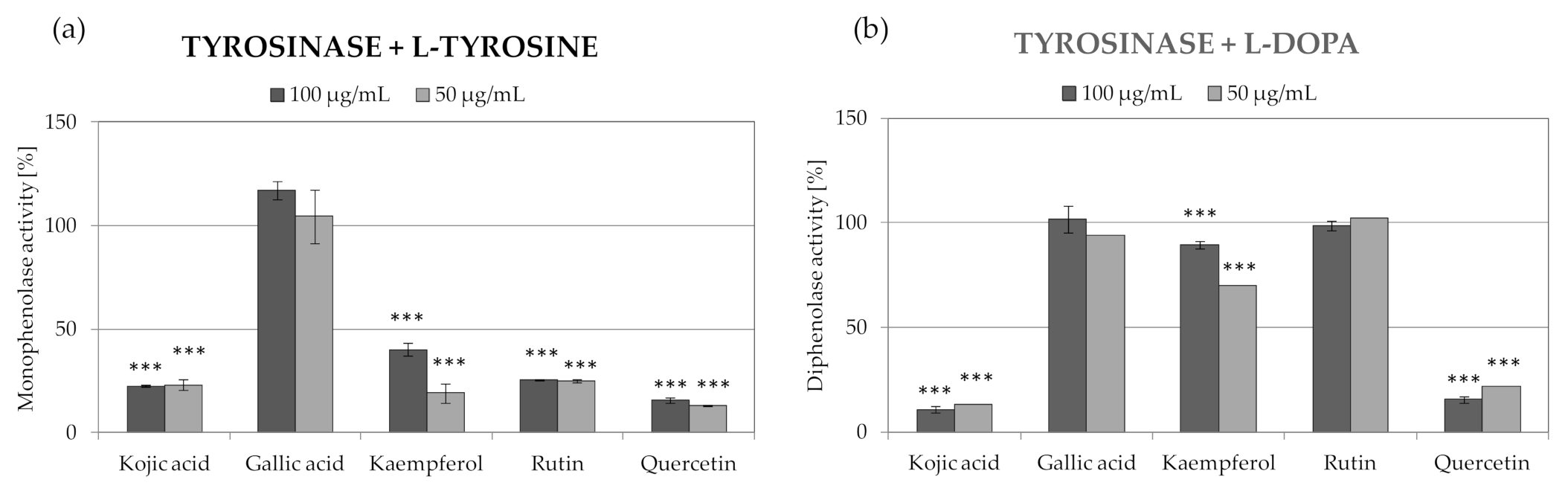
2.4. Anti-Tyrosinase Activity of R. platyacantha Extracts
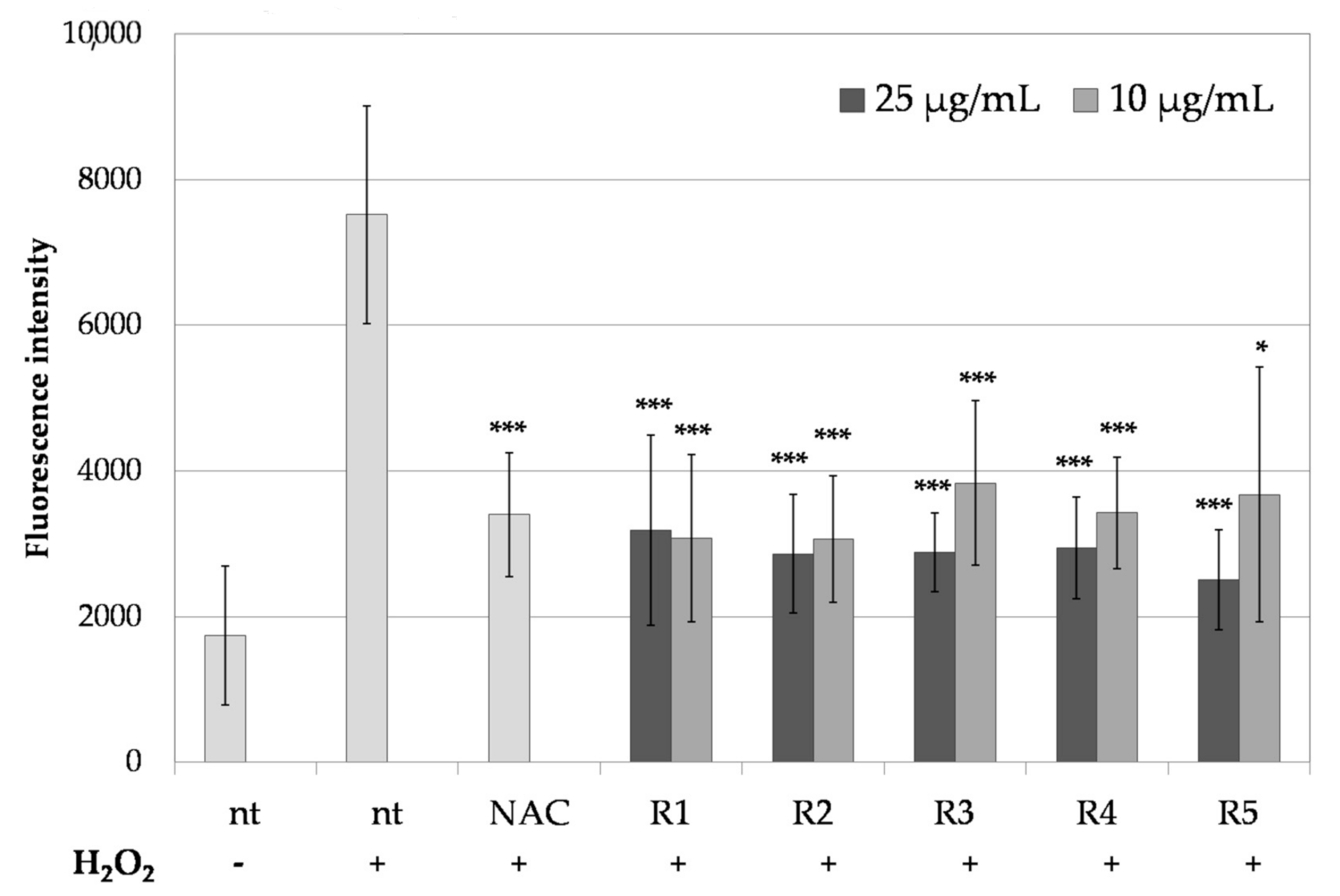
2.5. Anti-Melanoma Activity of R. platyacantha Extracts In Vitro
2.6. Fractionation of R3 Extract and Chromatographic Analysis of the Fractions
2.7. Antioxidant and Anti-Melanoma Activities of Fractions C-I Separated from R3 Extract
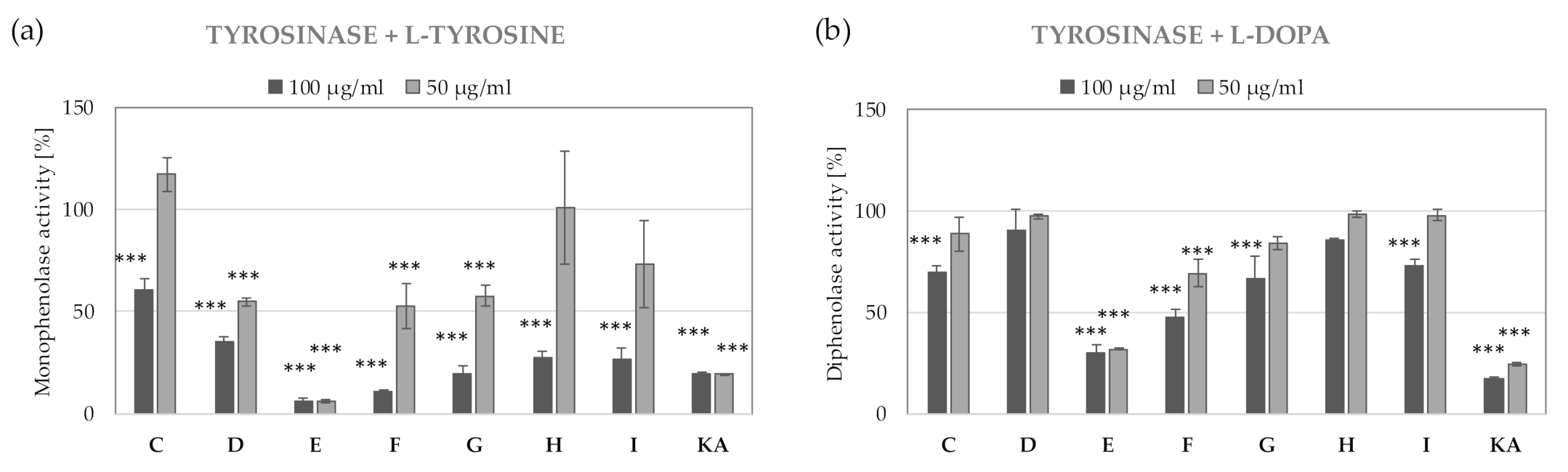
2.8. Identification of Tyrosinase Inhibitors from E and F Fractions Separated from R3 Extract
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material
3.3. Extraction Procedure and Fractionation
3.4. Total Phenolics and Flavonoids Content
3.5. Chromatographic Analysis
3.6. DPPH Radical Scavenging Assay
3.7. ABTS Radical Scavenging Assay
3.8. Tyrosinase Inhibitory Assay
3.9. Elastase Inhibitory Assay
3.10. Collagenase Inhibitory Assay
3.11. In Vitro Cytotoxicity Assay
3.12. Detection of Intracellular ROS using H2DCFDA
3.13. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Saint-Oyant, L.H.; Ruttink, T.; Hamama, L.; Kirov, I.; Lakhwani, D.; Zhou, N.N.; Bourke, P.M.; Daccord, N.; Leus, L.; Schulz, D.; et al. A high-quality genome sequence of Rosa chinensis to elucidate ornamental traits. Nat. Plants 2018, 4, 473–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsioutsiou, E.E.; Giordani, P.; Hanlidou, E.; Biagi, M.; De Feo, V.; Cornara, L. Ethnobotanical Study of Medicinal Plants Used in Central Macedonia, Greece. Evid. Based Complement. Altern. Med. 2019, 2019, 4513792. [Google Scholar] [CrossRef] [PubMed]
- Hayta, S.; Polat, R.; Selvi, S. Traditional uses of medicinal plants in ElazIǧ (Turkey). J. Ethnopharmacol. 2014, 154, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Phetcharat, L.; Wongsuphasawat, K.; Winther, K. The effectiveness of a standardized rose hip powder, containing seeds and shells of Rosa canina, on cell longevity, skin wrinkles, moisture, and elasticity. Clin. Interv. Aging 2015, 10, 1849–1856. [Google Scholar]
- Moradkhani, S.; Rezaei-Dehghanzadeh, T.; Nili-Ahmadabadi, A. Rosa persica hydroalcoholic extract improves cadmium-hepatotoxicity by modulating oxidative damage and tumor necrosis factor-alpha status. Environ. Sci. Pollut. Res. 2020, 27, 31259–31268. [Google Scholar] [CrossRef]
- Changizi Ashtiyani, S.; Najafi, H.; Jalalvandi, S.; Hosseinei, F. Protective effects of Rosa canina L fruit extracts on renal disturbances induced by reperfusion injury in rats. Iran. J. Kidney Dis. 2013, 7, 290–298. [Google Scholar]
- Komiazyk, M.; Palczewska, M.; Sitkiewicz, I.; Pikula, S.; Groves, P. Neutralization of cholera toxin by Rosaceae family plant extracts. BMC Complement. Altern. Med. 2019, 19, 140. [Google Scholar] [CrossRef] [Green Version]
- Cheng, B.C.Y.; Fu, X.Q.; Guo, H.; Li, T.; Wu, Z.Z.; Chan, K.; Yu, Z.L. The genus Rosa and arthritis: Overview on pharmacological perspectives. Pharmacol. Res. 2016, 114, 219–234. [Google Scholar] [CrossRef]
- Taghizadeh, M.; Rashidi, A.A.; Taherian, A.A.; Vakili, Z.; Mehran, M. The Protective Effect of Hydroalcoholic Extract of Rosa canina (Dog Rose) Fruit on Liver Function and Structure in Streptozotocin-Induced Diabetes in Rats. J. Diet. Suppl. 2018, 15, 624–635. [Google Scholar] [CrossRef]
- Ninomiya, K.; Matsuda, H.; Kubo, M.; Morikawa, T.; Nishida, N.; Yoshikawa, M. Potent anti-obese principle from Rosa canina: Structural requirements and mode of action of trans-tiliroside. Bioorg. Med. Chem. Lett. 2007, 17, 3059–3064. [Google Scholar] [CrossRef]
- Turan, I.; Demir, S.; Kilinc, K.; Yaman, S.O.; Misir, S.; Kara, H.; Genc, B.; Mentese, A.; Aliyazicioglu, Y.; Deger, O. Cytotoxic effect of Rosa canina extract on human colon cancer cells through repression of telomerase expression. J. Pharm. Anal. 2018, 8, 394–399. [Google Scholar] [CrossRef]
- Sargin, S.A. Potential anti-influenza effective plants used in Turkish folk medicine: A review. J. Ethnopharmacol. 2021, 265, 113319. [Google Scholar] [CrossRef]
- Jiménez, S.; Gascón, S.; Luquin, A.; Laguna, M.; Ancin-Azpilicueta, C.; Rodríguez-Yoldi, M.J. Rosa canina extracts have antiproliferative and antioxidant effects on caco-2 human colon cancer. PLoS ONE 2016, 11, e0159136. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, R.; Barros, L.; Calhelha, R.C.; Carvalho, A.M.; Queiroz, M.J.R.P.; Ferreira, I.C.F.R. Bioactivity of Different Enriched Phenolic Extracts of Wild Fruits from Northeastern Portugal: A Comparative Study. Plant Foods Hum. Nutr. 2014, 69, 37–42. [Google Scholar] [CrossRef]
- Lee, Y.H.; Jung, M.G.; Kang, H.B.; Choi, K.C.; Haam, S.; Jun, W.; Kim, Y.J.; Cho, H.Y.; Yoon, H.G. Effect of anti-histone acetyltransferase activity from Rosa rugosa Thunb. (Rosaceae) extracts on androgen receptor-mediated transcriptional regulation. J. Ethnopharmacol. 2008, 118, 412–417. [Google Scholar] [CrossRef]
- Cosmetic Ingredient Database (CosIng). Available online: https://ec.europa.eu/growth/sectors/cosmetics/cosing_en (accessed on 17 March 2021).
- Kwak, C.S.; Yang, J.; Shin, C.Y.; Chung, J.H. Rosa multiflora Thunb Flower Extract Attenuates Ultraviolet-Induced Photoaging in Skin Cells and Hairless Mice. J. Med. Food. 2020, 23, 988–997. [Google Scholar] [CrossRef]
- Shin, E.J.; Han, A.R.; Lee, M.H.; Song, Y.R.; Lee, K.M.; Nam, T.G.; Lee, P.; Lee, S.Y.; Lim, T.G. Extraction conditions for Rosa gallica petal extracts with anti-skin aging activities. Food Sci. Biotechnol. 2019, 28, 1439–1446. [Google Scholar] [CrossRef]
- Fujii, T.; Ikeda, K.; Saito, M. Inhibitory effect of rose hip (Rosa canina L.) on melanogenesis in mouse melanoma cells and on pigmentation in brown guinea pigs. Biosci. Biotechnol. Biochem. 2011, 75, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.R.; Lim, W.C.; Han, A.; Lee, M.H.; Shin, E.J.; Lee, K.M.; Nam, T.G.; Lim, T.G. Rose Petal Extract (Rosa gallica) Exerts Skin Whitening and Anti-Skin Wrinkle Effects. J. Med. Food 2020, 23, 870–878. [Google Scholar] [CrossRef]
- Ren, G.; Xue, P.; Sun, X.; Zhao, G. Determination of the volatile and polyphenol constituents and the antimicrobial, antioxidant, and tyrosinase inhibitory activities of the bioactive compounds from the by-product of Rosa rugosa Thunb. var. plena Regal tea. BMC Complement. Altern. Med. 2018, 18, 307. [Google Scholar] [CrossRef] [Green Version]
- Roman, I.; Stǎnilǎ, A.; Stǎnilǎ, S. Bioactive compounds and antioxidant activity of Rosa canina L. biotypes from spontaneous flora of Transylvania. Chem. Cent. J. 2013, 7, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dani, K.G.S.; Fineschi, S.; Michelozzi, M.; Trivellini, A.; Pollastri, S.; Loreto, F. Diversification of petal monoterpene profiles during floral development and senescence in wild roses: Relationships among geraniol content, petal colour, and floral lifespan. Oecologia 2020. [CrossRef] [PubMed]
- Mármol, I.; Sánchez-De-Diego, C.; Jiménez-Moreno, N.; Ancín-Azpilicueta, C.; Rodríguez-Yoldi, M. Therapeutic applications of rose hips from different Rosa species. Int. J. Mol. Sci. 2017, 18, 1137. [Google Scholar] [CrossRef]
- Christensen, L. Galactolipids as Potential Health Promoting Compounds in Vegetable Foods. Recent Patents Food Nutr. Agric. 2012, 1, 50–58. [Google Scholar]
- Tang, Y.; Zhu, Z.Y.; Liu, Y.; Sun, H.; Song, Q.Y.; Zhang, Y. The chemical structure and anti-aging bioactivity of an acid polysaccharide obtained from rose buds. Food Funct. 2018, 9, 2300–2312. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.W.; Du, Y.E.; Qi, Y.; Lee, J.S.; Goo, N.; Koo, B.K.; Bae, H.J.; Ryu, J.H.; Jang, D.S. New Depsides and Neuroactive Phenolic Glucosides from the Flower Buds of Rugosa Rose (Rosa rugosa). J. Agric. Food Chem. 2019, 67, 7289–7296. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.K.; Lee, D.Y.; Lee, H.; Kim, D.O.; Baek, N.I.; Kim, Y.E.; Kim, H.Y. Flavonoids from the buds of Rosa damascena inhibit the activity of 3-hydroxy-3-methylglutaryl-coenzyme a reductase and angiotensin I-converting enzyme. J. Agric. Food Chem. 2010, 58, 882–8866. [Google Scholar] [CrossRef] [PubMed]
- Nowak, R.; Gawlik-Dziki, U. Polyphenols of Rosa L. leaves extracts and their radical scavenging activity. Z. Naturforsch C J. Biosci. 2007, 62, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Polumackanycz, M.; Kaszuba, M.; Konopacka, A.; Marzec-Wróblewska, U.; Wesolowski, M.; Waleron, K.; Buciński, A.; Viapiana, A. Phenolic Composition and Biological Properties of Wild and Commercial Dog Rose Fruits and Leaves. Molecules 2020, 25, 5272. [Google Scholar] [CrossRef]
- Sermukhamedova, O.; Sakipova, Z.; Ternynko, I.; Gemedzhieva, N. Representatives of motherwort genus (Leonurus SPP.): Aspects of pharmacognostic features and relevance of new species application. Acta Poloniae Pharm.Drug Res. 2017, 74, 31–40. [Google Scholar]
- Kartbaeva, E.B.; Donald, G.R.; Sakipova, Z.B.; Ibragimova, L.N.; Bekbolatova, E.N.; Ternynko, I.I.; Fernandes, P.D.; Boylan, F. Antinociceptive activity of cistanche salsa stolons, growing in the republic of Kazakhstan. Rev. Bras. Farmacogn. 2017, 27, 587–591. [Google Scholar] [CrossRef]
- Di Ferdinando, M.; Brunettia, C.; Agatib, G.; Tattini, M. Multiple functions of polyphenols in plants inhabiting unfavorable Mediterranean areas. Environ. Exp. Bot. 2014, 103, 107–116. [Google Scholar] [CrossRef]
- Zillich, O.V.; Schweiggert-Weisz, U.; Eisner, P.; Kerscher, M. Polyphenols as active ingredients for cosmetic products. Int. J. Cosmet. Sci. 2015, 37, 455–464. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Wittgen, H.G.; van Kempen, L.C. Reactive Oxygen Species in Melanoma and Its Therapeutic Implications. Melanoma Res. 2007, 17, 400–409. [Google Scholar] [CrossRef]
- Poljšak, B.; Dahamane, R. Free radicals and extrinsic skin aging. Dermatol. Res. Pract. 2012, 2012, 135206. [Google Scholar] [CrossRef] [Green Version]
- Boukamp, P.; Petrussevska, R.T.; Breitkreutz, D.; Hornung, J.; Markham, A.; Fusenig, N.E. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 1988, 106, 761–771. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.; Fernandes, E.; Lima, J.L.F.C. Fluorescence probes used fordetection of reactive oxygen species. J. Biochem. Biophys. 2005, 65, 45–80. [Google Scholar] [CrossRef]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-Acetylcysteine as an antioxidant and disulphide breaking agent: The reasons why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef]
- Cendrowski, A.; Ścibisz, I.; Kieliszek, M.; Kolniak-Ostek, J.; Mitek, M. UPLC-PDA-Q/TOF-MS Profile of Polyphenolic Compounds of Liqueurs from Rose Petals (Rosa rugosa). Molecules 2017, 22, 1832. [Google Scholar] [CrossRef] [Green Version]
- Hosni, K.; Chrif, R.; Zahed, N.; Abid, I.; Medfei, W.; Sebei, H.; Brahim, N.B. Fatty acid and phenolic constituents of leaves, flowers and fruits of tunisian dog rose (Rosa canina L.). Riv. Ital. Sostanze Gr. 2010, 87, 117–123. [Google Scholar]
- Abdel-Hameed, E.S.S.; Bazaid, S.A.; Salman, M.S. Characterization of the phytochemical constituents of Taif rose and its antioxidant and anticancer activities. Biomed. Res. Int. 2013, 2013, 345465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikbulatova, T.N.; Beisekova, K.D. Chemical composition of the fruit of Rosa platyacantha. Khimiya Prir. Soedin. 1979, 3, 420–421. [Google Scholar]
- Bikbulatova, T.N.; Erzhanova, M.S.; Terent’ev, P.B.; Beisekova, K.D.; Seifullina, A.A. hydrolyzable tannin substance from the fruit of Rosa platyacantha. Chem. Nat. Compd. 1985, 21, 789–791. [Google Scholar] [CrossRef]
- Hussein, S.A.M.; Barakat, H.H.; Merfort, I.; Nawwar, M.A.M. Tannins from the leaves of Punica granatum. Phytochemistry 1997, 45, 819–823. [Google Scholar] [CrossRef]
- Wang, X.; Xing, D.; Ding, Y.; Chen, Y.; Meng, Z.; Du, L. Determination and pharmacokinetic study of brevifolin in rat after ig administration of pomegranate leaf extract. Chin. Pharmacol. Bull. 2005, 2, 369–372. [Google Scholar]
- Sójka, M.; Janowski, M.; Grzelak-Błaszczyk, K. Stability and transformations of raspberry (Rubus idaeus L.) ellagitannins in aqueous solutions. Eur. Food Res. Technol. 2019, 245, 1113–1122. [Google Scholar] [CrossRef] [Green Version]
- Fujii, T.; Saito, M. Inhibitory effect of quercetin isolated from rose hip (Rosa canina L.) against melanogenesis by mouse melanoma cells. Biosci. Biotechnol. Biochem. 2009, 73, 1989–1993. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Imokawa, G.; Ishida, K. Biological mechanisms underlying the ultraviolet radiation-induced formation of skin wrinkling and sagging I: Reduced skin elasticity, highly associated with enhanced dermal elastase activity, triggers wrinkling and sagging. Int. J. Mol. Sci. 2015, 16, 7753–7775. [Google Scholar] [CrossRef] [Green Version]
- Tundis, R.; Loizzo, M.R.; Bonesi, M.; Menichini, F. Potential role of natural compounds against skin aging. Curr. Med. Chem. 2015, 22, 1515–1538. [Google Scholar] [CrossRef]
- Jiratchayamaethasakul, C.; Ding, Y.; Hwang, O.; Im, S.-T.; Jang, Y.; Myung, S.-W.; Lee, J.M.; Kim, H.-S.; Ko, S.-C.; Lee, S.H. In vitro screening of elastase, collagenase, hyaluronidase, and tyrosinase inhibitory and antioxidant activities of 22 halophyte plant extracts for novel cosmeceuticals. Fish Aquatic. Sci. 2020, 23, 6. [Google Scholar] [CrossRef]
- Mawarni, E.; Ginting, C.N.; Chiuman, L.; Girsang, E.; Handayani, R.; Siwianti, A.; Widowati, W. Antioxidant and Elastase Inhibitor Potential of Petals and Receptacle of Rose Flower (Rosa damascena). Pharm. Sci. Res. 2020, 7, 105–113. [Google Scholar] [CrossRef]
- Choi, E.K.; Guo, H.; Choi, J.K.; Jang, S.K.; Shin, K.; Cha, Y.S.; Choi, Y.; Seo, D.W.; Lee, Y.B.; Joo, S.S.; et al. Extraction conditions of white rose petals for the inhibition of enzymes related to skin aging. Lab. Anim. Res. 2015, 31, 148–152. [Google Scholar] [CrossRef] [Green Version]
- Thring, T.S.; Hili, P.; Naughton, D.P. Anti-collagenase, anti-elastase and anti-oxidant activities of extracts from 21 plants. BMC Complement. Altern Med. 2009, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Kanlayavattanakul, M.; Lourith, N. Skin hyperpigmentation treatment using herbs: A review of clinical evidences. J. Cosmet. Laser Ther. 2018, 20, 123–131. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [Green Version]
- Jensen, J.D.; Dunn, J.H.; Luo, Y.; Liu, W.; Fujita, M.; Dellavalle, R.P. Ellagic acid inhibits melanoma growth in vitro. Dermatol. Rep. 2011, 3, e36. [Google Scholar] [CrossRef]
- Lo, C.; Lai, T.Y.; Yang, J.H.; Yang, J.S.; Ma, Y.S.; Weng, S.W.; Chen, Y.Y.; Lin, J.G.; Chung, J.G. Gallic acid induces apoptosis in A375.S2 human melanoma cells through caspase-dependent and -independent pathways. Int. J. Oncol. 2010, 37, 377–385. [Google Scholar]
- Yang, J.; Xiao, P.; Sun, J.; Guo, L. Anticancer effects of kaempferol in A375 human malignant melanoma cells are mediated via induction of apoptosis, cell cycle arrest, inhibition of cell migration and downregulation of m-TOR/PI3K/AKT pathway. J. BUON 2018, 23, 218–223. [Google Scholar]
- Harris, Z.; Donovan, M.G.; Branco, G.M.; Limesand, K.H.; Burd, R. Quercetin as an Emerging Anti-Melanoma Agent: A Four-Focus Area Therapeutic Development Strategy. Front. Nutr. 2016, 3, 48. [Google Scholar] [CrossRef]
- Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; Ahmad, N. Melanoma chemoprevention: Current status and future prospects. Photochem. Photobiol. 2017, 93, 975–989. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Li, S.-Y.; Huang, X.-E.; Cui, J.-J.; Zhao, T.; Zhang, H. Inhibition of tumor growth in vitro by a combination of extracts from Rosa roxburghii Tratt and fagopyrum cymosum. Asian Pac. J. Cancer Prev. 2012, 13, 2409–2414. [Google Scholar] [CrossRef] [Green Version]
- Fukumoto, L.R.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef]
- Matejić, J.S.; Džamić, A.M.; Mihajilov-Krstev, T.M.; Ranđelović, V.N.; Krivošej, Z.D.; Marin, P.D. Total phenolic and flavonoid content, antioxidant and antimicrobial activity of extracts from Tordylium maximum. J. Appl. Pharm. Sci. 2013, 3, 55–59. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Wang, Y.; Hao, M.-M.; Sun, Y.; Wang, L.-F.; Wang, H.; Zhang, Y.-J.; Li, H.-Y.; Zhuang, P.-W.; Yang, Z. Synergistic Promotion on Tyrosinase Inhibition by Antioxidants. Molecules 2018, 23, 106. [Google Scholar] [CrossRef] [Green Version]
- Uchida, R.; Ishikawa, S.; Tomoda, H. Inhibition of tyrosinase activity and melanin pigmentation by 2-hydroxytyrosol. Acta Pharm. Sin. B 2014, 4, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Horng, C.; Wu, H.C.; Chiang, N.N.; Lee, C.F.; Huang, Y.S.; Wang, H.Y.; Yang, J.S.; Chen, F.A. Inhibitory effects of burdock leaves on elastase and tyrosinase activity. Exp. Ther. Med. 2017, 14, 3247–3252. [Google Scholar] [CrossRef] [Green Version]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Wu, D.; Yotnda, P. Production and detection of reactive oxygen species (ROS) in cancers. J. Vis. Exp. 2011, 57, 3357. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| R1 | R2 | R3 | R4 | R5 | Vitamin C | |
|---|---|---|---|---|---|---|
| Total phenolic (mg GAE/g dw) | 8.61 ± 0.18 | 14.53 ± 0.18 | 13.30 ± 0.16 | 14.05 ± 0.28 | 13.68 ± 0.27 | - |
| Flavonoids (mg QE/g dw) | 2.42 ± 0.05 | 2.45 ± 0.03 | 2.42 ± 0.05 | 2.49 ± 0.09 | 2.03 ± 0.06 | - |
| DPPH Scavenging (IC50, µg/mL) | 2.77 ± 0.05 | 1.68 ± 0.25 | 1.50 ± 0.19 | 1.59 ± 0.14 | 1.10 ± 0.34 | 0.96 ± 0.05 |
| ABTS Scavenging (IC50, µg/mL) | 16.16 ± 1.26 | 7.16 ± 0.22 | 10.83 ± 0.85 | 9.89 ± 0.83 | 9.21 ± 0.54 | 0.97 ± 0.06 |
| No | Retention Time | Name | Formula | Molecular Ion [M − H]‘ | Fragmentation Ions | R1 | R2 | R3 | R4 | R5 |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1.920 | Quinic acid | C7H12O6 | 191.0581 | 173.0438; 127.0426; 109.0287 | +++ | +++ | ++ | +++ | +++ |
| 2 | 2.507 | Citric acid | C₆H₈O₇ | 191.0362 | 173.0119; 111.0086 | + | + | + | + | + |
| 3 | 2.955 | Gallic acid glucoside isomer | C13H16O10 | 331.0683 | 271.0432; 169.0143; 125.0230 | + | + | - | + | + |
| 4 | 3.396 | Homoisocitric acid | C7H10O7 | 205.0373 | 173.0207; 155.0111; 111.0145 | + | - | - | - | - |
| 5 | 4.092 | Gallic acid | C7H6O5 | 169.0147 | 125.0258; 107.0141 | + | + | ++ | + | ++ |
| 6 | 4.530 | Theogallin | C14H16O10 | 343.0685 | 191,0497; 127.0389 | + | +++ | + | +++ | + |
| 7 | 6.495 | Gallic acid glucoside isomer | C13H16O10 | 331.0659 | 169.0139; 125.0222 | + | + | + | + | + |
| 8 | 7.826 | Gallic acid derivative | C23H19O18 | 581.0449 | 313.0490; 169.0104 | + | - | + | - | + |
| 9 | 9.877 | Methoxygallic glucoside isomer | C14H18O10 | 345.0820 | 183.0275; 124.0151 | + | - | + | - | - |
| 10 | 10.127 | Chlorogenic acid | C16H18O9 | 353.0867 | 191.0373; 135.0308; 109.0238 | - | + | - | ++ | ++ |
| 11 | 11.639 | Methoxygallic acid isomer | C8H10O6 | 183.0453 | 168.0215; 124.0238 | ++ | + | +++ | ++ | +++ |
| 12 | 13.378 | Ellagic acid derivative | C34H26O22 | 783.0488 | 300.9905; 275.0111; 249.0302 | + | - | + | - | + |
| 13 | 16.619 | Ellagitanin derivative | C30H24O25 | 785.0836 | 300.9934; 275.0111; 249.0375; 169.0107 | + | + | + | - | - |
| 14 | 17.214 | Ellagitannin derivative | C34H26O22 | 785.0836 | 300.9981; 275.0185; 249.0428; 162.0121; 125.0163 | + | - | - | - | - |
| 15 | 17.255 | Cryptochlorogenic acid | C16H18O9 | 353.0496 | 191.0380; 179.0113; 173.0244; 135.0268 | + | + | - | + | - |
| 16 | 17.559 | Strictinin | C27H22O18 | 633.0723 | 300.9669; 247.9903; 249.0121; 168.9972; 125.0082 | - | + | + | + | + |
| 17 | 19.218 | Brevifolin carboxylic acid | C13H8O8 | 291.0133 | 247.0116; 205.0041 | ++ | ++ | + | +++ | + |
| 18 | 20.860 | Brevifolin | C12H7O6 | 247.0235 | 201.0169; 190.0258; 173.0207; 145.0278; 135.0421 | + | + | + | + | + |
| 19 | 21.130 | Methyl brevifolincarboxylate | C14H10O8 | 305.0290 | 273.0069; 245.0075; 217.0119; 189.0166; 161.0237; 145.0269; 133.0273; 117.0349 | +++ | ++ | ++ | +++ | ++ |
| 20 | 23.021 | Quercetin galloylglucoside isomer | C28H24O16 | 615.1014 | 463.0872; 300.0281; 271.0297; 255.0269; 169.0140; 151.0020; 124.0151; 107.0091 | + | + | + | + | + |
| 21 | 23.281 | Quercetin galloylglucoside isomer | C28H24O16 | 615.1014 | 463.0900; 300.0276; 271.0283; 255.0223; 169.0140; 150.9999; 124.0139 | + | - | - | - | + |
| 22 | 23.951 | Ellagic acid glucoside | C20H16O13 | 463.0584 | 300.9994; 226.9924; 200.0078; 173.0221; 145.0286; 117.0374 | - | - | ++ | + | ++ |
| 23 | 23.997 | Quercetin 3-O glucoside | C21H20O12 | 463.0896 | 300.0254; 271.0243; 255.0273; 179.0005; 151.0029; 135.0105; 108.0183 | + | + | + | - | - |
| 24 | 24.351 | Ellagic acid | C14H6O8 | 301.0003 | 283.9966; 245.0068; 201.0123; 173.0243; 145.0273; 117.0332 | + | - | - | - | - |
| 25 | 25.490 | Quercetin glucuronide | C21H18O13 | 477.0700 | 301.0350; 255.0315; 178.9994; 151.0048; 107.0153 | +++ | + | + | + | + |
| 26 | 26.172 | Digalloylglucoside | C22H12O13 | 483.0237 | 301.0079; 285.0134; 270.9967; 228.0068; 173.0232; 144.0312; 117.0321 | + | - | + | - | + |
| 27 | 26.326 | Quercetin 7-O-glucoside | C21H20O12 | 463.0896 | 300.0280; 257.0421; 179.0005; 151.0045; 107.0183 | + | - | + | - | + |
| 28 | 28.380 | Quercetin galloylglucoside isomer | C28H24O16 | 615.1014 | 301.0386; 255.0328; 179.0003; 169.0139; 151.0065; 125.0231 | +++ | + | + | + | + |
| 29 | 29.197 | Rutin | C27H30O16 | 609.1278 | 463.0905; 300.0289; 271.0273; 255.0265; 179.0031; 151.0050; 107.0139 | ++ | - | - | - | + |
| 30 | 31.014 | Kaempferol rutinoside | C27H30O15 | 593.1105 | 285.0352; 255.0270; 227.0305; 150.9958; 145.0285; 119.0489 | + | - | + | + | - |
| 31 | 34.432 | Kaempferol | C15H10O6 | 285.0388 | 229.0512; 150.9980; 107.0111 | + | - | - | - | - |
| R1 | R2 | R3 | R4 | R5 | |
|---|---|---|---|---|---|
| HaCaT | >500 | 180.60 | 241.40 | 304.30 | 293.90 |
| A365 | >500 | 120.40 | 97.31 | 199.50 | 72.90 |
| SH4 | >500 | 149.70 | 169.00 | 129.90 | 142.00 |
| B16F10 | >500 | 226.10 | 187.30 | 136.80 | 174.20 |
| No | Retention Time | Name | Formula | C | D | E | F | G | H | I |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1.920 | Quinic acid | C7H12O6 | - | - | - | - | + | + | + |
| 2 | 2.507 | Citric acid | C₆H₈O₇ | - | - | - | - | - | - | - |
| 3 | 2.955 | Gallic acid glucoside isomer | C13H16O10 | - | - | - | + | + | - | - |
| 4 | 3.396 | Homoisocitric acid | C7H10O7 | - | - | - | - | + | - | - |
| 5 | 4.092 | Gallic acid | C7H6O5 | - | +++ | ++ | + | + | + | + |
| 6 | 4.530 | Theogallin | C14H16O10 | - | - | - | - | + | + | + |
| 7 | 6.495 | Gallic acid glucoside isomer | C13H16O10 | - | - | + | + | - | - | - |
| 8 | 7.826 | Gallic acid derivative | C23H19O18 | - | - | - | + | - | - | - |
| 9 | 9.877 | Methoxygallic acid glucoside isomer | C14H18O10 | - | - | ++ | + | - | - | - |
| 10 | 10.127 | Chlorogenic acid | C16H18O9 | - | - | - | + | - | - | - |
| 11 | 11.639 | Methoxygallic acid isomer | C8H10O6 | +++ | +++ | ++ | - | + | + | + |
| 12 | 13.378 | Ellagic acid derivative | C34H26O22 | - | - | + | + | - | - | - |
| 13 | 16.619 | Ellagitanin derivative | C30H24O25 | - | - | + | - | - | - | - |
| 14 | 17.214 | Ellagitannin derivative | C34H26O22 | - | - | - | + | - | + | - |
| 15 | 17.255 | Cryptochlorogenic | C16H18O9 | - | - | - | - | - | - | - |
| 16 | 17.559 | Strictinin | C27H22O18 | - | - | - | + | - | - | - |
| 17 | 19.218 | Brevifolin carboxylic acid | C13H8O8 | - | - | - | + | + | + | + |
| 18 | 20.860 | Brevifolin | C12H7O6 | - | - | + | - | + | + | + |
| 19 | 21.130 | Methyl brevifolincarboxylate | C14H10O8 | - | - | + | + | + | + | + |
| 20 | 23.021 | Quercetin galloylglucoside isomer | C28H24O16 | - | - | + | - | - | - | - |
| 21 | 23.281 | Quercetin galloylglucoside isomer | C28H24O16 | - | - | + | - | - | - | - |
| 22 | 23.951 | Ellagic acid glucoside | C20H16O13 | - | - | - | - | - | - | - |
| 23 | 23.997 | Quercetin 3-O glucoside | C21H20O12 | - | - | + | + | + | + | + |
| 24 | 24.351 | Ellagic acid | C14H6O8 | - | - | - | - | - | - | - |
| 25 | 25.490 | Quercetin glucuronide | C21H18O13 | - | - | - | + | + | + | + |
| 26 | 26.172 | Digalloylglucoside | C22H12O13 | - | - | + | - | - | - | - |
| 27 | 26.326 | Quercetin 7-O-glucoside | C21H20O12 | - | - | - | - | - | - | - |
| 28 | 28.380 | Quercetin galloylglucoside isomer | C28H24O16 | - | - | + | + | + | - | + |
| 29 | 29.197 | Rutin | C27H30O16 | - | - | + | + | - | - | - |
| 30 | 31.014 | Kaempferol rutinoside | C27H30O15 | - | - | + | + | - | - | - |
| 31 | 34.432 | Kaempferol | C15H10O6 | - | - | - | - | - | - | - |
| C | D | E | F | G | H | I | |
|---|---|---|---|---|---|---|---|
| DPPH Scavenging | 11.99 ± 0.96 | 2.60 ± 0.10 | 2.17 ± 0.04 | 6.86 ± 0.50 | 3.87 ± 0.26 | 5.89 ± 0.60 | 5.14 ± 0.57 |
| ABTS Scavenging | 2 510.00 ± 449.81 | 4.77 ± 0.10 | 4.30 ± 0.55 | 13.87 ± 0.05 | 8.52 ± 0.27 | 11.14 ± 0.91 | 10.39 ± 0.19 |
| C | D | E | F | G | H | I | |
|---|---|---|---|---|---|---|---|
| HaCaT | 251.50 | 137.60 | 190.70 | >500 | >500 | >500 | >500 |
| A375 | 170.60 | 70.30 | 205.80 | >500 | >500 | >500 | >500 |
| Extract Symbol | R1 | R2 | R3 | R4 | R5 |
|---|---|---|---|---|---|
| R. platyacantha part | Flowers | Leaves | Closed flowers (buds) | Leaves with stems | Flowers without petals |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabitov, A.; Gaweł-Bęben, K.; Sakipova, Z.; Strzępek-Gomółka, M.; Hoian, U.; Satbayeva, E.; Głowniak, K.; Ludwiczuk, A. Rosa platyacantha Schrenk from Kazakhstan—Natural Source of Bioactive Compounds with Cosmetic Significance. Molecules 2021, 26, 2578. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092578
Sabitov A, Gaweł-Bęben K, Sakipova Z, Strzępek-Gomółka M, Hoian U, Satbayeva E, Głowniak K, Ludwiczuk A. Rosa platyacantha Schrenk from Kazakhstan—Natural Source of Bioactive Compounds with Cosmetic Significance. Molecules. 2021; 26(9):2578. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092578
Chicago/Turabian StyleSabitov, Askhat, Katarzyna Gaweł-Bęben, Zuriyadda Sakipova, Marcelina Strzępek-Gomółka, Uliana Hoian, Elmira Satbayeva, Kazimierz Głowniak, and Agnieszka Ludwiczuk. 2021. "Rosa platyacantha Schrenk from Kazakhstan—Natural Source of Bioactive Compounds with Cosmetic Significance" Molecules 26, no. 9: 2578. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092578