
Insights into the Behavior of Triple-Negative MDA-MB-231 Breast Carcinoma Cells Following the Treatment with 17β-Ethinylestradiol and Levonorgestrel

 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
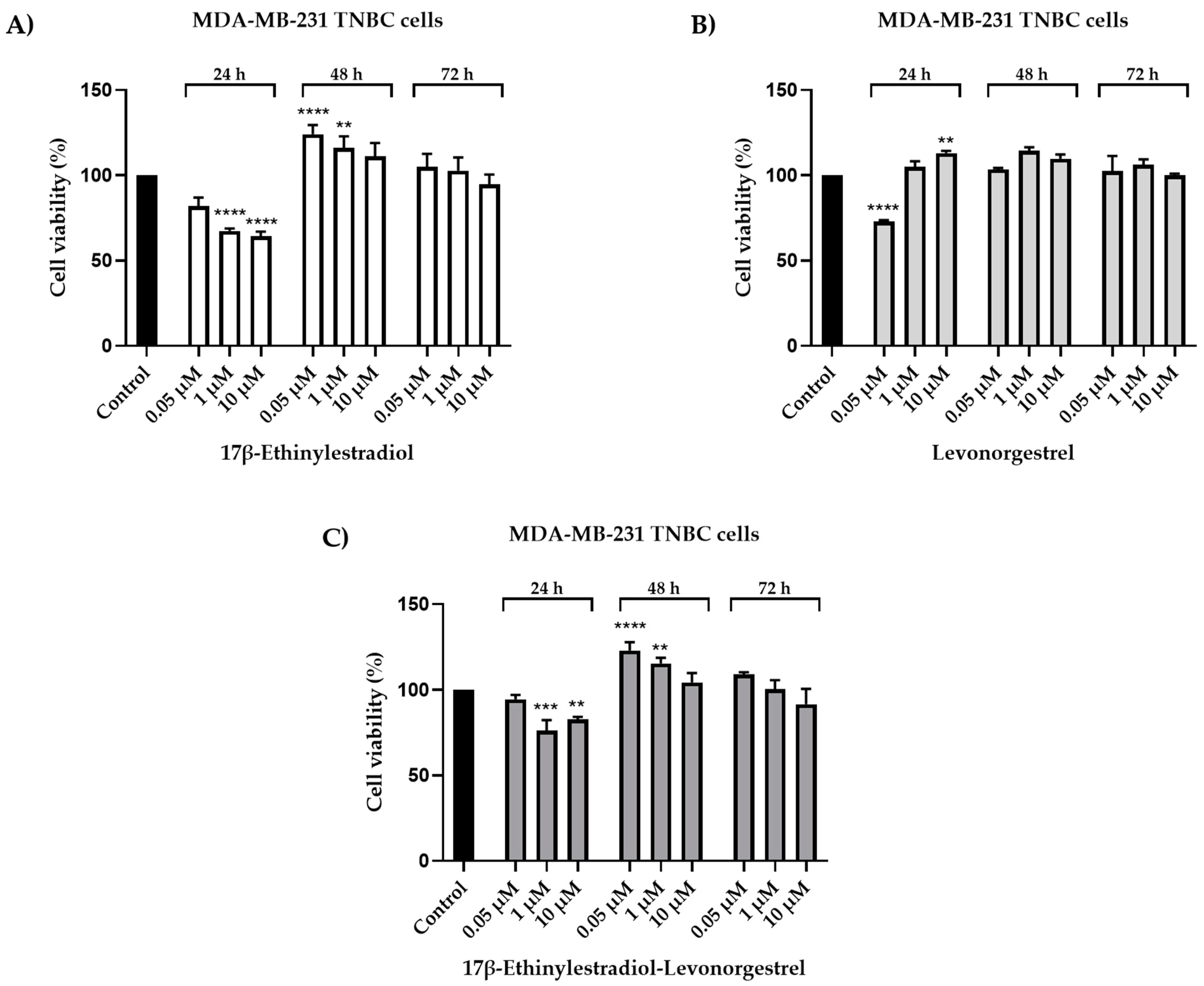
2.1. Cell Viability Evaluation
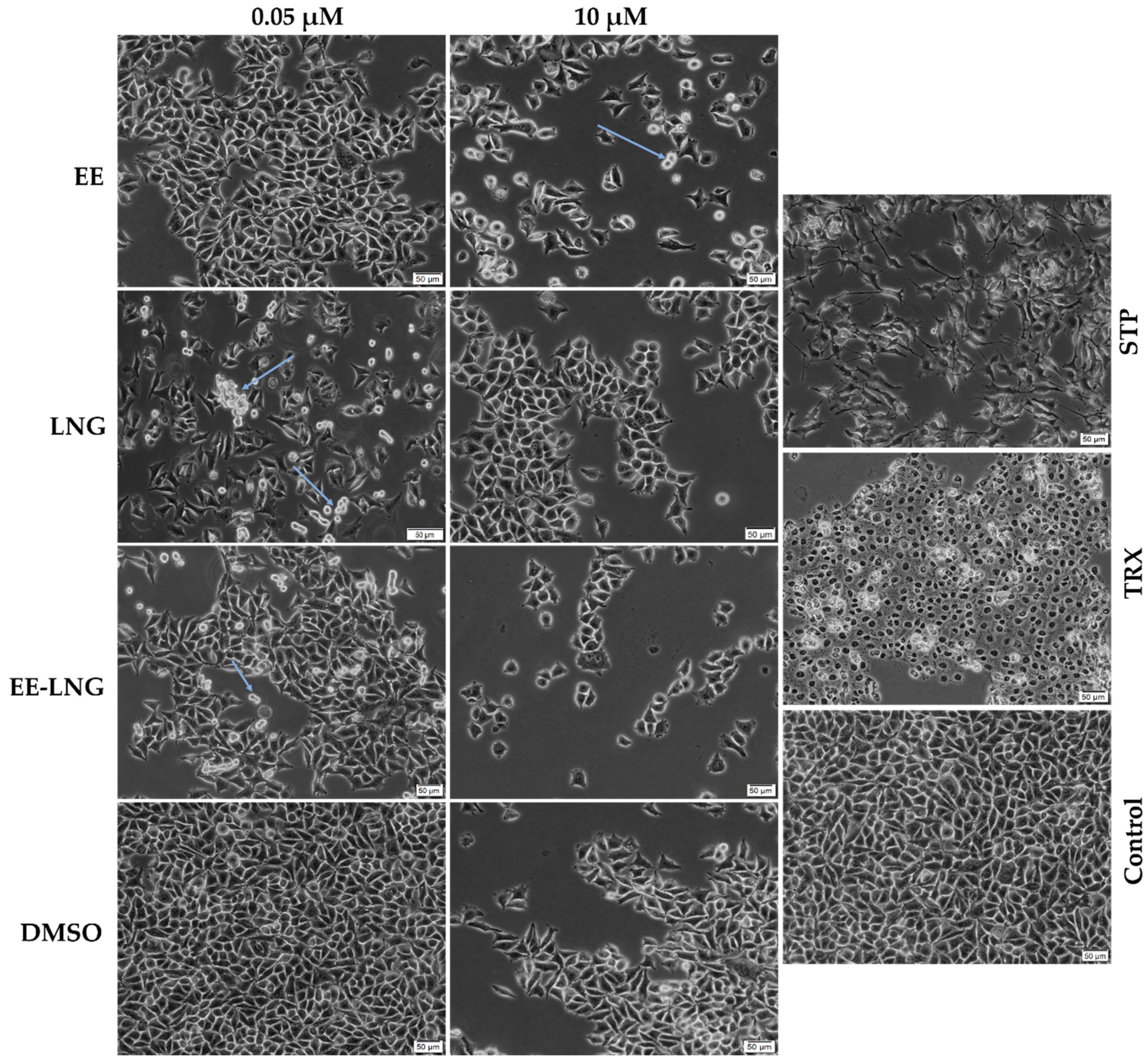
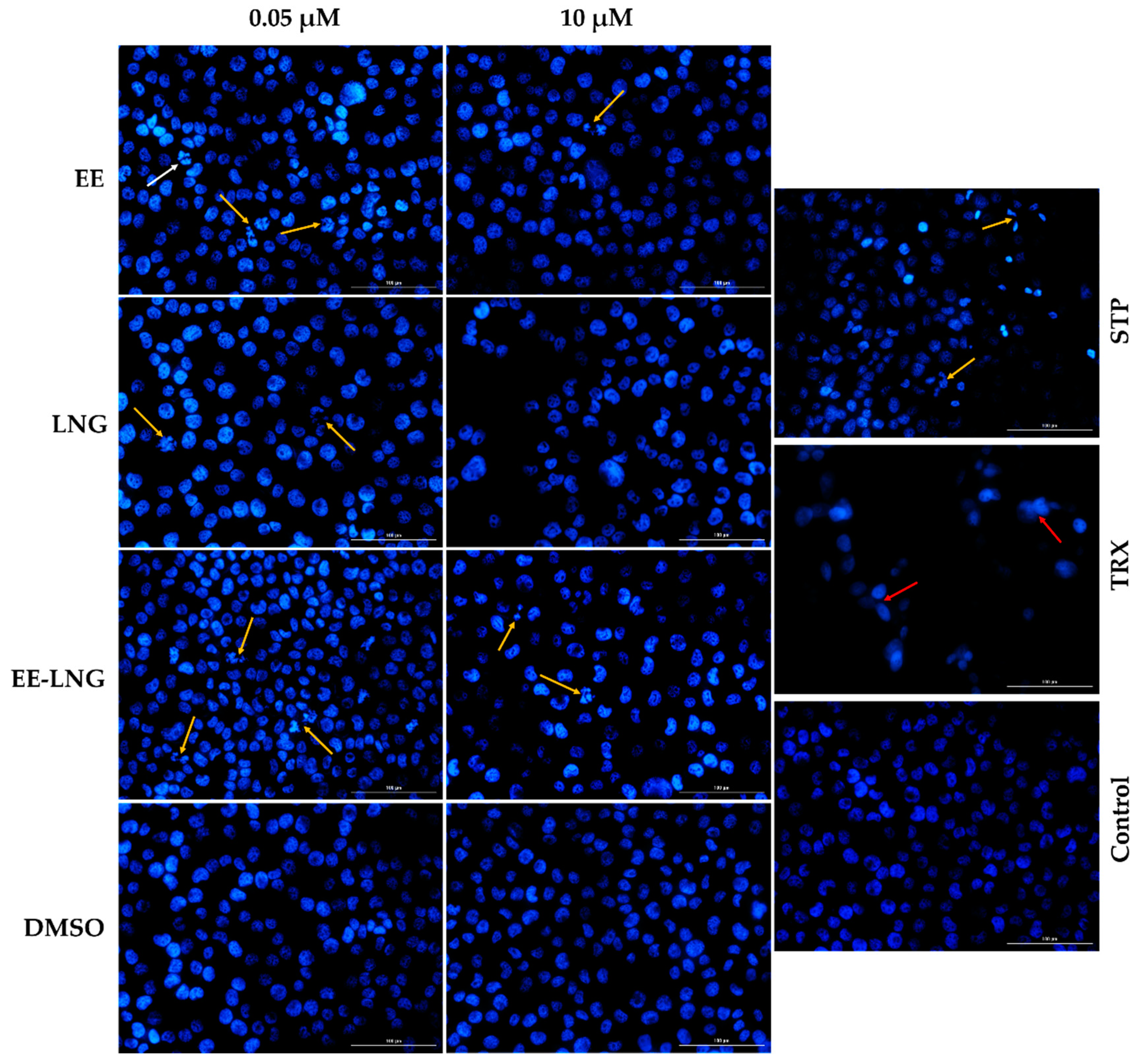
2.2. Cellular and Nuclear Morphology Assessment
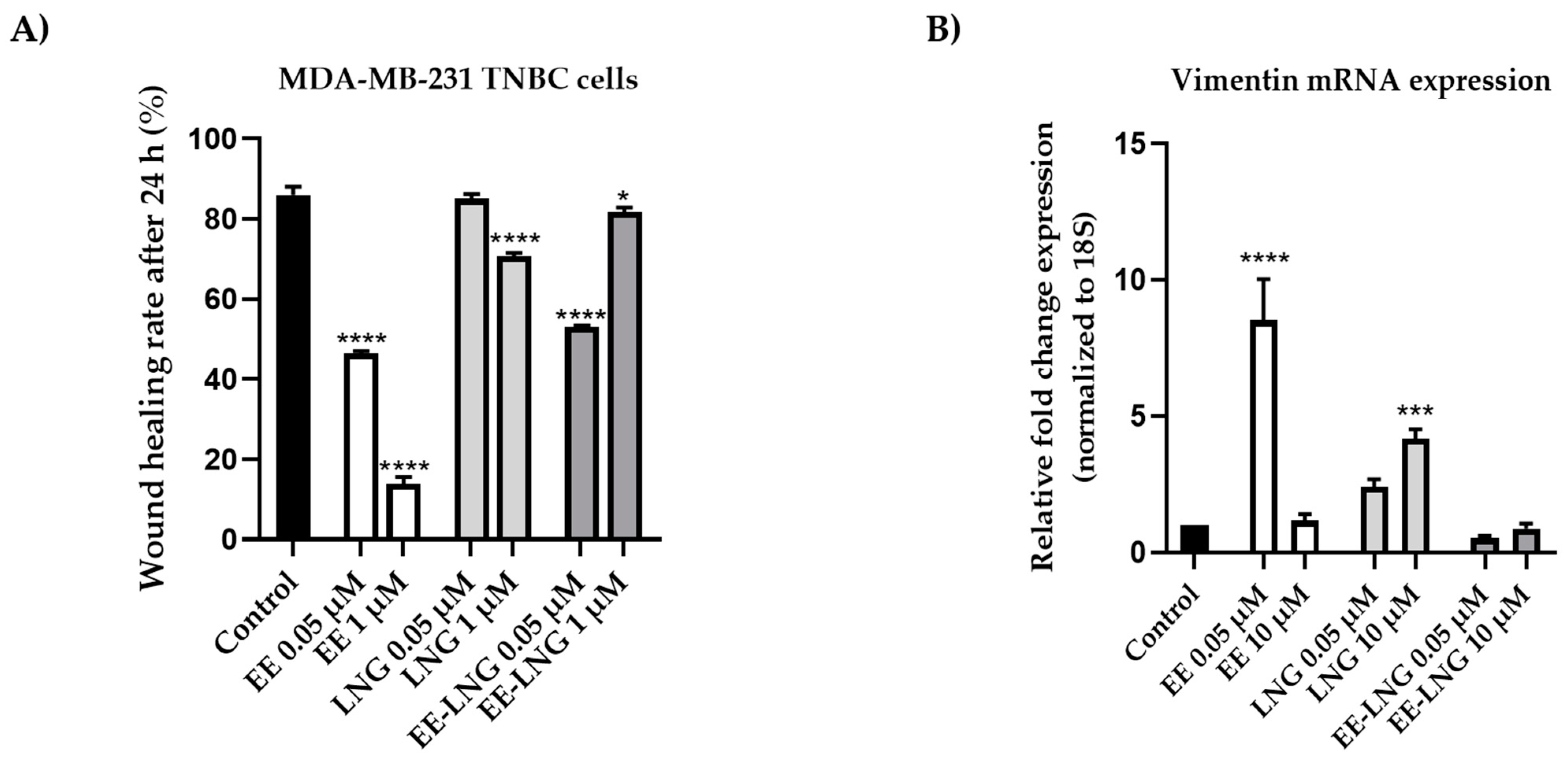
2.3. Wound Healing Assay and Quantitative Polymerase Chain Reaction (RT-qPCR)
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Line and Cell Growth Conditions
4.3. Cell Viability Evaluation
4.4. Cellular and Nuclear Morphology Assessment
4.5. Wound Healing Assay and Quantitative Polymerase Chain Reaction (RT-qPCR)
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Iversen, L.; Sivasubramaniam, S.; Lee, A.J.; Fielding, S.; Hannaford, P.C. Lifetime cancer risk and combined oral contraceptives: The Royal College of General Practitioners’ Oral Contraception Study. Am. J. Obstet. Gynecol. 2017, 216, 580.e1–580.e9. [Google Scholar] [CrossRef]
- Golobof, A.; Kiley, J. The Current Status of Oral Contraceptives: Progress and Recent Innovations. Semin. Reprod. Med. 2016, 34, 145–151. [Google Scholar] [CrossRef]
- Stanczyk, F.Z.; Archer, D.F.; Bhavnani, B.R. Ethinyl estradiol and 17β-estradiol in combined oral contraceptives: Pharmacokinetics, pharmacodynamics and risk assessment. Contraception 2013, 87, 706–727. [Google Scholar] [CrossRef] [PubMed]
- Dhont, M. History of oral contraception. Eur. J. Contracept. Reprod. Heal. Care 2010, 15. [Google Scholar] [CrossRef] [PubMed]
- Vrettakos, C.; Bajaj, T. Levonorgestrel. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- De Leo, V.; Musacchio, M.C.; Cappelli, V.; Piomboni, P.; Morgante, G. Hormonal contraceptives: Pharmacology tailored to women’s health. Hum. Reprod. Update 2016, 22, 634–646. [Google Scholar] [CrossRef] [Green Version]
- Cooper, D.B.; Mahdy, H. Oral Contraceptive Pills. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Caserta, D.; Ralli, E.; Matteucci, E.; Bordi, G.; Mallozzi, M.; Moscarini, M. Combined oral contraceptives: Health benefits beyond contraception. Panminerva Med. 2014, 56, 233–244. [Google Scholar] [PubMed]
- Sech, L.A.; Mishell, D.R. Oral steroid contraception. Womens Health 2015, 11, 743–748. [Google Scholar] [CrossRef] [Green Version]
- Coricovac, D.; Farcas, C.; Nica, C.; Pinzaru, I.; Simu, S.; Stoian, D.; Soica, C.; Proks, M.; Avram, S.; Navolan, D.; et al. Ethinylestradiol and levonorgestrel as active agents in normal skin, and pathological conditions induced by UVB exposure: In vitro and in ovo assessments. Int. J. Mol. Sci. 2018, 19, 3600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gierisch, J.M.; Coeytaux, R.R.; Urrutia, R.P.; Havrilesky, L.J.; Moorman, P.G.; Lowery, W.J.; Dinan, M.; McBroom, A.J.; Hasselblad, V.; Sanders, G.D.; et al. Oral contraceptive use and risk of breast, cervical, colorectal, and endometrial cancers: A systematic review. Cancer Epidemiol. Biomark. Prev. 2013, 22, 1931–1943. [Google Scholar] [CrossRef] [Green Version]
- Harbeck, N.; Gnant, M. Breast cancer. Lancet 2017, 389, 1134–1150. [Google Scholar] [CrossRef]
- Yip, C.H.; Rhodes, A. Estrogen and progesterone receptors in breast cancer. Future Oncol. 2014, 10, 2293–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Momenimovahed, Z.; Salehiniya, H. Epidemiological characteristics of and risk factors for breast cancer in the world. Breast Cancer (Dove Med. Press) 2019, 11, 151–164. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.N.; Schwab, R.B.; Martinez, M.E. Reproductive Risk Factors and Breast Cancer Subtypes: A Review of the Literature. Breast Cancer Res. Treat. 2014, 55, 8–30. [Google Scholar] [CrossRef]
- Şoica, C.; Voicu, M.; Ghiulai, R.; Dehelean, C.; Racoviceanu, R.; Trandafirescu, C.; Roșca, O.J.; Nistor, G.; Mioc, M.; Mioc, A. Natural Compounds in Sex Hormone-Dependent Cancers: The Role of Triterpenes as Therapeutic Agents. Front. Endocrinol. 2021, 11. [Google Scholar] [CrossRef]
- Hilton, H.N.; Clarke, C.L.; Graham, J.D. Estrogen and progesterone signalling in the normal breast and its implications for cancer development. Mol. Cell. Endocrinol. 2018, 466, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Wang, J.P.; Li, Y.; Fan, P.; Liu, G.; Zhang, N.; Conaway, M.; Wang, H.; Korach, K.S.; Bocchinfuso, W.; et al. Effects of estrogen on breast cancer development: Role of estrogen receptor independent mechanisms. Int. J. Cancer 2010, 127, 1748–1757. [Google Scholar] [CrossRef] [Green Version]
- Saha, T.; Makar, S.; Swetha, R.; Gutti, G.; Singh, S.K. Estrogen signaling: An emanating therapeutic target for breast cancer treatment. Eur. J. Med. Chem. 2019, 177, 116–143. [Google Scholar] [CrossRef]
- Settings, C.; Vingiani, A.; Pruneri, G.; Vernieri, C. Hormone Receptor Loss in Breast Cancer: Molecular and Therapeutic Implications. Cells 2020, 9, 2644. [Google Scholar]
- Treeck, O.; Schüler-Toprak, S.; Ortmann, O. Estrogen Actions in Triple-Negative Breast Cancer. Cells 2020, 9, 2358. [Google Scholar] [CrossRef]
- Khan, S.A. Progesterone Exposure and Breast Cancer Risk-Addressing Barriers. JAMA Netw. Open 2020, 3, e203608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.J.; Kurita, T.; Bulun, S.E. Progesterone action in endometrial cancer, endometriosis, uterine fibroids, and breast cancer. Endocr. Rev. 2013, 34, 130–162. [Google Scholar] [CrossRef] [Green Version]
- Yersal, O.; Barutca, S. Biological subtypes of breast cancer: Prognostic and therapeutic implications. World J. Clin. Oncol. 2014, 5, 412–424. [Google Scholar] [CrossRef]
- Lamb, C.A.; Vanzulli, S.I.; Lanari, C. Hormone receptors in breast cancer: More than estrogen receptors. Medicina 2019, 79, 540–545. [Google Scholar] [PubMed]
- Furrer, D.; Paquet, C.; Jacob, S.; Diorio, C. The Human Epidermal Growth Factor Receptor 2 (HER2) as a Prognostic and Predictive Biomarker: Molecular Insights into HER2 Activation and Diagnostic Implications. IntechOpen 2018. [Google Scholar] [CrossRef] [Green Version]
- da Silva, J.L.; Cardoso Nunes, N.C.; Izetti, P.; de Mesquita, G.G.; de Melo, A.C. Triple negative breast cancer: A thorough review of biomarkers. Crit. Rev. Oncol. Hematol. 2020, 145, 102855. [Google Scholar] [CrossRef]
- Hamilton, N.; Márquez-Garbán, D.; Mah, V.; Fernando, G.; Elshimali, Y.; Garbán, H.; Elashoff, D.; Vadgama, J.; Goodglick, L.; Pietras, R. Biologic roles of estrogen receptor- β and insulin-like growth factor-2 in triple-negative breast cancer. Biomed. Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhong, Y.; Zhang, H.; Yu, H.; Huang, Y. Association between oral contraceptive use as a risk factor and triple-negative breast cancer: A systematic review and meta-analysis. Mol. Clin. Oncol. 2017, 76–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Duan, J.J.; Bian, X.W.; Yu, S.C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 1–13. [Google Scholar] [CrossRef]
- Mehanna, J.; Haddad, F.G.H.; Eid, R.; Lambertini, M.; Kourie, H.R. Triple-negative breast cancer: Current perspective on the evolving therapeutic landscape. Int. J. Womens Health 2019, 11, 431–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowley, L.C.; Marfell, B.J.; Waterhouse, N.J. Analyzing cell death by nuclear staining with Hoechst 33342. Cold Spring Harb. Protoc. 2016, 2016, 778–781. [Google Scholar] [CrossRef]
- Sumit Bhateja, A.B. Carcinogenesis and Sex Hormones: A Review. Endocrinol. Metab. Syndr. 2015, 4, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Folkerd, E.; Dowsett, M. Sex hormones and breast cancer risk and prognosis. Breast 2013, 22, S38–S43. [Google Scholar] [CrossRef] [PubMed]
- Bardaweel, S.K.; Akour, A.A.; Al-Muhaissen, S.; Alsalamat, H.A.; Ammar, K. Oral contraceptive and breast cancer: Do benefits outweigh the risks? A case—Control study from Jordan. BMC Womens Health 2019, 19, 1–7. [Google Scholar] [CrossRef]
- Perkins, M.S.; Louw-du Toit, R.; Africander, D. A comparative characterization of estrogens used in hormone therapy via estrogen receptor (ER)-α and -β. J. Steroid Biochem. Mol. Biol. 2017, 174, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Nouriemamzaden, F.; Word, B.; Cotton, E.; Hawkins, A.; Littlejohn, K.; Moore, R.; Miranda-Carbon, G.; Orish, C.N.; Lyn-Cook, B. Modulation of estrogen α and progesterone receptors in triple negative breast cancer cell lines: The effects of vorinostat and indole-3-carbinol in vitro. Anticancer Res. 2020, 40, 3669–3683. [Google Scholar] [CrossRef]
- Zhou, L.; Zhou, W.; Zhang, H.; Hu, Y.; Yu, L.; Zhang, Y. Progesterone suppresses triple-negative breast cancer growth and metastasis to the brain via membrane progesterone receptor α. Int. J. Mol. Med. 2017, 40, 755–761. [Google Scholar] [CrossRef]
- Kores, K.; Lešnik, S.; Bren, U.; Janežič, D.; Konc, J. Discovery of Novel Potential Human Targets of Resveratrol by Inverse Molecular Docking. J. Chem. Inf. Model. 2019, 59, 2467–2478. [Google Scholar] [CrossRef]
- Kores, K.; Konc, J.; Bren, U. Mechanistic insights into side effects of troglitazone and rosiglitazone using a novel inverse molecular docking protocol. Pharmaceutics 2021, 13, 315. [Google Scholar] [CrossRef]
- Contreras-Zárate, M.J.; Day, N.L.; Ormond, D.R.; Borges, V.F.; Tobet, S.; Gril, B.; Steeg, P.S.; Cittelly, D.M. Estradiol induces BDNF/TrkB signaling in triple-negative breast cancer to promote brain metastases. Oncogene 2019, 38, 4685–4699. [Google Scholar] [CrossRef]
- Yaacob, N.S.; Nasir, R.; Norazmi, M.N. Influence of 17β-Estradiol on 15-Deoxy-Δ 12, 14 Prostaglandin J 2 -Induced Apoptosis in MCF-7 and MDA-MB-231 Cells. Asian Pac. J. Cancer Prev. 2013, 14, 6761–6767. [Google Scholar] [CrossRef] [Green Version]
- Press, D. Safety, efficacy and patient satisfaction with continuous daily administration of levonorgestrel / ethinylestradiol oral contraceptives. Patient Prefer. Adherence 2009, 3, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Kathryn, J.C.; Sireesha, V.G.; Stanley, L. Triple Negative Breast Cancer Cell Lines: One Tool in the Search for Better Treatment of Triple Negative Breast Cancer. Breast Dis. 2012, 32, 35–48. [Google Scholar] [CrossRef]
- Welsh, J.E. Animal Models for Studying Prevention and Treatment of Breast Cancer; Academic Press: Cambridge, MA, USA, 2013; pp. 997–1018. [Google Scholar] [CrossRef]
- Holliday, D.L.; Speirs, V. Choosing correct breast cancer cell line. Breast Cancer Res. 2011, 13, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, M.; Browne, E.; Guarinoni, L.; Darveau, T.; Hilton, K.; Witt-Enderby, P.A. Novel Melatonin, Estrogen, and Progesterone Hormone Therapy Demonstrates Anti-Cancer Actions in MCF-7 and MDA-MB-231 Breast Cancer Cells. Breast Cancer Basic Clin. Res. 2020, 14. [Google Scholar] [CrossRef] [PubMed]
- Coelingh Bennink, H.J.T.; Verhoeven, C.; Dutman, A.E.; Thijssen, J. The use of high-dose estrogens for the treatment of breast cancer. Maturitas 2017, 95, 11–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinsche, O.; Girgert, R.; Emons, G.; Gründker, C. Estrogen receptor β selective agonists reduce invasiveness of triple-negative breast cancer Cells. Int. J. Oncol. 2015, 46, 878–884. [Google Scholar] [CrossRef] [Green Version]
- Vladusic, E.A.; Hornby, A.E.; Guerra-Vladusic, F.K.; Lakins, J.; Lupu, R. Expression and regulation of estrogen receptor ß in human breast tumors and cell lines. Oncol. Rep. 2000, 7, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.M.; Yang, M.F.; Yu, W.; Tao, H.M. Molecular mechanisms of estrogen receptor β-induced apoptosis and autophagy in tumors: Implication for treating osteosarcoma. J. Int. Med. Res. 2019, 47, 4644–4655. [Google Scholar] [CrossRef] [Green Version]
- Karandeep, S.; Lauren, A. Breast and ovarian cell behavior in the presence of contraceptive ingredients. J. Appl. Biotechnol. Bioeng. 2020, 7, 67–80. [Google Scholar] [CrossRef]
- Santen, R.J.; Yue, W.; Wang, J.P. Estrogen metabolites and breast cancer. Steroids 2015, 99, 61–66. [Google Scholar] [CrossRef]
- Wen, C.; Wu, L.; Fu, L.; Wang, B.; Zhou, H. Unifying mechanism in the initiation of breast cancer by metabolism of estrogen (Review). Mol. Med. Rep. 2017, 16, 1001–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merki-Feld, G.S.; Seeger, H.; Mueck, A.O. Proliferative effects of estradiol- or ethinylestradiol-progestogen combinations on human breast cancer cells in an intermitted and a long-term regimen. Horm. Metab. Res. 2012, 44, 415–421. [Google Scholar] [CrossRef]
- Kuhl, H.; Schneider, H.P.G. Progesterone—Promoter or inhibitor of breast cancer. Climacteric 2013, 16, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Lemus, A.E. Mechanism of action of levonorgestrel: In vitro metabolism and specific interactions with steroid receptors in target organs. J. Steroid Biochem. Mol. Biol. 1992, 41, 881–890. [Google Scholar] [CrossRef]
- Wiebe, J.P.; Lewis, M.J. Activity and expression of progesterone metabolizing 5α-reductase, 20α-hydroxysteroid oxidoreductase and 3α(β)-hydroxysteroid oxidoreductases in tumorigenic (MCF-7, MDA-MB-231, T-47D) and nontumorigenic (MCF-10A) human breast cancer Cells. BMC Cancer 2003, 3, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, V.C. The New Biology of Estrogen-induced Apoptosis Applied to Treat and Prevent Breast Cancer. Physiol. Behav. 2016, 176, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.S.; Meeke, K.; Osipo, C.; Ross, E.A.; Kidawi, N.; Li, T.; Bell, E.; Chandel, N.S.; Jordan, V.C. Intrinsic mechanism of estradiol-induced apoptosis in breast cancer cells resistant to estrogen deprivation. J. Natl. Cancer Inst. 2005, 97, 1746–1759. [Google Scholar] [CrossRef] [PubMed]
- McGlorthan, L.; Paucarmayta, A.; Casablanca, Y.; Maxwell, G.L.; Syed, V. Progesterone induces apoptosis by activation of caspase-8 and calcitriol via activation of caspase-9 pathways in ovarian and endometrial cancer cells in vitro. Apoptosis 2021, 26, 184–194. [Google Scholar] [CrossRef]
- Zhao, X.; Tang, X.; Ma, T.; Ding, M.; Bian, L.; Chen, D.; Li, Y.; Wang, L.; Zhuang, Y.; Xie, M.; et al. Levonorgestrel Inhibits Human Endometrial Cell Proliferation through the Upregulation of Gap Junctional Intercellular Communication via the Nuclear Translocation of Ser255 Phosphorylated Cx43. Biomed. Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Wusu, A.D.; Bankole, H.A.; Fatai, A.A.; Obasieke, P.E.; Wusu, T.D.; Afolabi, O.K. Combined Oral Administration of Ethinylestradiol and Levonorgestrel Alters the Expression of Antioxidant and Apoptotic Markers in Female Rats. Sci. World J. 2021, 16, 57–63. [Google Scholar]
- Zhou, K.; Sun, P.; Zhang, Y.; You, X.; Li, P.; Wang, T. Estrogen stimulated migration and invasion of estrogen receptor-negative breast cancer cells involves an ezrin-dependent crosstalk between G protein-coupled receptor 30 and estrogen receptor beta signaling. Steroids 2016, 111, 113–120. [Google Scholar] [CrossRef]
- Liu, C.Y.; Lin, H.H.; Tang, M.J.; Wang, Y.K. Vimentin contributes to epithelial-mesenchymal transition ancer cell mechanics by mediating cytoskeletal organization and focal adhesion maturation. Oncotarget 2015, 6, 15966–15983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun-Ling Lin, V.; Jin, R.; Tan, P.H.; Aw, S.E.; Woon, C.T.; Bay, B.H. Progesterone induces cellular differentiation in MDA-MB-231 breast cancer cells transfected with progesterone receptor complementary DNA. Am. J. Pathol. 2003, 162, 1781–1787. [Google Scholar] [CrossRef] [Green Version]
- Felice, F.; Zambito, Y.; Belardinelli, E.; Fabiano, A.; Santoni, T.; Di Stefano, R. Effect of different chitosan derivatives on in vitro scratch wound assay: A comparative study. Int. J. Biol. Macromol. 2015, 76, 236–241. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simu, S.; Marcovici, I.; Dobrescu, A.; Malita, D.; Dehelean, C.A.; Coricovac, D.; Olaru, F.; Draghici, G.A.; Navolan, D. Insights into the Behavior of Triple-Negative MDA-MB-231 Breast Carcinoma Cells Following the Treatment with 17β-Ethinylestradiol and Levonorgestrel. Molecules 2021, 26, 2776. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092776
Simu S, Marcovici I, Dobrescu A, Malita D, Dehelean CA, Coricovac D, Olaru F, Draghici GA, Navolan D. Insights into the Behavior of Triple-Negative MDA-MB-231 Breast Carcinoma Cells Following the Treatment with 17β-Ethinylestradiol and Levonorgestrel. Molecules. 2021; 26(9):2776. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092776
Chicago/Turabian StyleSimu, Sebastian, Iasmina Marcovici, Amadeus Dobrescu, Daniel Malita, Cristina Adriana Dehelean, Dorina Coricovac, Flavius Olaru, George Andrei Draghici, and Dan Navolan. 2021. "Insights into the Behavior of Triple-Negative MDA-MB-231 Breast Carcinoma Cells Following the Treatment with 17β-Ethinylestradiol and Levonorgestrel" Molecules 26, no. 9: 2776. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092776