
Interactive Effect of Biological Agents Chitosan, Lentinan and Ningnanmycin on Papaya Ringspot Virus Resistance in Papaya (Carica papaya L.)
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Materials and Instruments
2.2. Experiment Design
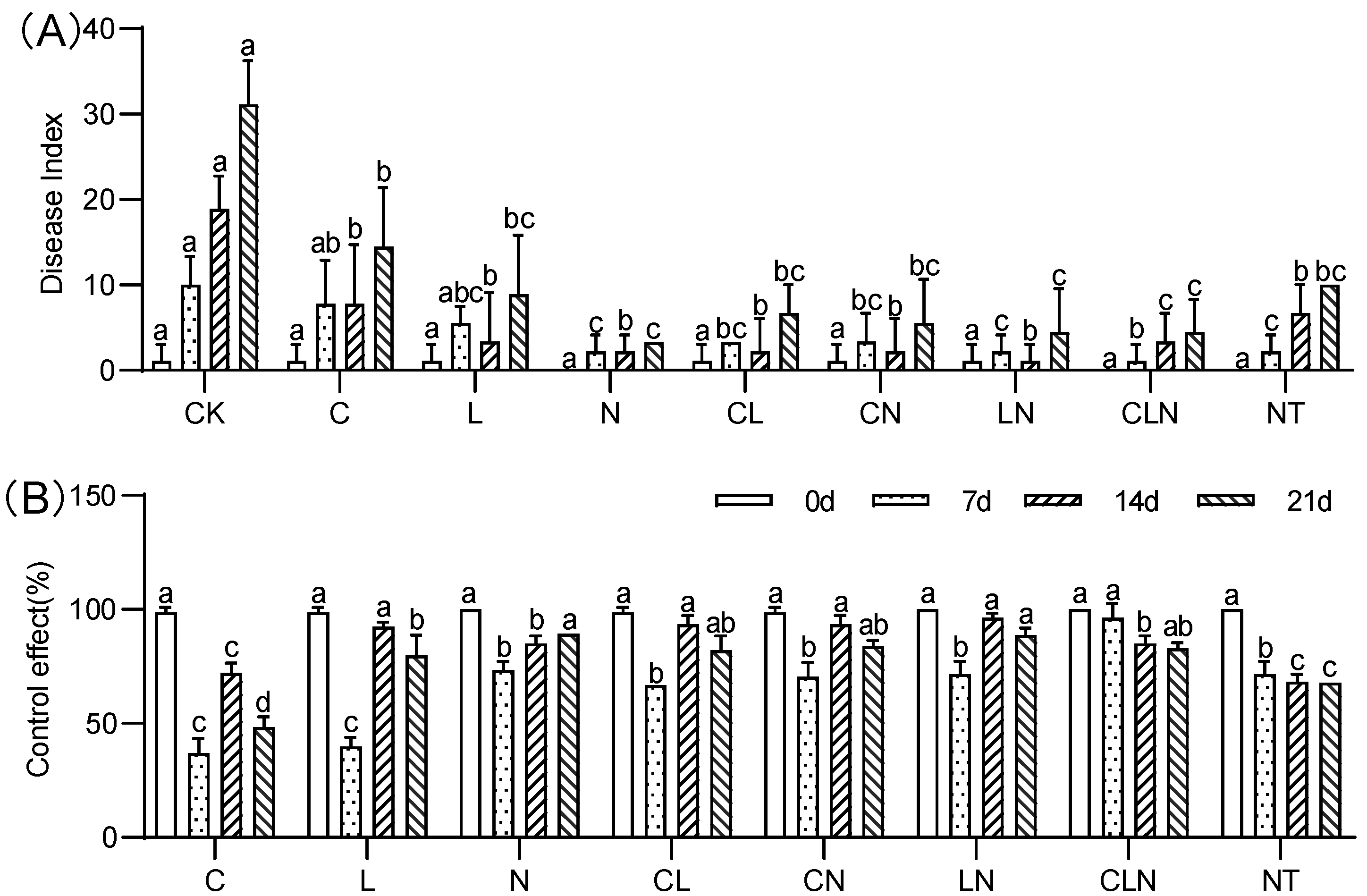
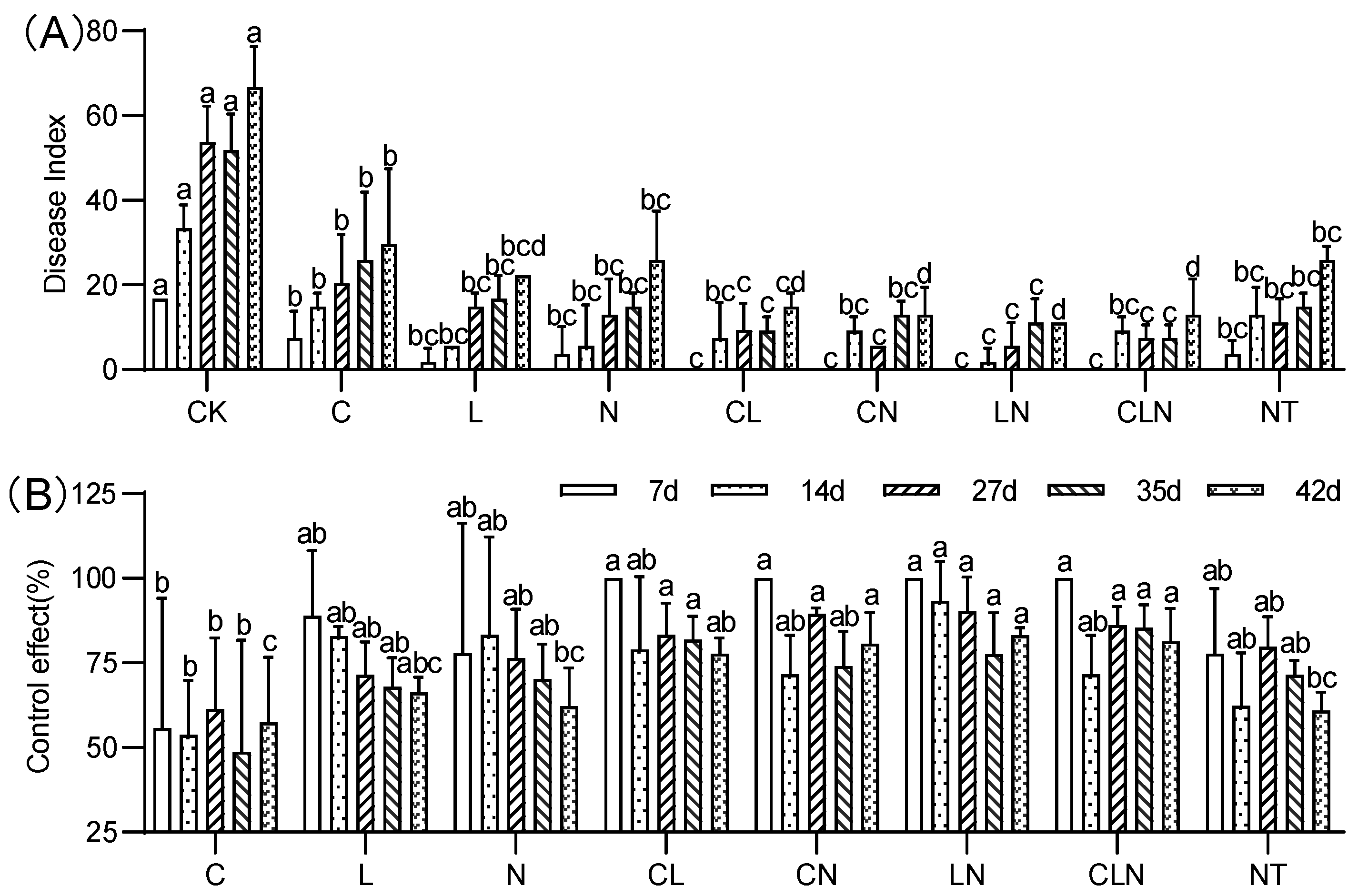
2.3. Investigation of Disease Index and Control Effect
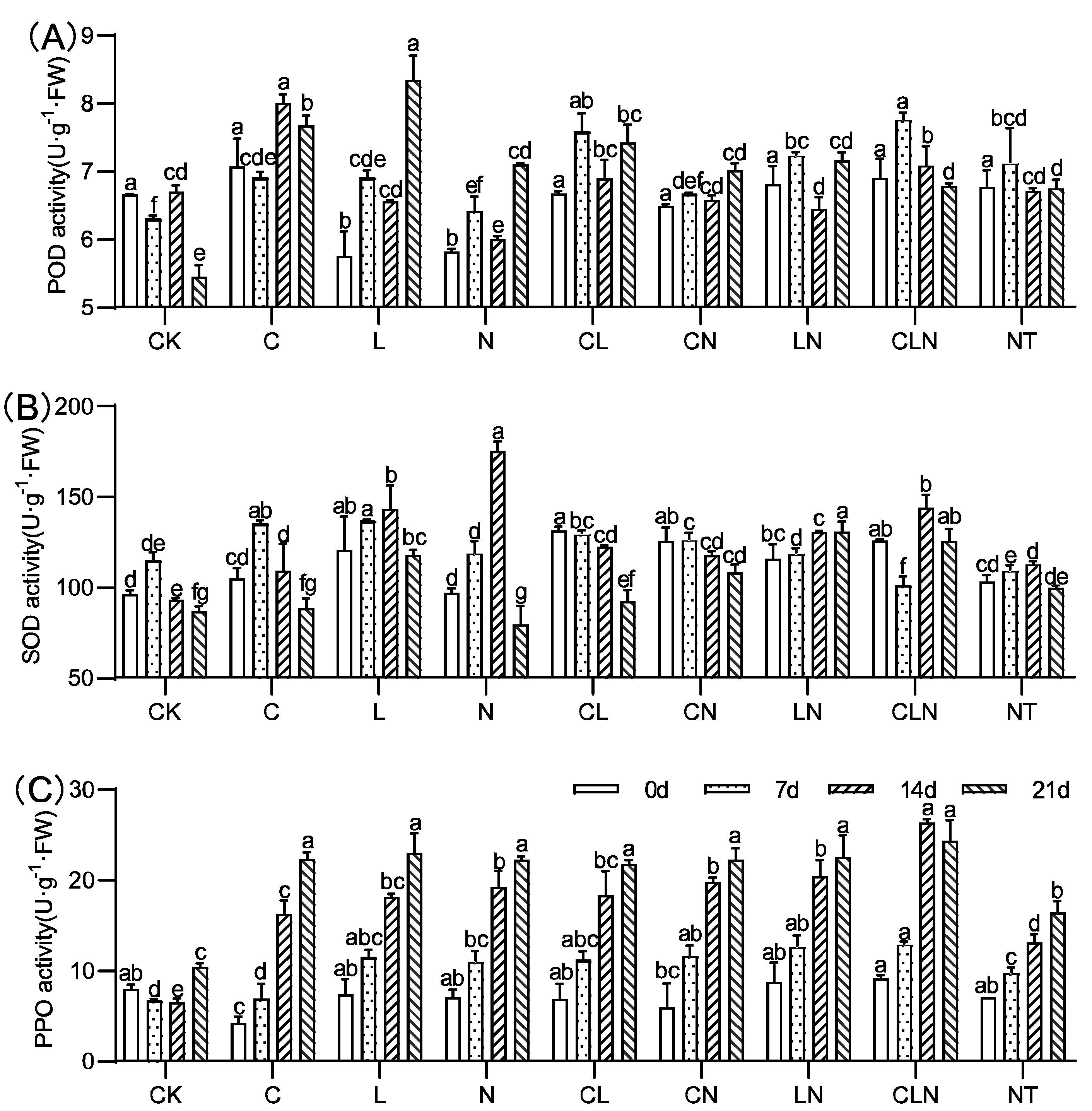
2.4. Determination of Related Physiological Indexes in Papaya
2.5. RNA-Seq Library Preparation and Sequencing
2.6. RNA-Seq Data Analysis
2.7. The qPCR Fluorescence Quantitative Test Method
2.8. Statistical Analysis of Data
3. Results
3.1. Effects of Chitosan, Lentinan and Ningnanmycin on PRSD during Seedling Stage
3.1.1. PRSD Disease Index and Control Effect during Seedling Stage
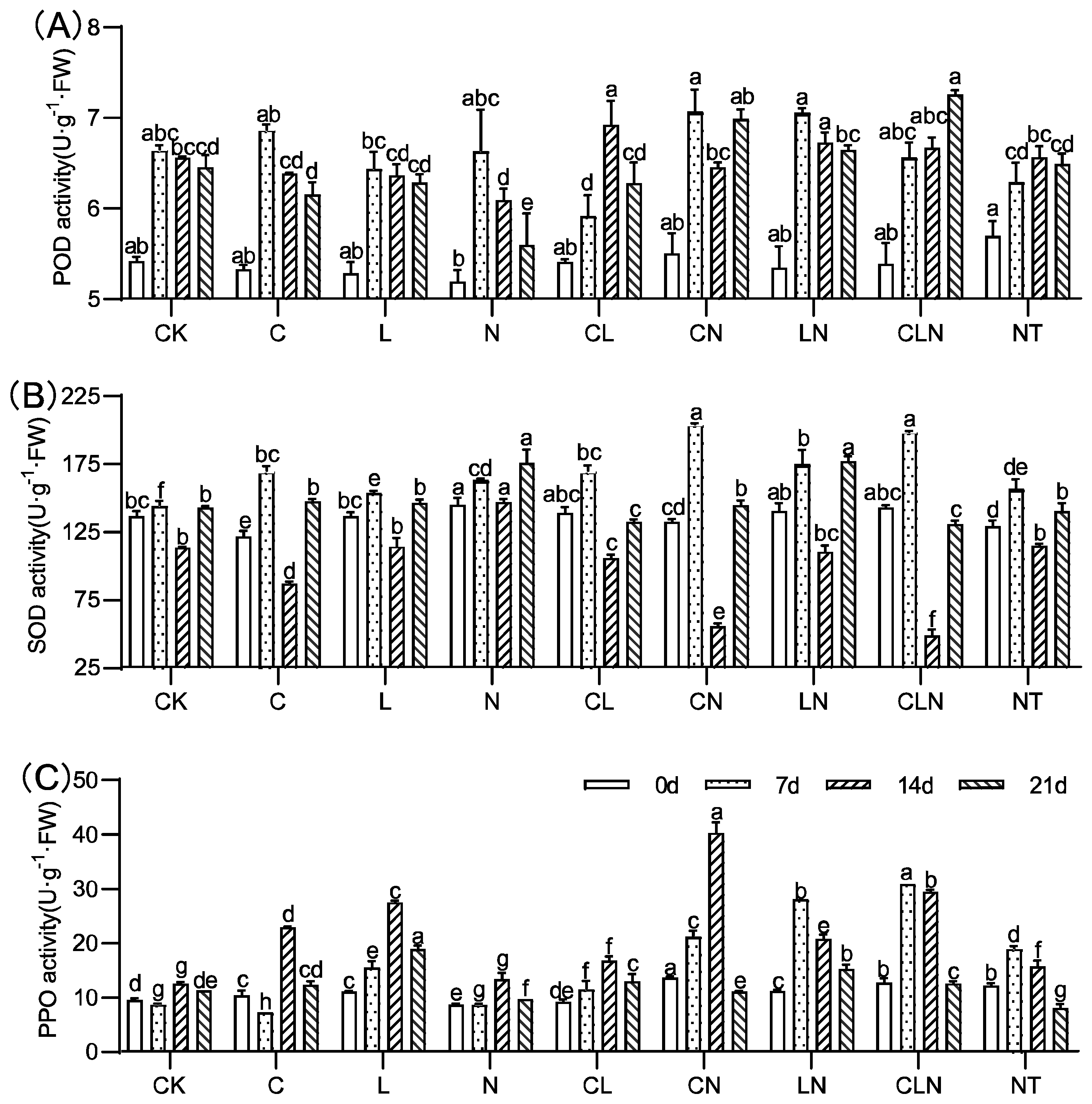
3.1.2. Changes of POD, SOD and PPO in Seedling Stage in Papaya
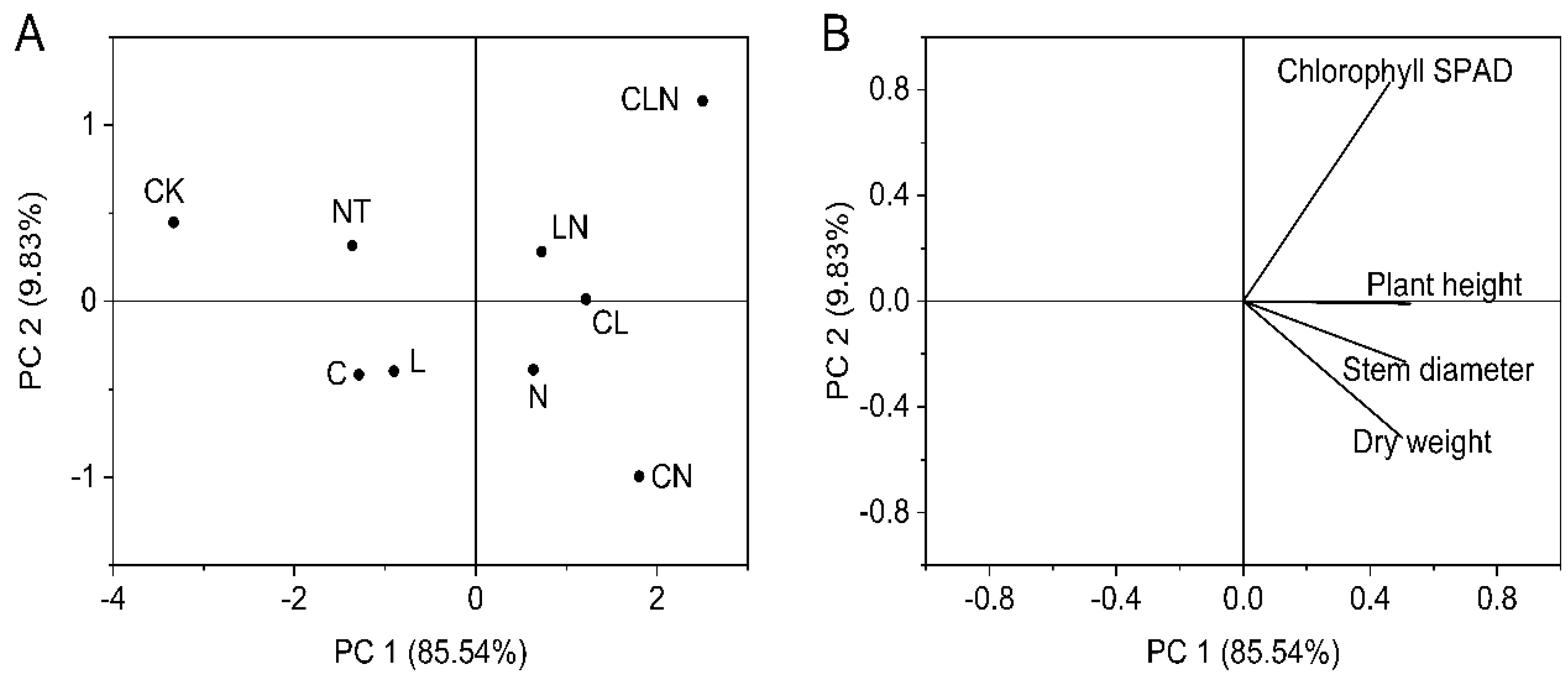
3.1.3. Growth of Stem and Leaf in Seedling Stage in Papaya
3.2. Effects of Chitosan, Lentinan and Ningnanmycin on PRSV during Fruiting Period
3.2.1. PRSD Disease Index and Control Effect during Fruiting Period
3.2.2. Changes of POD, SOD and PPO in Fruiting Period in Papaya
3.2.3. Growth of Stem, Leaf and Fruit in Fruiting Stage
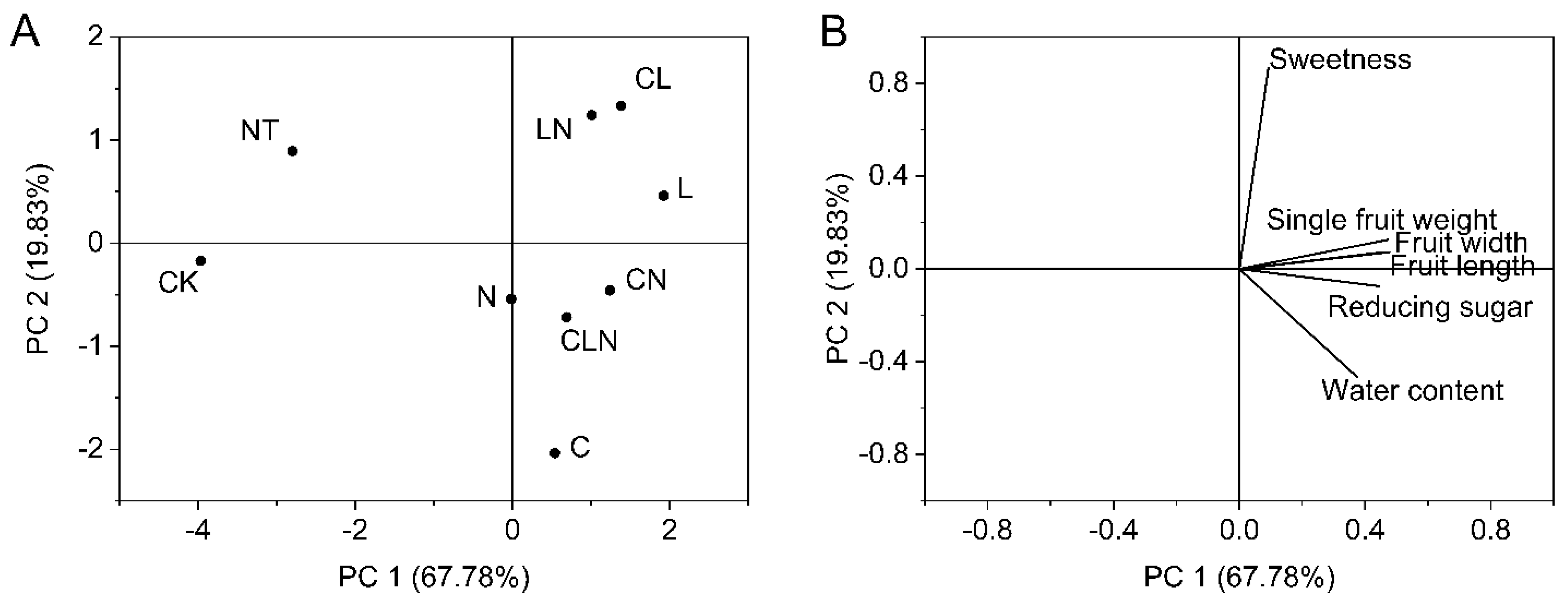
3.2.4. Growth Analysis of Fruit in Papaya
3.3. Transcriptome Analysis of Leaves of Papaya
3.3.1. RNA Quality Assessment
3.3.2. Illumina High-Throughput Sequencing and Comparison
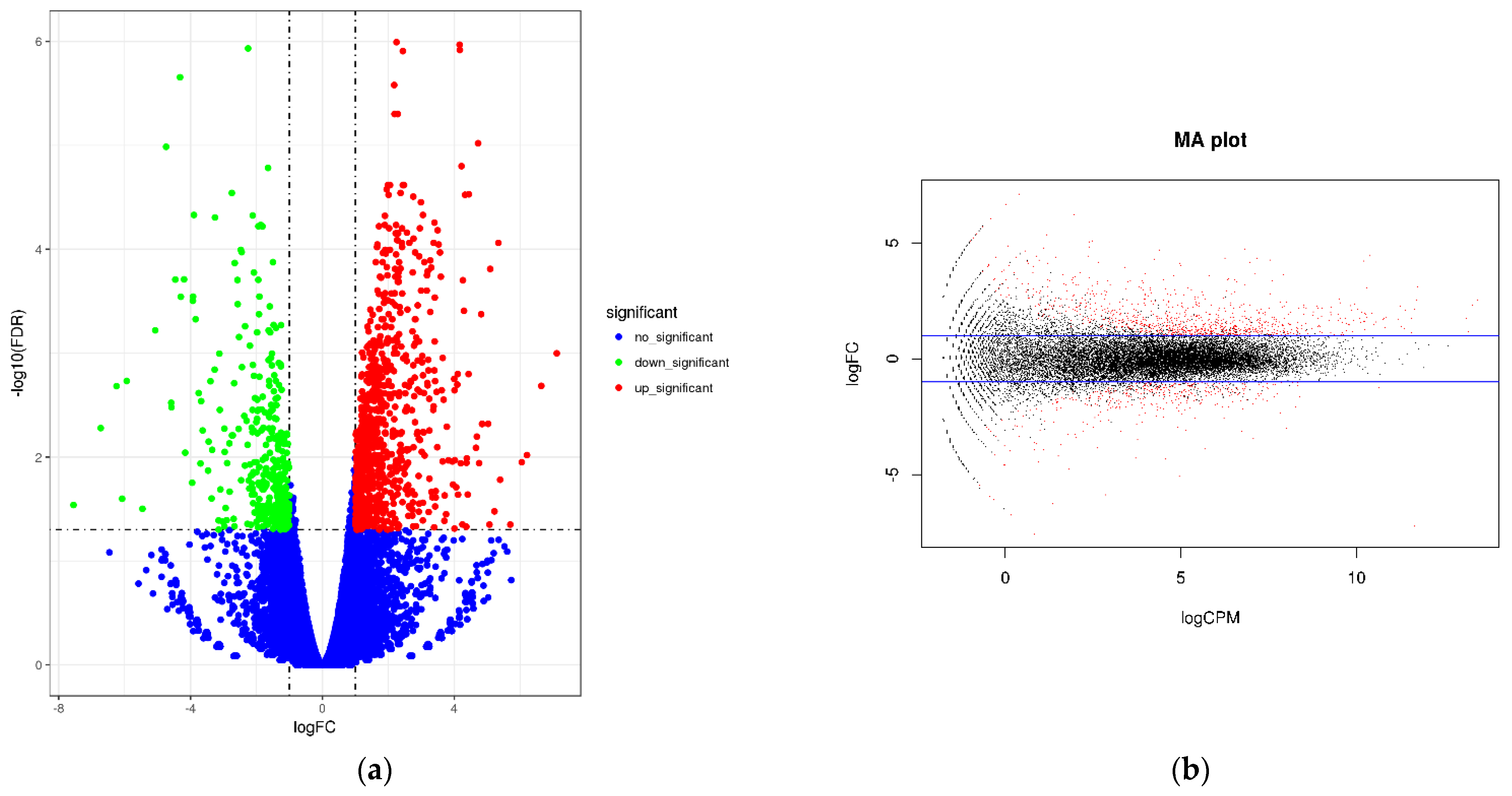
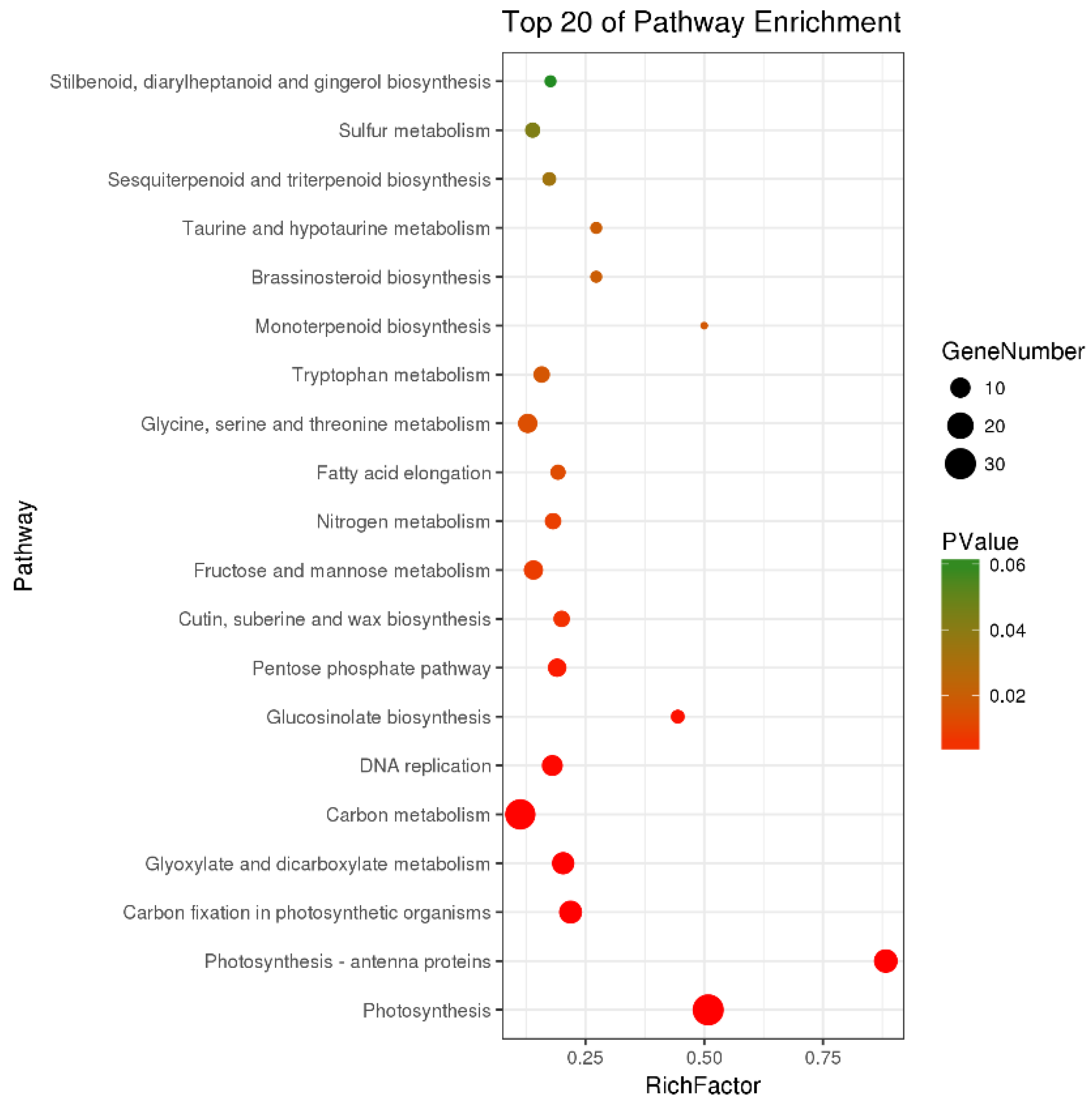
3.3.3. Analysis of 22 DEGs
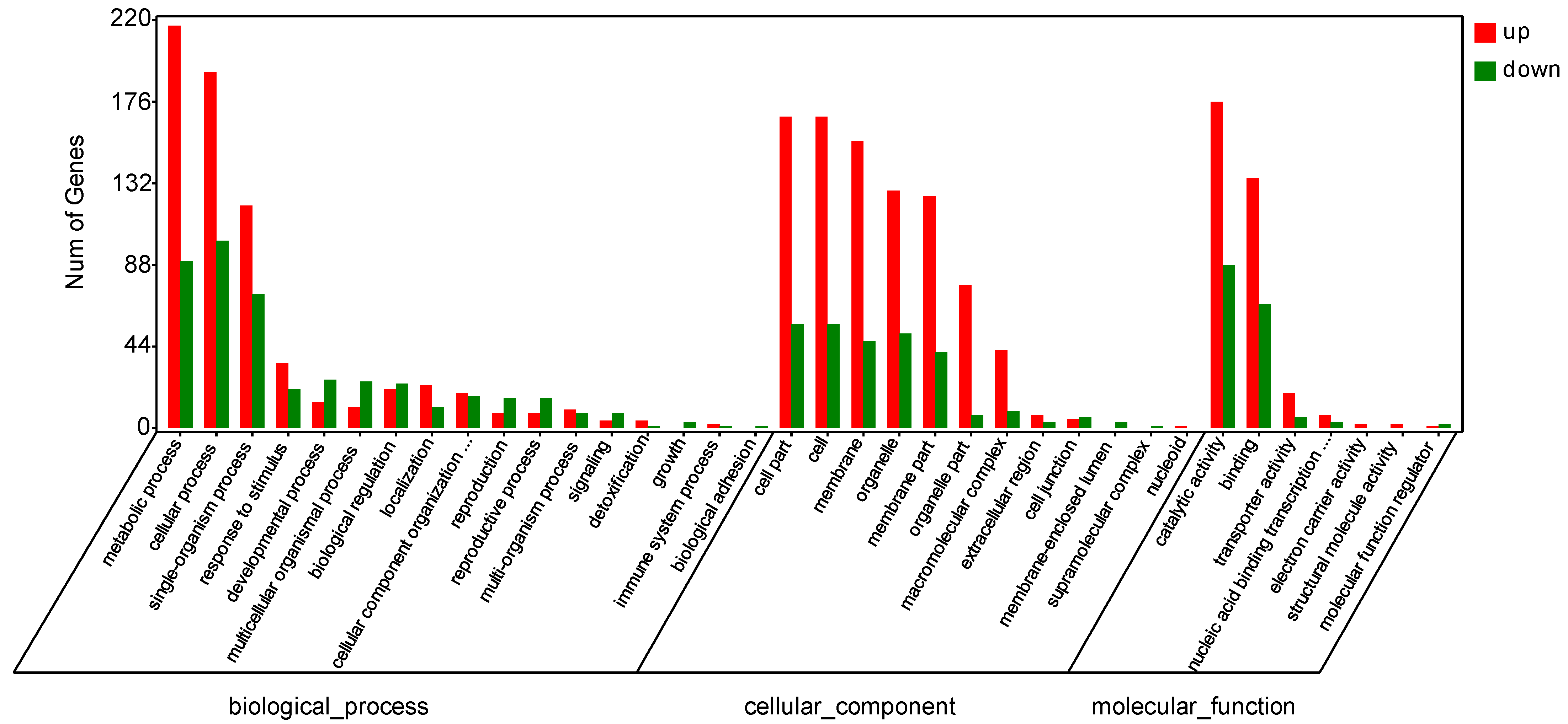
3.3.4. GO Analysis of DEGs
3.3.5. The qPCR Verification of 7 Genes
4. Discussion
4.1. Control Effects of Chitosan, Lentinan and Ningnanmycin on PRSV in Papaya
4.2. Defense Enzymes (POD, SOD, PPO) Regulating of Chitosan, Lentinan and Ningnanmycin in Papaya
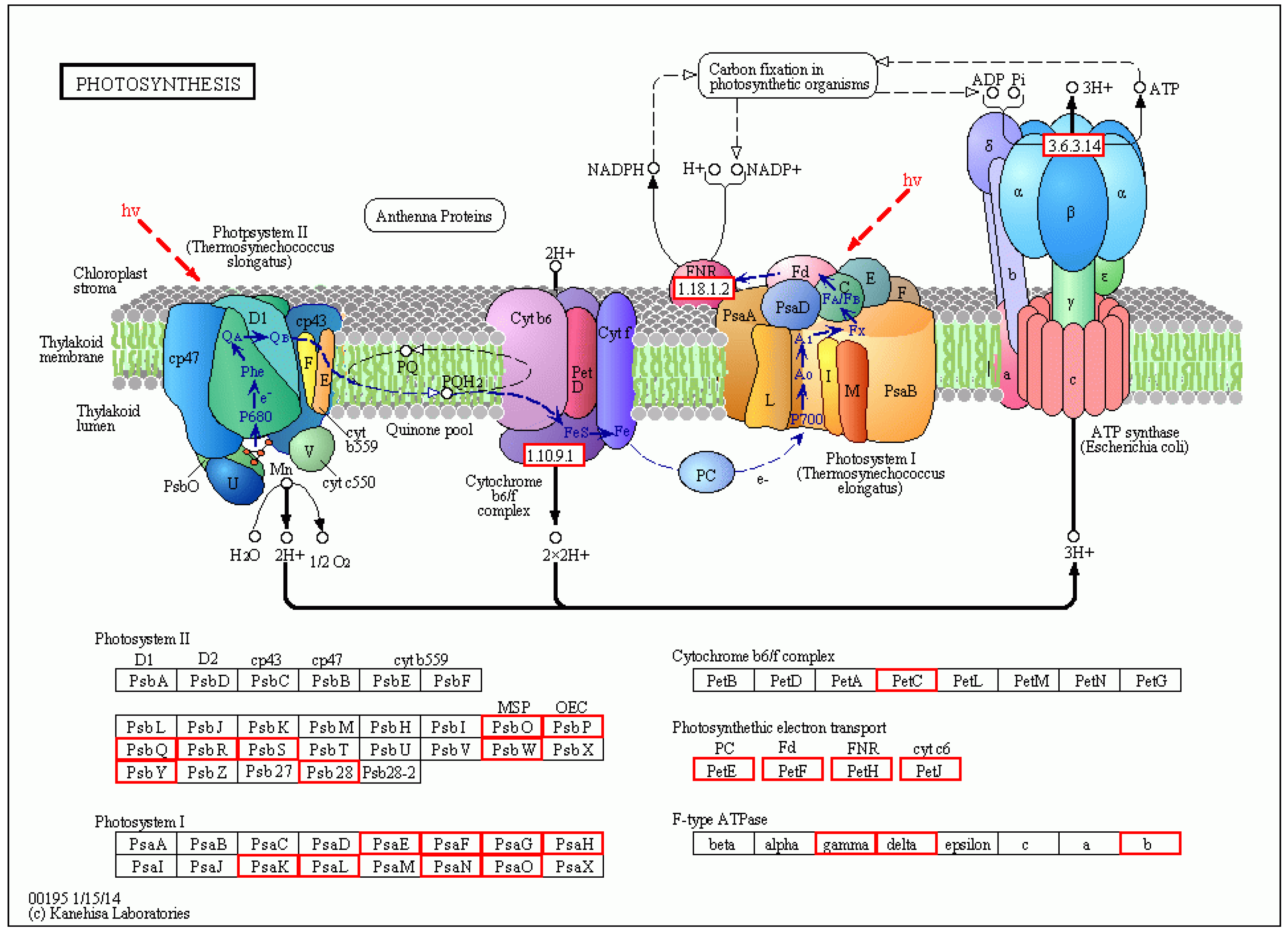
4.3. Molecular Mechanism of Resistance to PRSV Induced by Bio-Pesticides in Papaya
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Evans, E.A.; Ballen, F.H. An Overview of Global Papaya Production, Trade, and Consumption. Food Resour. Econ. 2015. Available online: http://edis.ifas.ufl.edu (accessed on 1 August 2022). [CrossRef]
- FAO. Crop Data; Food and Agriculture Organization of the United Nations (FAO): Roma, Italy, 2018; Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 28 February 2022).
- Zhao, G.; Yan, P.; Shen, W.; Tuo, D.; Li, X.; Zhou, P. Complete Genome Sequence of Papaya Ringspot Virus Isolated from Genetically Modified Papaya in Hainan Island, China. Genome Announc. 2015, 3, e01056-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, S.-D.; Jan, F.-J.; Chiang, C.-H.; Doong, T.-J.; Chen, M.-C.; Chung, P.-H.; Bau, H.-J. Complete Nucleotide Sequence and Genetic Organization of Papaya Ringspot Virus RNA. J. Gen. Virol. 1992, 73, 2531–2541. [Google Scholar] [CrossRef] [PubMed]
- Iriti, M.; Varoni, E.M. Chitosan-induced antiviral activity and innate immunity in plants. Environ. Sci. Pollut. Res. 2015, 22, 2935–2944. [Google Scholar] [CrossRef]
- Vitti, A.; La Monaca, E.; Sofo, A.; Scopa, A.; Cuypers, A.; Nuzzaci, M. Beneficial effects of Trichoderma harzianum T-22 in tomato seedlings infected by Cucumber mosaic virus (CMV). BioControl 2014, 60, 135–147. [Google Scholar] [CrossRef]
- An, N.; Lv, J.H.; Zhang, A.N.; Xiao, C.; Zhang, R.P.; Chen, P. Gene expression profiling of papaya (Carica papaya L.) immune response induced by CTS-N after inoculating PLDMV. Gene 2020, 755, 144845. [Google Scholar] [CrossRef]
- Yan, X.X.; Li, P.; Chen, P.; Fan, J.H.; Liu, R.D.; Zhang, R.P. Study on the effect and physiological basis of four biopesticides against papaya Mosaic virus disease. South China Fruits 2019, 48, 75–81. [Google Scholar]
- Xing, K.; Zhu, X.; Peng, X.; Qin, S. Chitosan antimicrobial and eliciting properties for pest control in agriculture: A review. Agron. Sustain. Dev. 2015, 35, 569–588. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.C.; Meng, Q.S.; Zeng, H.H.; Wang, W.X.; Yin, H. Chitosan oligosaccharide induces resistance to Tobacco mosaic virus in Arabidopsis via the salicylic acid-mediated signalling pathway. Sci. Rep. 2016, 6, 26144. [Google Scholar] [CrossRef] [Green Version]
- Nagorskaya, V.; Reunov, A.; Lapshina, L.; Davydova, V.; Yermak, I. Effect of chitosan on tobacco mosaic virus (TMV) accumulation, hydrolase activity, and morphological abnormalities of the viral particles in leaves of N. tabacum L. cv. Samsun. Virol. Sin. 2014, 29, 250–256. [Google Scholar] [CrossRef]
- Kumaraswamy, R.V.; Kumari, S.; Choudhary, R.C.; Pal, A.; Raliya, R.; Biswas, P.; Saharan, V. Engineered chitosan based nanomaterials: Bioactivities, mechanisms and perspectives in plant protection and growth. Int. J. Biol. Macromol. 2018, 113, 494–506. [Google Scholar] [CrossRef]
- Wang, J.; Wang, H.-Y.; Xia, X.-M.; Li, P.-P.; Wang, K.-Y. Inhibitory effect of sulfated lentinan and lentinan against tobacco mosaic virus (TMV) in tobacco seedlings. Int. J. Biol. Macromol. 2013, 61, 264–269. [Google Scholar] [CrossRef]
- Gao, D.; Wang, D.M.; Chen, K.; Huang, M.X.; Xie, X.; Li, X.Y. Activation of biochemical factors in CMV-infected tobacco by ningnanmycin. Pestic. Biochem. Physiol. 2019, 156, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.G.; Luo, Y.; Qin, S.R.; Xi, L.; Wan, B.; Du, L.F. Induction of systemic resistance against tobacco mosaic virus by Ningnanmycin in tobacco. Pestic. Biochem. Physiol. 2014, 111, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.Z.; Lv, Y.D.; Zhao, T.M.; Li, N.; Yang, Y.W.; Yu, W.G.; He, X.; Liu, T.L.; Zhang, B.L. Comparative Transcriptome Profiling of a Resistant vs. Susceptible Tomato (Solanum lycopersicum) Cultivar in Response to Infection by Tomato Yellow Leaf Curl Virus. PLoS ONE 2013, 8, e80816. [Google Scholar] [CrossRef] [PubMed]
- Luan, H.Y.; Zang, H.; Shen, H.Q.; Zhang, Y.H.; Qiao, H.L.; Tao, H.; Shen, Y.X.; Chen, H. Transcriptome Analysis of Albino Lemma Mutant in Barley. J. Nucl. Agric. Sci. 2017, 31, 2332–2339. [Google Scholar]
- Alazem, M.; Lin, N.-S. Interplay between ABA signaling and RNA silencing in plant viral resistance. Curr. Opin. Virol. 2020, 42, 1–7. [Google Scholar] [CrossRef]
- Alazem, M.; Lin, N.-S. Roles of plant hormones in the regulation of host–virus interactions. Mol. Plant Pathol. 2015, 16, 529–540. [Google Scholar] [CrossRef]
- Lu, H.Y. Monitoring and Transcriptome Analysis of the Infection of Sweet Potato Virus Disease (SPVD) in Sweet Potato. PhD Dissertation, Southwest University, Chongqing, China, 2016. [Google Scholar]
- Ming, R.; Hou, S.; Feng, Y.; Yu, Q.; Dionne-Laporte, A.; Saw, J.H.; Senin, P.; Wang, W.; Ly, B.V.; Lewis, K.L.T.; et al. The draft genome of the transgenic tropical fruit tree papaya (Carica papaya Linnaeus). Nature 2008, 452, 991–996. [Google Scholar] [CrossRef] [Green Version]
- Chalak, S.U.; Hasabnis, S.N.; Supe, V.S. Evaluation of Papaya Ring Spot Disease Resistant Lines under Western Maharashtra Conditions. J. Krishi Vigyan 2018, 7, 63–68. [Google Scholar] [CrossRef]
- Asghari, M.; Hasanlooe, A.R. Methyl jasmonate effectively enhanced some defense enzymes activity and Total Antioxidant content in harvested “Sabrosa” strawberry fruit. Food Sci. Nutr. 2016, 4, 377–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.P.; Fan, J.H.; Chen, H.J.; Yan, X.X.; Chen, P. Study on the control effect of three biopesticides on papaya ring spot. South China Fruits 2017, 46, 79–82. [Google Scholar]
- Badawy, M.E.I.; Rabea, E.I. Synthesis and antifungal property of N-(aryl) and quaternary N-(aryl) chitosan derivatives against Botrytis cinerea. Cellulose 2014, 21, 3121–3137. [Google Scholar] [CrossRef]
- Xiang, S.Y.; Lv, X.; He, L.H.; Shi, H.; Liao, S.Y.; Liu, C.Y.; Huang, Q.Q.; Li, X.Y.; He, X.T.; Chen, H.T.; et al. Dual-Action Pesticide Carrier That Continuously Induces Plant Resistance, Enhances Plant Anti-Tobacco Mosaic Virus Activity, and Promotes Plant Growth. J. Agric. Food Chem. 2019, 67, 10000–10009. [Google Scholar] [CrossRef]
- Fu, J.; Zhang, S.Y.; Wu, J.H.; Chen, Y.; Zhong, Y.; Zhou, Y.F.; Wang, J.G.; Chen, S.Y. Structural characterization of a polysaccharide from dry mycelium of Penicillium chrysogenum that induces resistance to Tobacco mosaic virus in tobacco plants. Int. J. Biol. Macromol. 2020, 156, 67–79. [Google Scholar] [CrossRef]
- Hao, Y.N.; Wang, K.H.; Wang, Z.W.; Liu, Y.X.; Ma, D.J.; Wang, Q.M. Luotonin A and Its Derivatives as Novel Antiviral and Antiphytopathogenic Fungus Agents. J. Agric. Food Chem. 2020, 68, 8764–8773. [Google Scholar] [CrossRef]
- Zu, G.C.; Gan, X.H.; Xie, D.D.; Yang, H.Y.; Zhang, A.W.; Li, S.Y.; Hu, D.Y.; Song, B.A. Design, Synthesis, and Anti-ToCV Activity of Novel 4(3H)-Quinazolinone Derivatives Bearing Dithioacetal Moiety. J. Agric. Food Chem. 2020, 68, 5539–5544. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.X.; Diao, H.L.; Wang, H.Y.; Wang, K.Y.; Zhao, M. Use of Ganoderma Lucidum polysaccharide to control cotton fusarium wilt, and the mechanism involved. Pestic. Biochem. Physiol. 2019, 158, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Ku, Y.; Xu, G.; Su, S.; Cao, C. Effects of biological agents on soil microbiology, enzyme activity and fruit quality of kiwifruit with root rot. Soil Res. 2021, 60, 279–293. [Google Scholar] [CrossRef]
- Abdel-Ghany, T.M.; Bakri, M.M. Effectiveness of a biological agent (Trichoderma harzianum and its culture filtrate) and a fungicide (methyl benzimacold-2-ylcarbamate) on the tomato rotting activity (growth, cellulolytic, and pectinolytic activities) of Alternaria solani. BioResources 2019, 14, 1591–1620. [Google Scholar] [CrossRef]
- Rajyaguru, R.H.; Thirumalaisamy, P.P.; Patel, K.G.; Thumar, J.T. Biochemical basis of genotypic and bio-agent induced stem rot resistance in groundnut. Legum. Res. Int. J. 2017, 40, 929–939. [Google Scholar] [CrossRef]
- Chen, Y.E.; Mao, H.T.; Wu, N.; Ma, J.; Yuan, M.; Zhang, Z.W.; Yuan, S.; Zhang, H.Y. Effects of Stripe Rust Infection on the Levels of Redox Balance and Photosynthetic Capacities in Wheat. Int. J. Mol. Sci. 2020, 21, 268. [Google Scholar] [CrossRef] [Green Version]
- Venzhik, Y.V.; Titov, A.F.; Talanova, V.V.; Frolova, S.A.; Talanov, A.V.; Nazarkina, Y.A. Influence of lowered temperature on the resistance and functional activity of the photosynthetic apparatus of wheat plants. Biol. Bull. 2011, 38, 132–137. [Google Scholar] [CrossRef]
- Sheikh, T.M.M.; Zhang, L.Y.; Zubair, M.; Hanif, A.; Li, P.; Farzand, A.; Ali, H.; Bilal, M.S.; Hu, Y.Q.; Chen, X.C.; et al. The Type III Accessory Protein HrpE of Xanthomonas oryzae pv. oryzae Surpasses the Secretion Role, and Enhances Plant Resistance and Photosynthesis. Microorganisms 2019, 7, 572. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.M.; Li, Z.X.; Ran, Q.J.; Li, P.; Peng, Z.H.; Zhang, J.R. ZmNF-YB16 Overexpression Improves Drought Resistance and Yield by Enhancing Photosynthesis and the Antioxidant Capacity of Maize Plants. Front. Plant Sci. 2018, 9, 709. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.Z.; Li, P.P.; Liu, X.W.; Li, C.; Zhang, S.K.; Wang, X.H.; Tao, X. Poly-γ-glutamic acid enhanced the drought resistance of maize by improving photosynthesis and affecting the rhizosphere microbial community. BMC Plant Biol. 2022, 22, 1–15. [Google Scholar] [CrossRef]
- Bwalya, J.; Alazem, M.; Kim, K.H. Photosynthesis-related genes induce resistance against soybean mosaic virus: Evidence for involvement of the RNA silencing pathway. Mol. Plant Pathol. 2022, 23, 543–560. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviations | Chitosan | Lentinan | Ningnanmycin | Note |
|---|---|---|---|---|
| CK | 0 | 0 | 0 | inoculated with PRSV |
| C | 0.5 | 0 | 0 | |
| L | 0 | 10 | 0 | |
| N | 0 | 0 | 0.05 | |
| CL | 0.5 | 10 | 0 | |
| CN | 0.5 | 0 | 0.05 | |
| LN | 0 | 10 | 0.05 | |
| CLN | 0.5 | 10 | 0.05 |
| Factors | Pot Experiment | Field Experiment | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| POD | SOD | PPO | POD | SOD | PPO | |||||||||||||
| 7th | 14th | 21st | 7th | 14th | 21st | 7th | 14th | 21st | 7th | 14th | 21st | 7th | 14th | 21st | 7th | 14th | 21st | |
| C× | *** | *** | Ns | Ns | ** | Ns | Ns | *** | *** | Ns | * | ** | *** | *** | *** | *** | *** | *** |
| L× | *** | Ns | *** | Ns | ** | *** | *** | *** | *** | * | ** | ** | * | *** | ** | *** | ** | *** |
| N× | Ns | *** | Ns | *** | *** | *** | *** | *** | *** | * | Ns | ** | *** | *** | *** | *** | *** | *** |
| C×L | Ns | * | *** | *** | * | *** | Ns | Ns | *** | ** | Ns | Ns | ** | *** | *** | *** | *** | *** |
| C×N | Ns | Ns | ** | ** | ** | *** | Ns | Ns | ** | Ns | Ns | *** | ** | *** | *** | *** | *** | ** |
| L×N | * | *** | *** | *** | *** | ** | ** | * | ** | * | Ns | ** | Ns | *** | Ns | *** | *** | Ns |
| C×L×N | Ns | * | *** | Ns | *** | Ns | Ns | *** | *** | Ns | ** | * | Ns | * | Ns | *** | Ns | ** |
| Treatments | Growth and Development Index | |||
|---|---|---|---|---|
| Plant Height (m plant−1) | Stem Diameter (cm plant−1) | Chlorophyll SPAD | Dry Weight (g plant−1) | |
| CK | 19.48 ± 2.25 e | 5.03 ± 0.32 d | 9.44 ± 0.89 e | 1.59 ± 0.11 e |
| C | 22.80 ± 3.71 de | 8.86 ± 0.43 c | 10.72 ± 0.97 e | 3.25 ± 0.48 cd |
| L | 24.41 ± 2.91 cde | 9.45 ± 1.39 bc | 11.32 ± 1.83 de | 3.28 ± 0.09 cd |
| N | 26.92 ± 1.42 bcd | 10.21 ± 0.29 bc | 15.98 ± 1.34 bc | 4.99 ± 1.06 ab |
| CL | 28.54 ± 3.58 abc | 11.07 ± 0.55 ab | 18.67 ± 2.11 b | 4.85 ± 0.79 ab |
| CN | 32.38 ± 4.15 ab | 11.99 ± 0.31 a | 14.97 ± 1.25 c | 5.59 ± 0.32 a |
| LN | 28.71 ± 3.23 abc | 10.12 ± 0.82 bc | 18.21 ± 1.49 b | 4.23 ± 0.02 bc |
| CLN | 33.21 ± 2.77 a | 11.94 ± 1.87 a | 25.67 ± 1.46 a | 4.64 ± 0.91 ab |
| NT | 21.29 ± 1.31 e | 9.46 ± 0.57 bc | 13.58 ± 2.22 cd | 2.42 ± 0.32 de |
| ANOVA | ||||
| C× | ** | *** | *** | *** |
| L× | * | *** | *** | Ns |
| N× | *** | *** | *** | *** |
| C×L | Ns | Ns | *** | Ns |
| C×N | Ns | Ns | Ns | * |
| L×N | Ns | *** | Ns | *** |
| C×L×N | Ns | Ns | Ns | Ns |
| Treatments | Plant Height (m plant−1) | Stem Diameter (cm plant−1) | Chlorophyll SPAD |
|---|---|---|---|
| CK | 2.47 ± 0.06 f | 36.67 ± 4.36 bc | 9.42 ± 1.45 f |
| C | 2.52 ± 0.06 e | 33.33 ± 3.51 d | 15.01 ± 1.51 c |
| L | 2.64 ± 0.05 cd | 36.00 ± 1.00 bcd | 13.64 ± 2.61 d |
| N | 2.73 ± 0.05 b | 38.33 ± 2.08 abc | 15.59 ± 2.96 c |
| CL | 2.67 ± 0.06 c | 40.67 ± 4.51 a | 19.71 ± 2.73 b |
| CN | 2.62 ± 0.10 d | 35.33 ± 0.58 cd | 24.15 ± 1.43 a |
| LN | 2.84 ± 0.03 a | 38.67 ± 0.58 ab | 19.84 ± 2.94 b |
| CLN | 2.86 ± 0.09 a | 41.00 ± 1.73 a | 20.23 ± 1.54 b |
| NT | 2.53 ± 0.08 e | 35.33 ± 3.79 cd | 11.82 ± 0.98 e |
| ANOVA | |||
| C× | Ns | Ns | *** |
| L× | *** | *** | *** |
| N× | *** | * | *** |
| C×L | ** | *** | *** |
| C×N | *** | Ns | * |
| L×N | Ns | Ns | *** |
| C×L×N | ** | Ns | *** |
| Treatments | Single Fruit Weight (kg per fruit−1) | Fruit Length (cm per fruit−1) | Fruit Width (cm per fruit−1) | Water Content (%) | Reducing Sugar (mg g−1) | Sweetness |
|---|---|---|---|---|---|---|
| CK | 0.76 ± 0.15 d | 22.00 ± 1.00 b | 23.33 ± 4.93 c | 85.82 ± 1.19 d | 3.57 ± 0.02 e | 10.76 ± 0.29 e |
| C | 1.18 ± 0.20 abc | 24.00 ± 1.00 a | 39.00 ± 1.73 ab | 90.42 ± 0.55 ab | 3.81 ± 0.02 b | 6.43 ± 0.20 h |
| L | 1.34 ± 0.21 a | 25.00 ± 0.48 a | 41.67 ± 5.77 ab | 88.85 ± 0.05 c | 3.94 ± 0.01 a | 13.33 ± 0.10 c |
| N | 1.04 ± 0.03 bc | 24.33 ± 1.15 a | 36.67 ± 2.89 ab | 90.72 ± 0.61 a | 3.71 ± 0.01 d | 11.83 ± 0.17 d |
| CL | 1.31 ± 0.09 a | 25.00 ± 0.67 a | 43.33 ± 1.53 a | 88.61 ± 0.66 c | 3.77 ± 0.03 c | 15.72 ± 0.25 a |
| CN | 1.29 ± 0.12 a | 25.00 ± 0.85 a | 41.33 ± 3.06 ab | 89.10 ± 1.25 c | 3.81 ± 0.01 b | 10.26 ± 0.29 f |
| LN | 1.27 ± 0.04 ab | 25.33 ± 1.15 a | 39.00 ± 4.58 ab | 88.28 ± 1.82 c | 3.77 ± 0.02 c | 15.38 ± 0.13 b |
| CLN | 1.26 ± 0.13 ab | 24.67 ± 0.58 a | 42.00 ± 4.58 ab | 89.33 ± 1.92 bc | 3.70 ± 0.04 d | 9.43 ± 0.01 g |
| NT | 0.99 ± 0.11 c | 22.00 ± 0.58 b | 35.00 ± 2.65 b | 84.08 ± 1.99 e | 3.57 ± 0.03 e | 12.10 ± 0.58 h |
| ANOVA | ||||||
| C× | ** | Ns | ** | ** | *** | *** |
| L× | *** | ** | ** | Ns | *** | *** |
| N× | Ns | * | Ns | ** | *** | *** |
| C×L | ** | * | * | Ns | *** | *** |
| C×N | Ns | Ns | Ns | *** | * | *** |
| L×N | * | * | * | ** | *** | *** |
| C×L×N | Ns | Ns | Ns | *** | *** | *** |
| Sample | Total Pair Reads | Unmapped Pair Reads | Unique Mapped Pair Reads | Multiple Mapped Pair Reads | Mapping Ratio |
|---|---|---|---|---|---|
| CK1 | 31,635,080 | 6,301,472 (19.92%) | 25,237,526 (79.78%) | 96,082 (0.30%) | 80.08% |
| CK2 | 36,613,018 | 9,377,139 (25.61%) | 27,134,133 (74.11%) | 101,746 (0.28%) | 74.39% |
| CK3 | 34,442,498 | 6,132,542 (17.81%) | 28,205,012 (81.89%) | 104,944 (0.30%) | 82.19% |
| LN_1 | 37,993,586 | 6,308,417 (16.60%) | 31,559,478 (83.07%) | 125,691 (0.33%) | 83.40% |
| LN_2 | 35,965,986 | 5,583,588 (15.52%) | 30,250,979 (84.11%) | 131,419 (0.37%) | 84.48% |
| LN_3 | 42,409,640 | 6,849,230 (16.15%) | 35,368,850 (83.40%) | 191,560 (0.45%) | 83.85% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, H.; Yan, X.; Fu, M.; Liu, D.; Awan, A.W.; Chen, P.; Rasheed, S.M.; Gao, L.; Zhang, R. Interactive Effect of Biological Agents Chitosan, Lentinan and Ningnanmycin on Papaya Ringspot Virus Resistance in Papaya (Carica papaya L.). Molecules 2022, 27, 7474. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27217474
Fan H, Yan X, Fu M, Liu D, Awan AW, Chen P, Rasheed SM, Gao L, Zhang R. Interactive Effect of Biological Agents Chitosan, Lentinan and Ningnanmycin on Papaya Ringspot Virus Resistance in Papaya (Carica papaya L.). Molecules. 2022; 27(21):7474. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27217474
Chicago/Turabian StyleFan, Heling, Xingxiang Yan, Mingqing Fu, Difa Liu, Abdul Waheed Awan, Ping Chen, Syed Majid Rasheed, Ling Gao, and Rongping Zhang. 2022. "Interactive Effect of Biological Agents Chitosan, Lentinan and Ningnanmycin on Papaya Ringspot Virus Resistance in Papaya (Carica papaya L.)" Molecules 27, no. 21: 7474. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27217474