
Treatment with Glyphosate Induces Tolerance of Citrus Pathogens to Glyphosate and Fungicides but Not to 1,8-Cineole
 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results
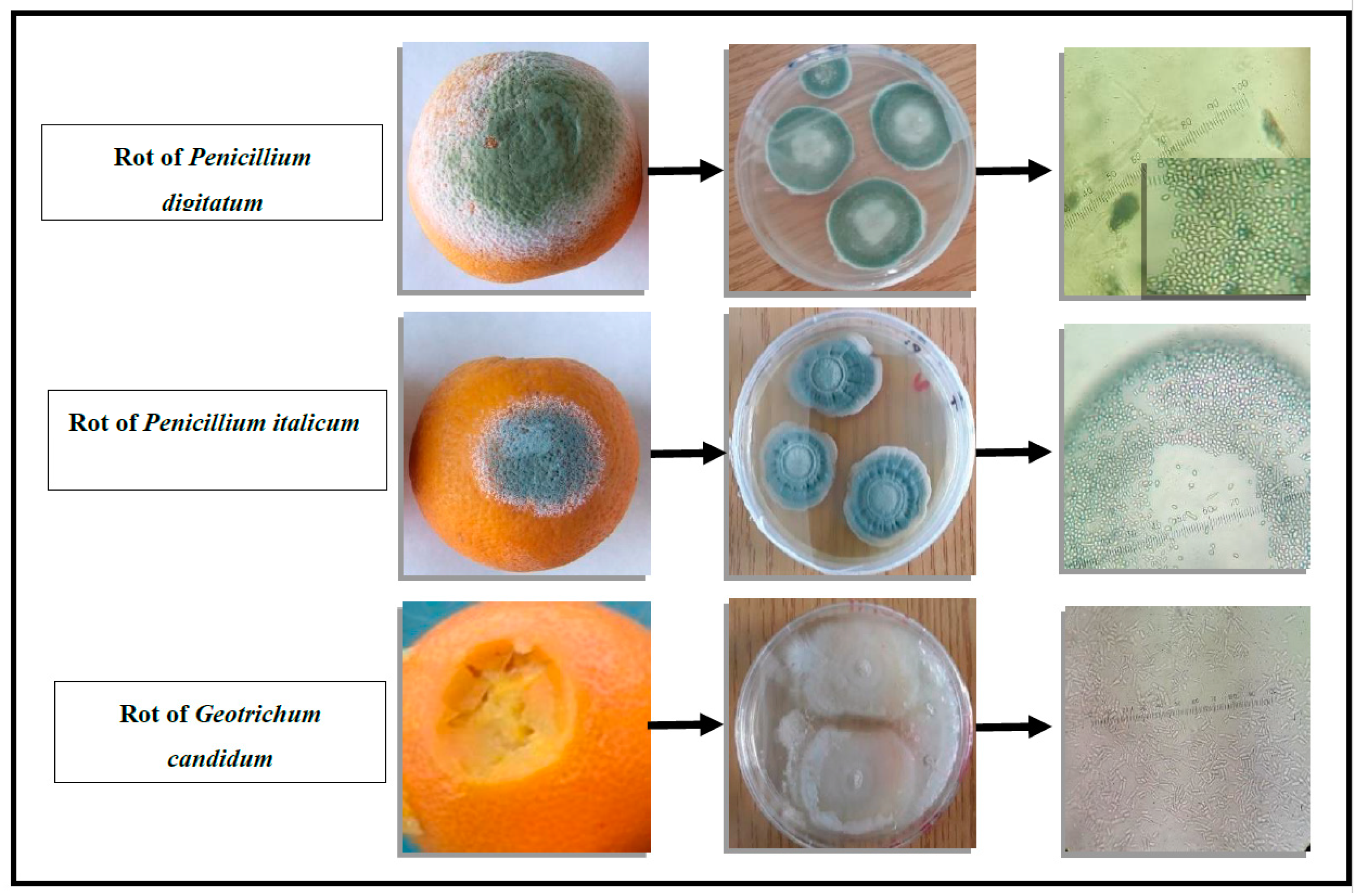
2.1. Isolation and Identification of Fungi Responsible for Postharvest Citrus Fruit Rot
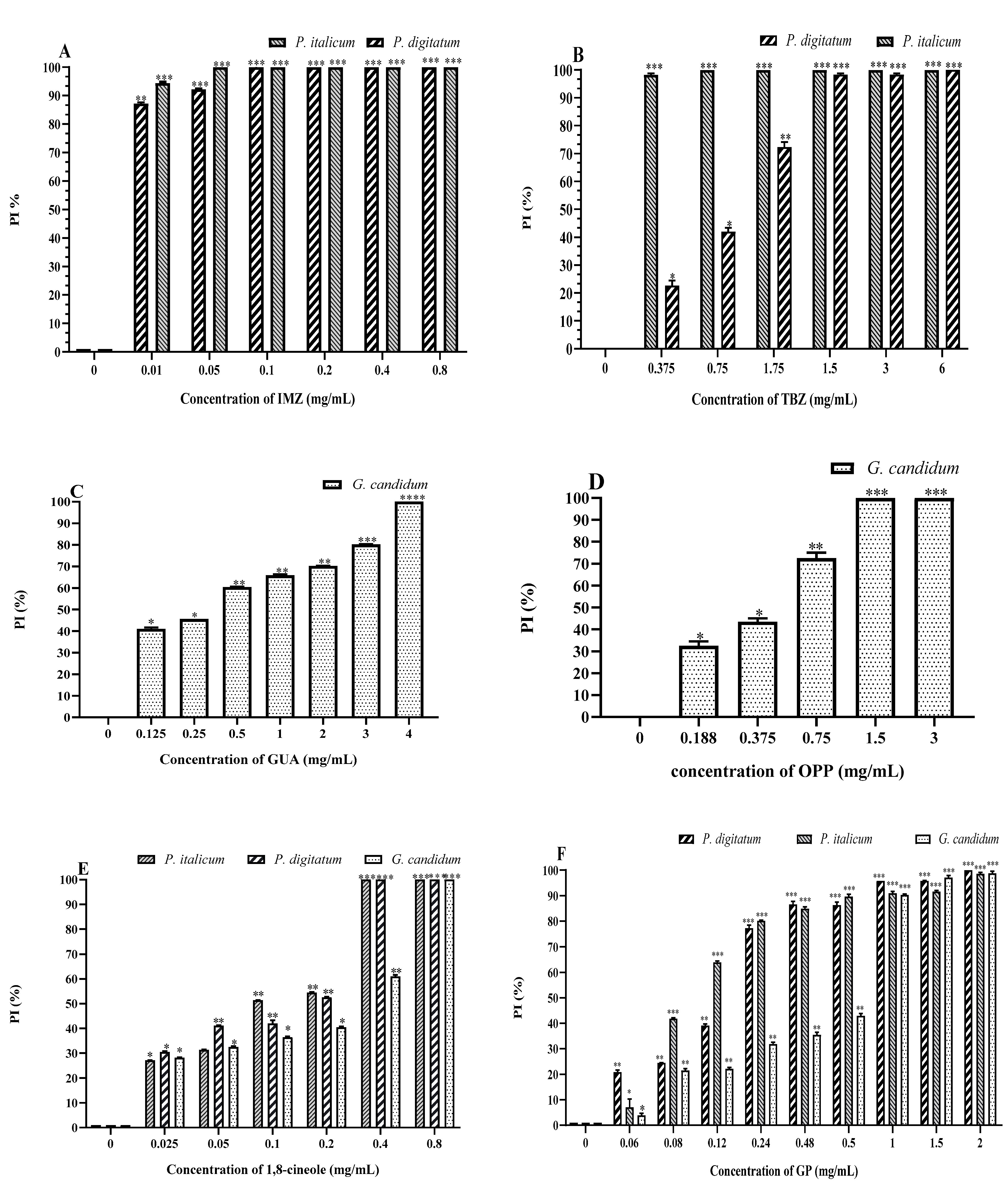
2.2. Antifungal Activity
2.3. Inhibition of Spore Germination
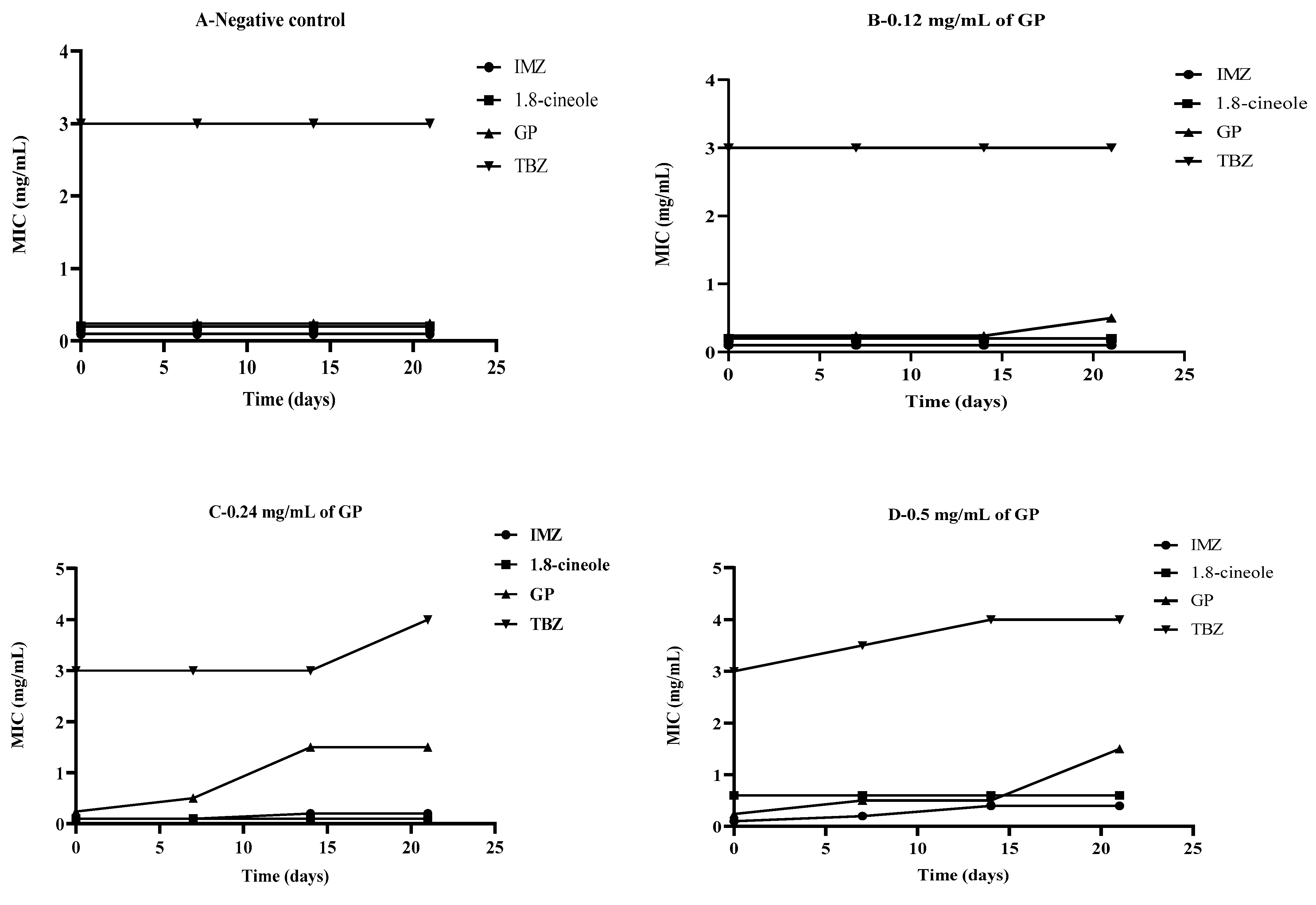
2.4. The effect of Glyphosate on Fungal Sensitivity to Different Antifungals In Vitro
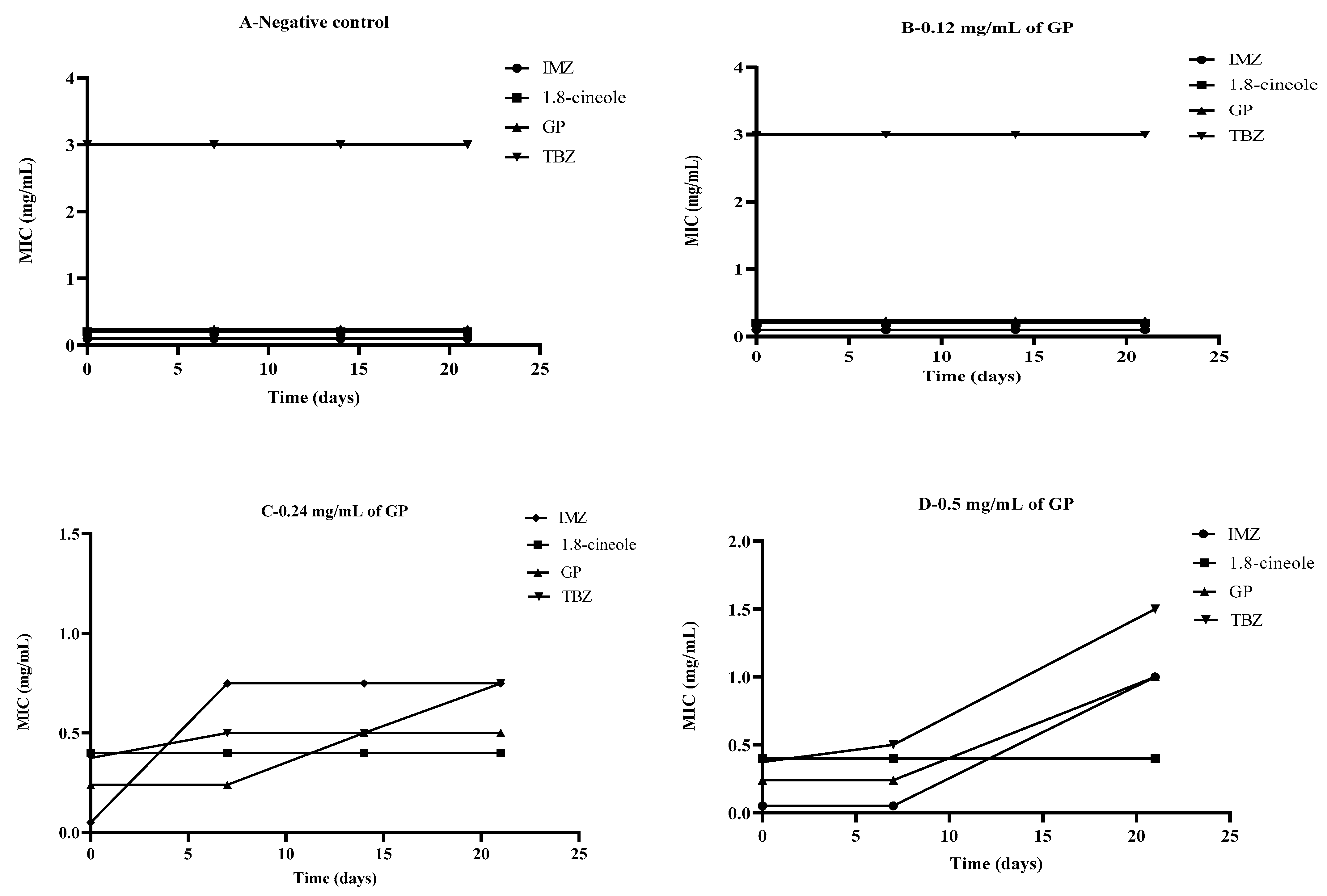
2.5. The Effect of Glyphosate on Fungal Sensitivity to Different Antifungals In Vitro in Contaminated Soil
3. Discussion
4. Material and Methods
4.1. Chemicals and Growth Media
4.2. Sampling
4.3. Morphological Identification of Fungal Species
4.4. Spore Suspension
4.5. In Vitro Antifungal Activity
4.6. Spore Germination Assay
4.7. The Effect of Glyphosate on Fungal Sensitivity to Different Antifungals In Vitro
4.8. The Effect of Glyphosate on Fungal Sensitivity to Different Antifungals In Vitro in Contaminated Soil
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
Sample Availability
References
- FAO. Citrus Fruit Statistical Compendium 2020. In Citrus Fruit Fresh and Processed Statistical Bulletin 2020; FAO: Rome, Italy, 2021; Available online: https://www.fao.org/3/cb6492en/cb6492en.pdf (accessed on 8 November 2022).
- Global Citrus Outlook. 2019. Available online: https://worldcitrusorganisation.org/wp-content/uploads/2020/01/Citrus-Market-Trends-2019.pdf (accessed on 16 September 2022).
- Martínez-Blay, V.; Taberner, V.; Pérez-Gago, M.B.; Palou, L. Control of major citrus postharvest diseases by sulfur-containing food additives. Int. J. Food Microbiol. 2020, 330, 108713. [Google Scholar] [CrossRef] [PubMed]
- Moroccan Minister of Agriculture. “Citrus industry,” The Minister of Agriculture. 2019. Available online: https://www.agriculture.gov.ma/fr/filiere/agrumicole (accessed on 16 September 2022).
- Talibi, I.; Askarne, L.; Boubaker, H.; Boudyach, E.H.; Msanda, F.; Saadi, B.; Ait Ben Aoumar, A. Antifungal activity of some Moroccan plants against Geotrichum candidum, the causal agent of postharvest citrus sour rot. Crop Prot. 2012, 35, 41–46. [Google Scholar] [CrossRef]
- Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. The future of citrus fruit: The impact of climate change on citriculture. Metod. Sci. Stud. J. 2021, 12, 123–129. [Google Scholar] [CrossRef]
- Smilanick, J.L.; Erasmus, A.; Palou, L. Citrus Fruits. In Postharvest Pathology of Fresh Horticultural Produce; CRC Press: Boca Raton, FL, USA, 2019; pp. 3–53. ISBN 9781315209180. Available online: https://www.taylorfrancis.com/chapters/edit/10.1201/9781315209180-1/citrus-fruits-joseph-smilanick-arno-erasmus-lluís-palou (accessed on 16 September 2022).
- de Vilhena Araújo, É.; Vendramini, P.H.; Costa, J.H.; Eberlin, M.N.; Montagner, C.C.; Fill, T.P. Determination of tryptoquialanines A and C produced by Penicillium digitatum in oranges: Are we safe? Food Chem. 2019, 301, 125285. [Google Scholar] [CrossRef]
- Kanashiro, A.M.; Akiyama, D.Y.; Kupper, K.C.; Fill, T.P. Penicillium italicum: An Underexplored Postharvest Pathogen. Front. Microbiol. 2020, 11, 606852. [Google Scholar] [CrossRef]
- Saini, A.; Parkash, V.; Kumar, V.; Gaur, A.; Chirag, H. Detection of mycotoxins from some micro and macro endophytic ascomycetes fungi. Int. J. Environ. Agric. Biotechnol. 2019, 6, 11. [Google Scholar] [CrossRef]
- Erasmus, A.; Lennox, C.L.; Korsten, L.; Lesar, K.; Fourie, P.H. Imazalil resistance in Penicillium digitatum and P. italicum causing citrus postharvest green and blue mould: Impact and options. Postharvest Biol. Technol. 2015, 107, 66–76. [Google Scholar] [CrossRef]
- Hao, W.; Zhong, G.; Hu, M.; Luo, J.; Weng, Q.; Rizwan-ul-Haq, M. Control of citrus postharvest green and blue mold and sour rot by tea saponin combined with imazalil and prochloraz. Postharvest Biol. Technol. 2010, 56, 39–43. [Google Scholar] [CrossRef]
- Altieri, G.; Di Renzo, G.C.; Genovese, F.; Calandra, M.; Strano, M.C. A new method for the postharvest application of imazalil fungicide to citrus fruit. Biosyst. Eng. 2013, 115, 434–443. [Google Scholar] [CrossRef]
- Sanzani, S.M.; Reverberi, M.; Geisen, R. Mycotoxins in harvested fruits and vegetables: Insights in producing fungi, biological role, conducive conditions, and tools to manage postharvest contamination. Postharvest Biol. Technol. 2016, 122, 95–105. [Google Scholar] [CrossRef]
- Zhang, H.; Mahunu, G.K.; Castoria, R.; Yang, Q.; Apaliya, M.T. Recent developments in the enhancement of some postharvest biocontrol agents with unconventional chemicals compounds. Trends Food Sci. Technol. 2018, 78, 180–187. [Google Scholar] [CrossRef]
- Guillot, J.; Dannaoui, É. La résistance aux antifongiques: Importance en médecine humaine et vétérinaire. Bull. Acad. Vet. Fr. 2015, 168, 314–319. [Google Scholar] [CrossRef]
- Kinay, P.; Mansour, M.F.; Mlikota Gabler, F.; Margosan, D.A.; Smilanick, J.L. Characterization of fungicide-resistant isolates of Penicillium digitatum collected in California. Crop Prot. 2007, 26, 647–656. [Google Scholar] [CrossRef]
- Dayan, F.E.; Cantrell, C.L.; Duke, S.O. Natural products in crop protection. Bioorganic Med. Chem. 2009, 17, 4022–4034. [Google Scholar] [CrossRef]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 63, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef]
- Mesnage, R.; Antoniou, M.N. Facts and Fallacies in the Debate on Glyphosate Toxicity. Front. Public Heal. 2017, 5, 316. [Google Scholar] [CrossRef] [PubMed]
- Van Bruggen, A.H.C.; He, M.M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.R.; Morris, J.G. Environmental and health effects of the herbicide glyphosate. Sci. Total Environ. 2018, 616–617, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Ben Khadda, Z.; Fagroud, M.; El Karmoudi, Y.; Ezrari, S.; Berni, I.; De Broe, M.; Behl, T.; Bungau, S.G.; Houssaini, T.S. Farmers’ knowledge, attitudes, and perceptions regarding carcinogenic pesticides in fez meknes region (Morocco). Int. J. Environ. Res. Public Health 2021, 18, 10879. [Google Scholar] [CrossRef] [PubMed]
- Pollegioni, L.; Schonbrunn, E.; Siehl, D. Molecular basis of glyphosate resistance—Different approaches through protein engineering. FEBS J. 2011, 278, 2753–2766. [Google Scholar] [CrossRef]
- Xing, Y.; Wu, S.; Men, Y. Exposure to Environmental Levels of Pesticides Stimulates and Diversifies Evolution in Escherichia coli toward Higher Antibiotic Resistance. Environ. Sci. Technol. 2020, 54, 8770–8778. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, J.; Wang, L.; Zhai, Z. Glyphosate escalates horizontal transfer of conjugative plasmid harboring antibiotic resistance genes. Bioengineered 2021, 12, 63–69. [Google Scholar] [CrossRef]
- Barbosa da Costa, N.; Hébert, M.-P.; Fugère, V.; Terrat, Y.; Fussmann, G.F.; Gonzalez, A.; Shapiro, B.J. A Glyphosate-Based Herbicide Cross-Selects for Antibiotic Resistance Genes in Bacterioplankton Communities. mSystems 2022, 7, e01482-21. [Google Scholar] [CrossRef] [PubMed]
- Tao, B.; Shao, B.H.; Qiao, Y.X.; Wang, X.Q.; Chang, S.J.; Qiu, L.J. Identification and functional analysis of a new glyphosate resistance gene from a fungus cDNA library. Pestic. Biochem. Physiol. 2017, 140, 65–68. [Google Scholar] [CrossRef]
- Guo, J.; Song, X.; Zheng, C.; Sun, S.; Zhuang, B.; Tao, B. Transcriptome analysis and identification of candidate genes involved in glyphosate resistance in the fungus Fusarium verticillioides. J. Environ. Sci. Health 2021, 56, 658–669. [Google Scholar] [CrossRef]
- Raoult, D.; Hadjadj, L.; Baron, S.A.; Rolain, J.M. Role of glyphosate in the emergence of antimicrobial resistance in bacteria? J. Antimicrob. Chemother. 2021, 76, 1655–1657. [Google Scholar] [CrossRef] [PubMed]
- Kurenbach, B.; Gibson, P.S.; Hill, A.M.; Bitzer, A.S.; Silby, M.W.; Godsoe, W.; Heinemann, J.A. Herbicide ingredients change Salmonella enterica sv. Typhimurium and Escherichia coli antibiotic responses. Microbiology 2017, 163, 1791–1801. [Google Scholar] [CrossRef]
- Al Zuhairi, J.J.M.J.; Jookar Kashi, F.; Rahimi-Moghaddam, A.; Yazdani, M. Antioxidant, cytotoxic and antibacterial activity of Rosmarinus officinalis L. essential oil against bacteria isolated from urinary tract infection. Eur. J. Integr. Med. 2020, 38, 101192. [Google Scholar] [CrossRef]
- Belasli, A.; Ben Miri, Y.; Aboudaou, M.; Aït Ouahioune, L.; Montañes, L.; Ariño, A.; Djenane, D. Antifungal, antitoxigenic, and antioxidant activities of the essential oil from laurel (Laurus nobilis L.): Potential use as wheat preservative. Food Sci. Nutr. 2020, 89, 4717–4729. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.N.; Lam, N.T.T.; Cuong, N.T.; Ngan, T.Q.; Truong, N.C.; Ogunwande, I.A. Composition of Essential Oil of Cinnamomum tetragonum. Chem. Nat. Compd. 2020, 56, 545–547. [Google Scholar] [CrossRef]
- Aleksic Sabo, V.; Knezevic, P. Antimicrobial activity of Eucalyptus camaldulensis Dehn. plant extracts and essential oils: A review. Ind. Crops Prod. 2019, 132, 413–429. [Google Scholar] [CrossRef] [PubMed]
- Regnier, T.; Combrinck, S.; Veldman, W.; Plooy, W. Du Application of essential oils as multi-target fungicides for the control of Geotrichum citri-aurantii and other postharvest pathogens of citrus. Ind. Crop. Prod. 2014, 61, 151–159. [Google Scholar] [CrossRef]
- Vitoratos, A.; Bilalis, D.; Karkanis, A.; Efthimiadou, A. Antifungal Activity of Plant Essential Oils Against Botrytis cinerea, Penicillium italicum and Penicillium digitatum. Not. Bot. Horti Agrobot. Cluj-Napoca 2013, 41, 86–92. [Google Scholar] [CrossRef]
- Klieber, A.; Scott, E.; Wuryatmo, E. Effect of method of application on antifungal efficacy of citral againstpostharvest spoilage fungi of citrus in culture. Australas. Plant Pathol. 2002, 31, 329–332. [Google Scholar] [CrossRef]
- Trabelsi, D.; Hamdane, A.M.; Said, M.B.; Abdrrabba, M. Chemical Composition and Antifungal Activity of Essential Oils from Flowers, Leaves and Peels of Tunisian Citrus aurantium Against Penicillium digitatum and Penicillium italicum. J. Essent. Oil-Bearing Plants 2016, 19, 1660–1674. [Google Scholar] [CrossRef]
- Sánchez-Torres, P.; Tuset, J.J. Molecular insights into fungicide resistance in sensitive and resistant Penicillium digitatum strains infecting citrus. Postharvest Biol. Technol. 2011, 59, 159–165. [Google Scholar] [CrossRef]
- Fatima, S.; Iram, S. Identification of Fungal Pathotypes Associated with Skin Disorders of Citrus Reticulata Blanco Through Classical and Molecular Approach. Pakistan J. Agric. Res. 2019, 32, 95. [Google Scholar] [CrossRef]
- Wang, C.-J.K.; Zabel, R.A. Identification Manual for Fungi from Utility Poles in the Eastern United States; American Type Culture Collection: Manassas, VA, USA, 1990. [Google Scholar]
- Garg, S.; Naidu, J.; Singh, S.M.; Nawange, S.R.; Jharia, N.; Saxena, M. In vitro activity of terbinafine against Indian clinical isolates of Candida albicans and non-albicans using a macrodilution method. J. Mycol. Med. 2006, 16, 119–125. [Google Scholar] [CrossRef]
- Talibi, I.; Askarne, L.; Boubaker, H.; Boudyach, E.H.; Aoumar, A.A. Ben In vitro and in vivo antifungal activities of organic and inorganic salts against citrus sour rot agent geotrichum candidum. Plant Pathol. J. 2011, 10, 138–145. [Google Scholar] [CrossRef]
- Saghrouchni, H.; El Barnossi, A.; Chefchaou, H.; Mzabi, A.; Tanghort, M.; Remmal, A.; Fouzia, C. Study the effect of carvacrol, eugenol and thymol on fusariums sp responsible for lolium perenne fusariosis. EM Int. 2020, 26, 1059–1067. [Google Scholar]
- Bhandari, M.; Bhandari, N.; Dhital, M. Effect of fungicide and essential oils amended wax coating on quality and shelf life of sweet orange (Citrus sinensis Osbeck). J. Hortic. Sci. 2019, 16, 2021. [Google Scholar] [CrossRef]
- Bhowal, M.; Gopal, M. Eucalyptol: Safety and Pharmacological Profile. RGUHS J. Pharm. Sci. 2016, 5, 125–131. [Google Scholar] [CrossRef]
- El Barnossi, A.; Saghrouchni, H.; Moussaid, F.; Chahmi, H.; Housseini, A.I. Microbiological study of effects of solid organic waste (chicken droppings and sheep manure) decomposed in the soil used for Pisum sativum cultivation. Int. J. Environ. Stud. 2019, 77, 830–842. [Google Scholar] [CrossRef]
- Saghrouchni, H.; El Barnossi, A.; Salamatullah, A.M.; Bourhia, M.; Alzahrani, A.; Alkaltham, M.S.; Alyahya, H.K.; Tahiri, N.E.H.; Imtara, H.; Var, I. Carvacrol: A promising environmentally friendly agent to fight seeds damping-off diseases induced by fungal species. agronomy 2021, 11, 985. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antifungal Agent | Fungal Pathogen | MIC (mg/mL) |
|---|---|---|
| IMZ | P. digitatum | 0.01 |
| P. italicum | 0.05 | |
| TBZ | P. digitatum | 3 |
| P. italicum | 0.375 | |
| GUA | G. candidum | 4 |
| OPP | G. candidum | 1.5 |
| 1,8-Cineole | P. digitatum | 0.4 |
| P. italicum | 0.2 | |
| G. candidum | 0.4 | |
| GP | P. digitatum | 0.24 |
| P. italicum | 0.24 | |
| G. candidum | 1 |
| Antifungal Agent | Number of Colonies (CFU) | ||||
|---|---|---|---|---|---|
| IMZ | 0.05 mg/mL | 0.1 mg/mL | 0.2 mg/mL | 0.4 mg/mL | 0.8 mg/mL |
| 9 ± 20 | 0 | 0 | 0 | 0 | |
| TBZ | 0.375 mg/mL | 0.75 mg/mL | 1.5 mg/mL | 3 mg/mL | 6 mg/mL |
| 11 ± 30 | 0 | 0 | 0 | 0 | |
| 1,8-Cineole | 0.025 mg/mL | 0.05 mg/mL | 0.1 mg/mL | 0.2 mg/mL | 0.4 mg/mL |
| 245 ± 15 | 112 ± 10 | 59 ± 66 | 3 ± 10 | 0 | |
| GP | 0.12 mg/mL | 0.24 mg/mL | 0.5 mg/mL | 1 mg/mL | 1.5 mg/mL |
| 19 ± 20 | 4 ± 10 | 0 | 0 | 0 | |
| Negative control | ≥300 | ≥300 | ≥300 | ≥300 | ≥300 |
| Antifungal Agent | Number of Colonies (CFU) | ||||
|---|---|---|---|---|---|
| IMZ | 0.05 mg/mL | 0.1 mg/mL | 0.2 mg/mL | 0.4 mg/mL | 0.8 mg/mL |
| 11 ± 20 | 0 | 0 | 0 | 0 | |
| TBZ | 0.375 mg/mL | 0.75 mg/mL | 1.5 mg/mL | 3 mg/mL | 6 mg/mL |
| 90 ± 50 | 25 ± 30 | 0 | 0 | 0 | |
| 1,8-Cineole | 0.025 mg/mL | 0.05 mg/mL | 0.1 mg/mL | 0.2 mg/mL | 0.4 mg/mL |
| 270 ± 11 | 188 ± 14 | 47 ± 90 | 4 ± 50 | 0 | |
| GP | 0.12 mg/mL | 0.24 mg/mL | 0.5 mg/mL | 1 mg/mL | 1.5 mg/mL |
| 56 ± 30 | 9 ± 20 | 0 | 0 | 0 | |
| Negative control | ≥300 | ≥300 | ≥300 | ≥300 | ≥300 |
| Antifungal Agent | Number of Colonies (CFU) | ||||
|---|---|---|---|---|---|
| OPP | 0.188 mg/mL | 0.375 mg/mL | 0.75 mg/mL | 1.5 mg/mL | 3 mg/mL |
| ≥300 | 88 ± 90 | 48 ± 40 | 0 | 0 | |
| GUA | 0.125 mg/mL | 0.05 mg/mL | 1 mg/mL | 2 mg/mL | 4 mg/mL |
| ≥300 | 78 ± 90 | 16 ± 50 | 4 ± 20 | 0 | |
| 1,8-Cineole | 0.025 mg/mL | 0.05 mg/mL | 0.1 mg/mL | 0.2 mg/mL | 0.4 mg/mL |
| 290 ± 15 | 90 ± 11 | 42 ± 50 | 0 | 0 | |
| GP | 0.12 mg/mL | 0.24 mg/mL | 0.5 mg/mL | 1 mg/mL | 1.5 mg/mL |
| 150 ± 18 | 52 ± 20 | 3 ± 10 | 0 | 0 | |
| Negative control | ≥300 | ≥300 | ≥300 | ≥300 | ≥300 |
| Fungal Pathogen | Initial MIC Value | MIC after MSubculturing |
|---|---|---|
| P. digitatum | 0.24 mg/mL | 1 mg/mL |
| P. italicum | 0.24 mg/mL | 0.5 mg/mL |
| G. candidum | 1 mg/mL | 2 mg/mL |
| Fungal Pathogen | MIC of IMZ before and after Subculturing | MIC of TBZ before and after Subculturing | MIC of GUA before and after Subculturing | MIC of OPP before and after Subculturing |
|---|---|---|---|---|
| P. digitatum (7 subcultures) | from 0.1 to 0.4 mg/Ml | No change (1.5 mg/mL) | - | - |
| P. italicum (10 subcultures) | from 0.5 to 0.8 mg/Ml | from 0.375 to 3 mg/mL | - | - |
| G. candidum (12 subcultures) | - | - | from 1.5 to 3 mg/mL | from 1 to 2 mg/mL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tahiri, N.E.H.; Saghrouchni, H.; Hamamouch, N.; Khomsi, M.E.; Alzahrani, A.; Salamatullah, A.M.; Badiaa, L.; Lrhorfi, L.A. Treatment with Glyphosate Induces Tolerance of Citrus Pathogens to Glyphosate and Fungicides but Not to 1,8-Cineole. Molecules 2022, 27, 8300. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27238300
Tahiri NEH, Saghrouchni H, Hamamouch N, Khomsi ME, Alzahrani A, Salamatullah AM, Badiaa L, Lrhorfi LA. Treatment with Glyphosate Induces Tolerance of Citrus Pathogens to Glyphosate and Fungicides but Not to 1,8-Cineole. Molecules. 2022; 27(23):8300. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27238300
Chicago/Turabian StyleTahiri, Nor EL Houda, Hamza Saghrouchni, Noureddine Hamamouch, Mostafa El Khomsi, Abdulhakeem Alzahrani, Ahmad Mohammad Salamatullah, Lyoussi Badiaa, and Lalla Aicha Lrhorfi. 2022. "Treatment with Glyphosate Induces Tolerance of Citrus Pathogens to Glyphosate and Fungicides but Not to 1,8-Cineole" Molecules 27, no. 23: 8300. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27238300