
Impact of a Shorter Brine Soaking Time on Nutrient Bioaccessibility and Peptide Formation in 30-Months-Ripened Parmigiano Reggiano Cheese

 , , , ,
, , , ,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Fatty Acid Bioaccessibility
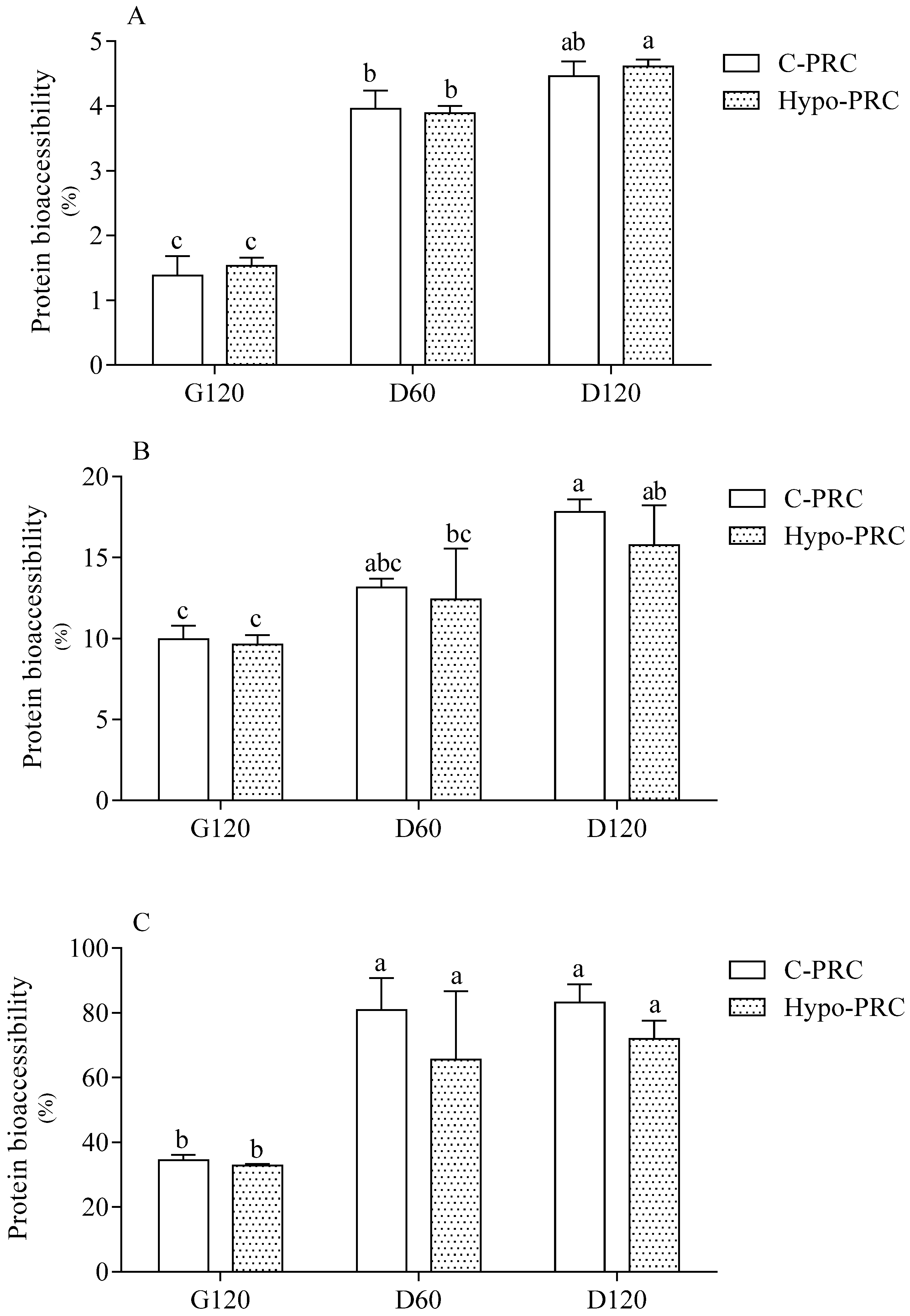
2.2. Protein Bioaccessibility
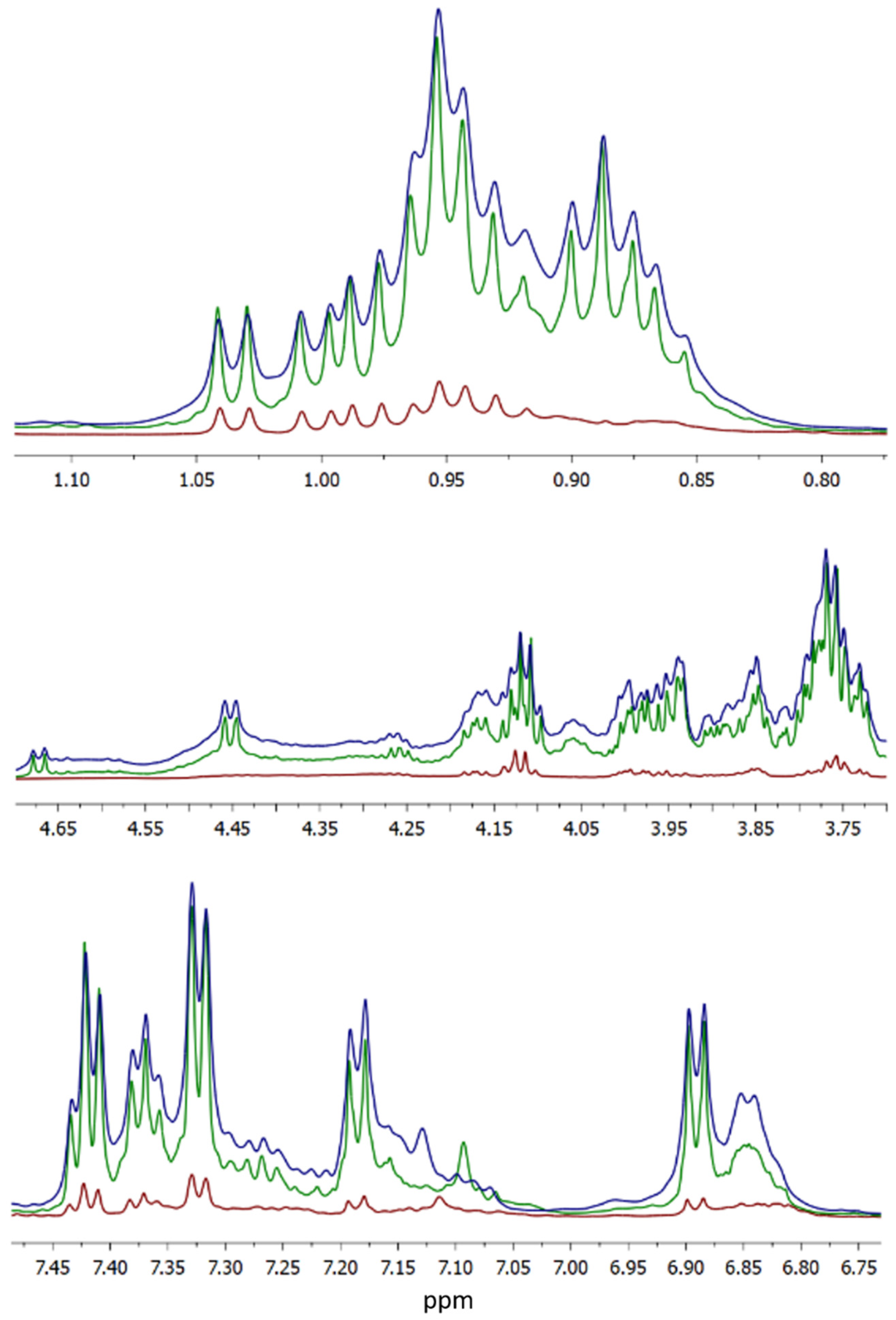
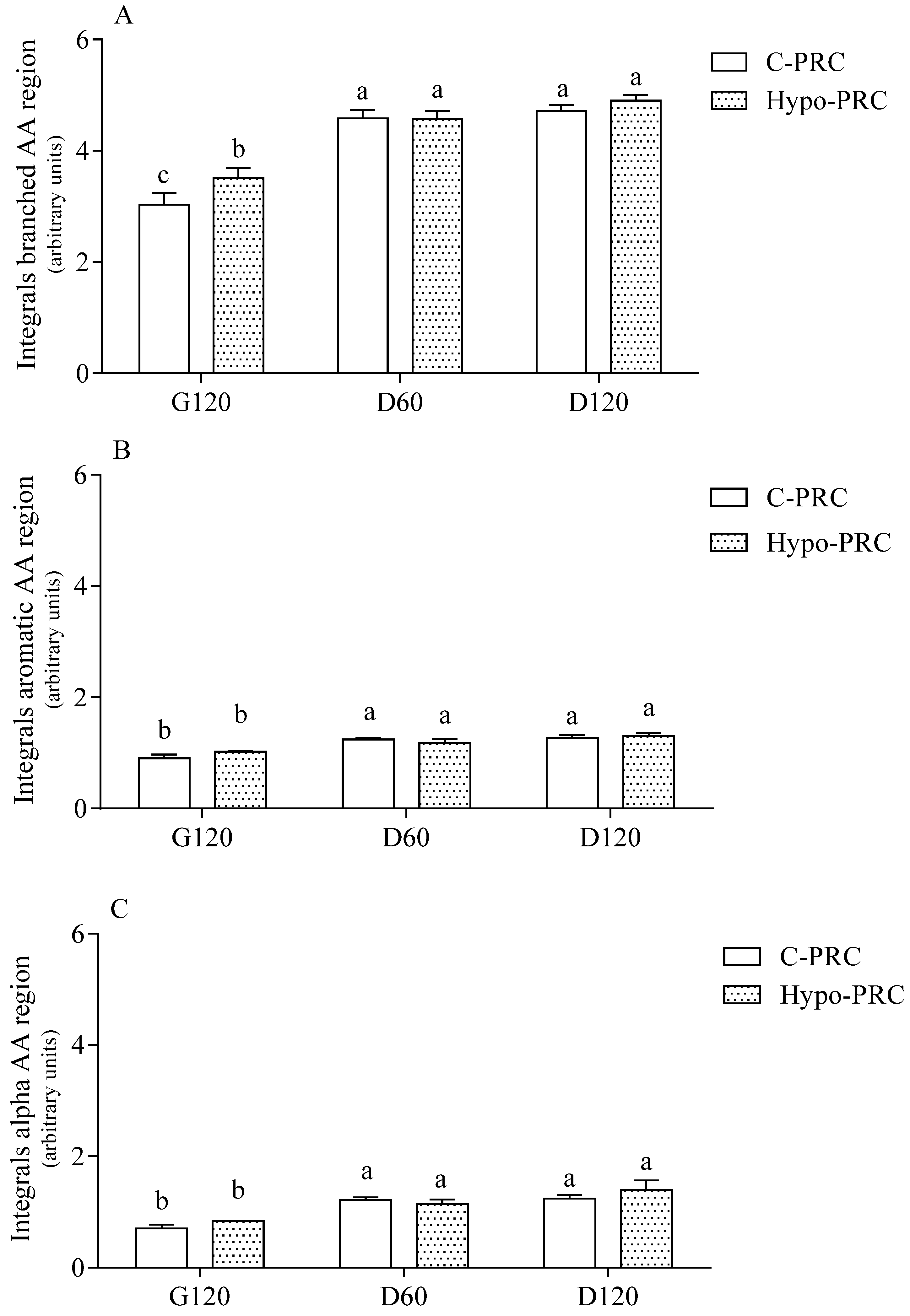
2.3. HR-NMR Spectroscopy
2.4. Peptide Formation
2.5. Bioinformatic Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Parmigiano Reggiano Cheese (PRC)
4.3. In Vitro Digestion
4.4. Fatty Acid Bioaccessibility
4.5. Protein Bioaccessibility
4.6. HR-NMR Spectroscopy
4.7. Bioactive Peptide Release and Identification
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Cai, W.; Lang, M.; Jiang, X.; Yu, Q.; Zhou, C.; Zou, W.; Zhang, X.; Lei, J. Correlation among high salt intake, blood pressure variability, and target organ damage in patients with essential hypertension: Study protocol clinical trial (SPIRIT compliant). Medicine 2020, 99, e19548. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Hibino, M.; Kobeissi, E.; Aune, D. Blood pressure, hypertension and the risk of sudden cardiac death: A systematic review and meta-analysis of cohort studies. Eur. J. Epidemiol. 2020, 35, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Thomas-Danguin, T.; Guichard, E.; Salles, C. Cross-modal interaction as a strategy to enhance salty taste and to maintain liking of low-salt food: A review. Food Funct. 2019, 10, 5269–5281. [Google Scholar] [CrossRef] [PubMed]
- Inguglia, E.S.; Zhang, Z.; Tiwari, B.K.; Kerry, J.P.; Burgess, C.M. Salt reduction strategies in processed meat products—A review. Trends Food Sci. Technol. 2017, 59, 70–78. [Google Scholar] [CrossRef]
- Ali, S.H.; Luo, R.; Li, Y.; Liu, X.; Tang, C.; Zhang, P. Application of Mobile Health Technologies Aimed at Salt Reduction: Systematic Review. JMIR Mhealth Uhealth 2019, 7, e13250. [Google Scholar] [CrossRef]
- Solieri, L.; Bianchi, A.; Giudici, P. Inventory of non starter lactic acid bacteria from ripened Parmigiano Reggiano cheese as assessed by a culture dependent multiphasic approach. Syst. Appl. Microbiol. 2012, 35, 270–277. [Google Scholar] [CrossRef]
- Bottesini, C.; Paolella, S.; Lambertini, F.; Galaverna, G.; Tedeschi, T.; Dossena, A.; Marchelli, R.; Sforza, S. Antioxidant capacity of water soluble extracts from Parmigiano-Reggiano cheese. Int. J. Food Sci. Nutr. 2013, 64, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Mazorra-Manzano, M.A.; Robles-Porchas, G.R.; González-Velázquez, D.A.; Torres-Llanez, M.J.; Martínez-Porchas, M.; García-Sifuentes, C.O.; González-Córdova, A.F.; Vallejo-Córdoba, B. Cheese Whey Fermentation by Its Native Microbiota: Proteolysis and Bioactive Peptides Release with ACE-Inhibitory Activity. Fermentation 2020, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Solieri, L.; Baldaccini, A.; Martini, S.; Bianchi, A.; Pizzamiglio, V.; Tagliazucchi, D. Peptide Profiling and Biological Activities of 12-Month Ripened Parmigiano Reggiano Cheese. Biology 2020, 9, 170. [Google Scholar] [CrossRef]
- Loffi, C.; Bortolazzo, E.; Garavaldi, A.; Musi, V.; Reverberi, P.; Galaverna, G.; Sforza, S.; Tedeschi, T. Reduction in the Brining Time in Parmigiano Reggiano Cheese Production Minimally Affects Proteolysis, with No Effect on Sensory Properties. Foods 2021, 10, 770. [Google Scholar] [CrossRef]
- Santos, D.I.; Saraiva, J.M.A.; Vicente, A.A.; Moldão-Martins, M. 2—Methods for determining bioavailability and bioaccessibility of bioactive compounds and nutrients. In Innovative Thermal and Non-Thermal Processing, Bioaccessibility and Bioavailability of Nutrients and Bioactive Compounds; Barba, F.J., Saraiva, J.M.A., Cravotto, G., Lorenzo, J.M., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 23–54. [Google Scholar] [CrossRef]
- Bordoni, A.; Laghi, L.; Babini, E.; Di Nunzio, M.; Picone, G.; Ciampa, A.; Valli, V.; Danesi, F.; Capozzi, F. The foodomics approach for the evaluation of protein bioaccessibility in processed meat upon in vitro digestion. Electrophoresis 2014, 35, 1607–1614. [Google Scholar] [CrossRef]
- Bordoni, A.; Picone, G.; Babini, E.; Vignali, M.; Danesi, F.; Valli, V.; Di Nunzio, M.; Laghi, L.; Capozzi, F. NMR comparison of in vitro digestion of Parmigiano Reggiano cheese aged 15 and 30 months. Magn. Reson. Chem. 2011, 49, S61–S70. [Google Scholar] [CrossRef]
- Minekus, M.; Marie, A.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in-vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malacarne, M.; Summer, A.; Franceschi, P.; Formaggioni, P.; Pecorari, M.; Panari, G.; Mariani, P. Free fatty acid profile of Parmigiano–Reggiano cheese throughout ripening: Comparison between the inner and outer regions of the wheel. Int. Dairy J. 2009, 19, 637–641. [Google Scholar] [CrossRef]
- Bennett, J.E.; Stevens, G.A.; Mathers, C.D.; Bonita, R.; Rehm, J.; Kruk, M.E.; Riley, L.M.; Dain, K.; Kengne, A.P.; Chalkidou, K.; et al. NCD Countdown 2030: Worldwide trends in non-communicable disease mortality and progress towards Sustainable Development Goal target 3.4. Lancet 2018, 392, 1072–1088. [Google Scholar] [CrossRef] [Green Version]
- Filippini, T.; Malavolti, M.; Whelton, P.K.; Naska, A.; Orsini, N.; Vinceti, M. Blood Pressure Effects of Sodium Reduction: Dose-Response Meta-Analysis of Experimental Studies. Circulation 2021, 143, 1542–1567. [Google Scholar] [CrossRef]
- Bhat, Z.F.; Morton, J.D.; Mason, S.L.; Bekhit, A.E.A. The application of pulsed electric field as a sodium reducing strategy for meat products. Food Chem. 2020, 306, 125622. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yan, Y.; Yu, Z.; Wu, T.; Bennett, L.E. Effects of high pressure processing on microbial, textural and sensory properties of low-salt emulsified beef sausage. Food Control 2022, 133, 108596. [Google Scholar] [CrossRef]
- Carbonell-Capella, J.M.; Buniowska, M.; Barba, F.J.; Esteve, M.J.; Frígola, A. Analytical Methods for Determining Bioavailability and Bioaccessibility of Bioactive Compounds from Fruits and Vegetables: A Review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 155–171. [Google Scholar] [CrossRef]
- Ferranti, P.; Nitride, C.; Nicolai, M.A.; Mamone, G.; Picariello, G.; Bordoni, A.; Valli, V.; Di Nunzio, M.; Babini, E.; Marcolini, E.; et al. In vitro digestion of Bresaola proteins and release of potential bioactive peptides. Food Res. Int. 2014, 63, 157–169. [Google Scholar] [CrossRef]
- Salaberría, F.; Marzocchi, S.; Bortolazzo, E.; Carrín, M.E.; Caboni, M.F. Study of the Effect of NaCl on Lipolysis in Parmigiano Reggiano Cheese. ACS Food Sci. Technol. 2021, 1, 54–59. [Google Scholar] [CrossRef]
- Urbinati, E.; Di Nunzio, M.; Picone, G.; Chiarello, E.; Bordoni, A.; Capozzi, F. The Effect of Balsamic Vinegar Dressing on Protein and Carbohydrate Digestibility is Dependent on the Food Matrix. Foods 2021, 10, 411. [Google Scholar] [CrossRef]
- Egger, L.; Schlegel, P.; Baumann, C.; Stoffers, H.; Guggisberg, D.; Brügger, C.; Dürr, D.; Stoll, P.; Vergères, G.; Portmann, R. Physiological comparability of the harmonized INFOGEST in vitro digestion method to in vivo pig digestion. Food Res. Int. 2017, 102, 567–574. [Google Scholar] [CrossRef]
- Deng, R.; Seimys, A.; Mars, M.; Janssen, A.E.M.; Smeets, P.A.M. Monitoring pH and whey protein digestion by TD-NMR and MRI in a novel semi-dynamic in vitro gastric simulator (MR-GAS). Food Hydrocoll. 2022, 125, 107393. [Google Scholar] [CrossRef]
- Martini, S.; Conte, A.; Tagliazucchi, D. Effect of ripening and in vitro digestion on the evolution and fate of bioactive peptides in Parmigiano-Reggiano cheese. Int. Dairy J. 2020, 105, 104668. [Google Scholar] [CrossRef]
- Qi, W.; Su, R.X.; He, Z.M.; Zhang, Y.B.; Jin, F.M. Pepsin-Induced Changes in the Size and Molecular Weight Distribution of Bovine Casein During Enzymatic Hydrolysis. J. Dairy Sci. 2007, 90, 5004–5011. [Google Scholar] [CrossRef] [Green Version]
- Sams, L.; Amara Ep Douzi, S.; Puppo, R.; Lebrun, R.; Paume, J.; Giallo, J.; Carrière, F. Characterization of pepsin from rabbit gastric extract, its action on β-casein and the effects of lipids on proteolysis. Food Funct. 2018, 9, 5975–5988. [Google Scholar] [CrossRef] [PubMed]
- Tagliazucchi, D.; Helal, A.; Verzelloni, E.; Bellesia, A.; Conte, A. Composition and properties of peptides that survive standardised in vitro gastro-pancreatic digestion of bovine milk. Int. Dairy J. 2016, 61, 196–204. [Google Scholar] [CrossRef]
- Liu, H.; Li, Q.; Zhao, D.; Zhang, M.; Jiang, S.; Li, C. Changes in the structure and digestibility of myoglobin treated with sodium chloride. Food Chem. 2021, 363, 130284. [Google Scholar] [CrossRef]
- Basiricò, L.; Catalani, E.; Morera, P.; Cattaneo, S.; Stuknytė, M.; Bernabucci, U.; De Noni, I.; Nardone, A. Release of angiotensin converting enzyme-inhibitor peptides during in vitro gastrointestinal digestion of Parmigiano Reggiano PDO cheese and their absorption through an in vitro model of intestinal epithelium. J. Dairy Sci. 2015, 98, 7595–7601. [Google Scholar] [CrossRef] [Green Version]
- Bottari, B.; Quartieri, A.; Prandi, B.; Raimondi, S.; Leonardi, A.; Rossi, M.; Ulrici, A.; Gatti, M.; Sforza, S.; Nocetti, M.; et al. Characterization of the peptide fraction from digested Parmigiano Reggiano cheese and its effect on growth of lactobacilli and bifidobacteria. Int. J. Food Microbiol. 2017, 255, 32–41. [Google Scholar] [CrossRef]
- Rai, A.K.; Sanjukta, S.; Jeyaram, K. Production of angiotensin I converting enzyme inhibitory (ACE-I) peptides during milk fermentation and their role in reducing hypertension. Crit. Rev. Food Sci. Nutr. 2017, 57, 2789–2800. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Ghini, V.; Di Nunzio, M.; Tenori, L.; Valli, V.; Danesi, F.; Capozzi, F.; Luchinat, C.; Bordoni, A. Evidence of a DHA Signature in the Lipidome and Metabolome of Human Hepatocytes. Int. J. Mol. Sci. 2017, 18, 359. [Google Scholar] [CrossRef] [Green Version]
- Bub, A.; Malpuech-Brugère, C.; Orfila, C.; Amat, J.; Arianna, A.; Blot, A.; Di Nunzio, M.; Holmes, M.; Kertész, Z.; Marshall, L.; et al. A Dietary Intervention of Bioactive Enriched Foods Aimed at Adults at Risk of Metabolic Syndrome: Protocol and Results from PATHWAY-27 Pilot Study. Nutrients 2019, 11, 1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Nunzio, M.; Betoret, E.; Taccari, A.; Dalla Rosa, M.; Bordoni, A. Impact of processing on the nutritional and functional value of mandarin juice. J. Sci. Food Agric. 2020, 100, 4558–4564. [Google Scholar] [CrossRef]
- Spellman, D.; McEvoy, E.; O’Cuinn, G.; FitzGerald, R.J. Proteinase and exopeptidase hydrolysis of whey protein: Comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy J. 2003, 13, 447–453. [Google Scholar] [CrossRef]
- Valli, V.; Danesi, F.; Gianotti, A.; Di Nunzio, M.; Saa, D.L.T.; Bordoni, A. Antioxidative and anti-inflammatory effect of in vitro digested cookies baked using different types of flours and fermentation methods. Food Res. Int. 2016, 88, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Di Nunzio, M.; Toselli, M.; Verardo, V.; Caboni, M.F.; Bordoni, A. Counteraction of oxidative damage by pomegranate juice: Influence of the cultivar. J. Sci. Food Agric. 2013, 93, 3565–3573. [Google Scholar] [CrossRef]
- Di Nunzio, M.; Picone, G.; Pasini, F.; Chiarello, E.; Caboni, M.F.; Capozzi, F.; Gianotti, A.; Bordoni, A. Olive oil by-product as functional ingredient in bakery products. Influence of processing and evaluation of biological effects. Food Res. Int. 2020, 131, 108940. [Google Scholar] [CrossRef] [Green Version]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Chaudhary, K.; Sharma, M.; Nagpal, G.; Chauhan, J.S.; Singh, S.; Gautam, A.; Raghava, G.P.S. AHTPDB: A comprehensive platform for analysis and presentation of antihypertensive peptides. Nucleic Acids Res. 2014, 43, D956–D962. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| G120 | D60 | D120 | ||||
|---|---|---|---|---|---|---|
| C-PRC | Hypo-PRC | C-PRC | Hypo-PRC | C-PRC | Hypo-PRC | |
| C8:0 | 1.53 ± 0.58 b | 1.87 ± 0.12 b | 11.70 ± 3.73 ab | 1.08 ± 1.53 b | 16.32 ± 6.09 a | 16.69 ± 1.82 a |
| C10:0 | 1.24 ± 0.34 c | 2.05 ± 0.16 c | 22.18 ± 3.55 ab | 16.76 ± 6.50 b | 31.36 ± 4.48 a | 32.10 ± 3.63 a |
| C12:0 | 1.55 ± 0.17 c | 2.51 ± 0.11 c | 27.32 ± 4.78 b | 35.60 ± 3.62 ab | 42.51 ± 4.30 a | 38.21 ± 1.20 a |
| C14:0 | 1.48 ± 0.23 c | 3.00 ± 0.05 c | 26.30 ± 4.92 b | 45.17 ± 1.24 a | 43.99 ± 9.63 ab | 34.19 ± 5.46 ab |
| C16:0 | 1.52 ± 0.38 b | 2.89 ± 0.02 b | 22.32 ± 4.81 ab | 43.83 ± 5.05 a | 39.80 ± 11.00 a | 28.24 ± 4.79 a |
| C16:1 n-7 | 0.00 ± 0.00 c | 1.32 ± 1.86 c | 33.92 ± 5.47 b | 49.02 ± 3.33 ab | 53.47 ± 9.05 a | 44.45 ± 4.37 ab |
| C18:0 | 1.84 ± 0.37 c | 2.88 ± 0.08 c | 20.68 ± 5.26 bc | 43.23 ± 5.89 a | 37.24 ± 10.12 a | 25.30 ± 5.6 ab |
| C18:1 n-9 | 0.96 ± 0.74 c | 2.03 ± 0.15 c | 27.99 ± 6.26 b | 45.29 ± 6.01 a | 47.79 ± 5.10 a | 47.86 ± 3.24 a |
| C18:2 n-6 | 0.53 ± 0.92 c | 1.07 ± 1.52 c | 34.01 ± 5.20 b | 47.58 ± 7.64 ab | 56.43 ± 9.48 a | 48.29 ± 4.37 ab |
| ΣSFA | 1.47 ± 0.30 b | 2.69 ± 0.05 b | 21.72 ± 4.69 a | 39.12 ± 2.00 a | 37.58 ± 8.64 a | 27.95 ± 4.27 a |
| ΣMUFA | 0.88 ± 0.68 c | 1.98 ± 0.00 c | 28.40 ± 6.24 b | 45.60 ± 5.76 a | 48.22 ± 5.38 a | 47.60 ± 2.69 a |
| ΣPUFA | 0.53 ± 0.92 c | 1.07 ± 1.52 c | 34.01 ± 5.20 b | 47.58 ± 7.64 ab | 56.43 ± 9.48 a | 48.29 ± 4.37 ab |
| Total | 1.28 ± 0.40 c | 2.45 ± 0.07 c | 23.95 ± 5.13 b | 41.13 ± 3.15 a | 40.97 ± 7.59 a | 33.90 ± 2.48 ab |
| Protein Source | Not digested a | G120 | D60 | D120 | ANOVA | ||||
|---|---|---|---|---|---|---|---|---|---|
| C-PRC | Hypo-PRC | C-PRC | Hypo-PRC | C-PRC | Hypo-PRC | C-PRC | Hypo-PRC | ||
| β-casein | 41.0 ± 0.0 c | 41.0 ± 0.0 c | 46.33 ± 2.52 c | 50.0 ± 2.0 bc | 66.0 ± 5.2 a | 60.0 ± 3.46 ab | 65.33 ± 9.45 a | 66.0 ± 2.0 a | p < 0.0001 |
| αS1-casein | 25.0 ± 0.0 a | 25.0 ± 0.0 a | 19.67 ± 0.58 a | 21.67 ± 1.15 a | 25.0 ± 3.61 a | 23.3 ± 2.61 a | 21.67 ± 3.79 a | 24.33 ± 3.06 a | p = 0.0992 |
| αS2-casein | 9.0 ± 0.0 a | 9.0 ± 0.0 a | 7.0 ± 1.0 ab | 7.33 ± 1.15 ab | 6.67 ± 0.58 b | 8.33 ± 0.58 ab | 7.33 ± 1.53 ab | 8.0 ± 0.0 ab | p = 0.0167 |
| κ-casein | 3.0 ± 0.0 c | 3.0 ± 0.0 c | 9.33 ± 0.58 a | 9.67 ± 1.15 a | 3.33 ± 0.58 c | 6.0 ± 1.0 b | 3.33 ± 1.15 c | 3.67 ± 0.58 c | p < 0.0001 |
| β-LG(A and B isoforms) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 1.0 ± 0.0 a | 0.67 ± 0.58 ab | 1.0 ± 0.0 a | 0.67 ± 0.58 ab | p = 0.0004 |
| Total | 78.0 ± 0.0 c | 78.0 ± 0.0 c | 82.33 ± 2.31 bc | 88.67 ± 4.16 ABC | 102.0 ± 8.72 a | 98.0 ± 6.56 ab | 98.67 ± 12.50 ab | 102.67 ± 4.93 a | p = 0.0002 |
| Peptide Sequence | Protein Source | Not Digested | G120 | D60 | D120 | Reported Activity (uM IC50) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| C-PRC | Hypo-PRC | C-PRC | Hypo-PRC | C-PRC | Hypo-PRC | C-PRC | Hypo-PRC | |||
| LHLPLP | β-CN (133–138) | 62 ± 3.8 a | 64.7 ± 7.8 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | ACE inhibition (5.5) |
| RPKH | αS1-CN (1–4) | 0.9 ± 0.1 a | 0.9 ± 0.1 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | ACE inhibition (>1863) |
| YKVPQL | αS1-CN (104–109) | 0.6 ± 0.1 a | 0.6 ± 0.1 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | ACE inhibition (>22) |
| NLHLPLPLL | β-CN (132–140) | 25.2 ± 1.9 a | 25.5 ± 2.7 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | ACE inhibition (15) |
| RELEEL | β-CN (1–6) | 1.5 ± 0.4 ab | 2.0 ± 0.5 a | 0.9 ± 0.1 b | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | ACE inhibition (1000) |
| LLYQEPVLGPVRGPFPIIV | β-CN (191–209) | 2.6 ± 0.3 a | 2.7 ± 0.3 a | 0.3 ± 0.0 b | 0.3 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | ACE inhibition andimmunomodulating (nr) |
| RPKHPIKHQGLPQEVLNENLLRF | αS1-CN (1–23) | 4.7 ± 1.3 a | 5.22 ± 1.2 a | 0.3 ± 0.1 b | 0.3 ± 0.1 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | Antibacterial (nr) |
| AYFYPEL | αS1-CN (143–149) | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.26 ± 0.0 a | 0.22 ± 0.04 b | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | ACE inhibition (6.6) |
| DAYPSGAW | αS1-CN (157–164) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.05 ± 0.01 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | ACE inhibition (98) |
| LKKISQRYQKFALPQYLKT | αS2-CN (164–182) | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 1.5 ± 0.0 a | 1.4 ± 0.06 b | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | Hemolytic (nr) |
| PLW | αS1-CN (196–199) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.1 ± 0.01 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | ACE inhibition (36) |
| VENLHLPLPLL | β-CN (129–139) | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.4 ± 0.01 a | 0.35 ± 0.03 b | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | Anticancer (nr) |
| VYQHQKAMKPWIQPKTKVIPYVRYL | αS2-CN (183–207) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 2.3 ± 0.4 a | 2.4 ± 0.1 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | Antibacterial (nr) |
| YQKFPQY | αS2-CN (89–95) | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.8 ± 0.08 a | 0.7 ± 0.0 b | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | ACE inhibition (20.1) |
| TPEVDDEALEK | β-Lg (125–135) | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.4 ± 0.05 a | 0.2 ± 0.0 b | 0.0 ± 0.0 c | 0.0 ± 0.0 c | DPP-IV inhibition (319.5) |
| VPYPQ | β-CN (177–181) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.1 ± 0.01 a | 0.1 ± 0.01 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | Antioxidant (nr) |
| FYPEL | αS1-CN (145–149) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.7 ± 0.0 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | Antioxidant (nr) |
| HLPLP | β-CN (134–138) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.5 ± 0.2 b | 1.7 ± 0.8 a | 0.4 ± 0.08 b | 0.4 ± 0.1 b | ACE inhibition (41) |
| EMPFPK | β-CN (108–113) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 2.5 ± 0.4 a | 2.8 ± 0.4 a | 2.7 ± 0.1 a | 2.4 ± 0.1 a | ACE inhibition (423) |
| HLPLPL | β-CN (134–139) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.5 ± 0.2 b | 1.7 ± 0.8 a | 0.4 ± 0.08 b | 0.4 ± 0.1 b | Antiamnestic (10) |
| KEDVPSE | αS1-CN (83–89) | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.06 ± 0.0 b | 0.1 ± 0.04 a | 0.1 ± 0.0 c | 0.1 ± 0.0 c | ACE inhibition (41) |
| LPLPL | β-CN (135–139) | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.8 ± 0.3 b | 1.8 ± 0.3 a | 1.1 ± 0.2 b | 1.1 ± 0.3 b | DPP-IV inhibition (325) |
| LPYP | k-CN (56–59) | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 1.1 ± 0.3 ab | 1.5 ± 0.1 a | 1.1 ± 0.2 ab | 0.9 ± 0.2 b | ACE inhibition (5) |
| NIPPLTQTPV | β-CN (73–82) | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.9 ± 0.2 ab | 0.5 ± 0.09 bc | 1.3 ± 0.6 a | 0.9 ± 0.2 ab | ACE inhibition (173) |
| PQNIPPL | β-CN (71–77) | 0.0 ± 0.0 d | 0.0 ± 0.0 d | 0.0 ± 0.0 d | 0.0 ± 0.0 d | 0.02 ± 0.0 c | 0.05 ± 0.01 b | 0.025 ± 0.0 c | 0.1 ± 0.01 a | DPP-IV inhibition (1500) |
| VVPPFLQPE | β-CN (83–91) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 1.5 ± 0.1 a | 1.6 ± 0.6 a | 1.5 ± 0.5 a | 2.0 ± 0.2 a | Antioxidant (nr) |
| VYPFPGPI | β-CN (59–66) | 0.0±0.0 d | 0.0 ± 0.0 d | 0.0 ± 0.0 d | 0.0 ± 0.0 d | 2.9 ± 0.2 c | 4.0 ± 0.2 b | 4.2 ± 0.6 b | 5.2 ± 0.4 a | Antiamnestic (110) |
| YPEL | αS1–CN (146-149) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 1.4 ± 0.2 a | 1.5 ± 0.2 a | 1.2 ± 0.1 a | 1.3 ± 0.1 a | Antioxidant (nr) |
| YPFPGPI | β-CN (60–66) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.8 ± 0.06 a | 1.1 ± 0.4 a | 0.9 ± 0.1 a | 0.8 ± 0.007 a | ACE inhibition (500) |
| YPVEPF | β-CN (114–119) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 7.6 ± 1.0 a | 9.0 ± 1.8 a | 7.7 ± 0.8 a | 8.7 ± 0.8 a | Opioid (nr) |
| LHLPLPL | β-CN (133–139) | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.2 ± 0.09 b | 0.3 ± 0.03 a | ACE inhibition (432.7) |
| PGPIPN | β-CN (63–68) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.09 ± 0.0 a | Immunomodulating (nr) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Nunzio, M.; Loffi, C.; Chiarello, E.; Dellafiora, L.; Picone, G.; Antonelli, G.; Di Gregorio, C.; Capozzi, F.; Tedeschi, T.; Galaverna, G.; et al. Impact of a Shorter Brine Soaking Time on Nutrient Bioaccessibility and Peptide Formation in 30-Months-Ripened Parmigiano Reggiano Cheese. Molecules 2022, 27, 664. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27030664
Di Nunzio M, Loffi C, Chiarello E, Dellafiora L, Picone G, Antonelli G, Di Gregorio C, Capozzi F, Tedeschi T, Galaverna G, et al. Impact of a Shorter Brine Soaking Time on Nutrient Bioaccessibility and Peptide Formation in 30-Months-Ripened Parmigiano Reggiano Cheese. Molecules. 2022; 27(3):664. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27030664
Chicago/Turabian StyleDi Nunzio, Mattia, Cecilia Loffi, Elena Chiarello, Luca Dellafiora, Gianfranco Picone, Giorgia Antonelli, Clarissa Di Gregorio, Francesco Capozzi, Tullia Tedeschi, Gianni Galaverna, and et al. 2022. "Impact of a Shorter Brine Soaking Time on Nutrient Bioaccessibility and Peptide Formation in 30-Months-Ripened Parmigiano Reggiano Cheese" Molecules 27, no. 3: 664. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27030664