
Antidiabetic Potential of Volatile Cinnamon Oil: A Review and Exploration of Mechanisms Using In Silico Molecular Docking Simulations

Abstract
:1. Introduction
2. Cinnamon Essential Oil and Its Constituents
3. Literature Review of Antidiabetic Properties of Cinnamon Essential Oil and Its Constituents
4. In Silico Docking Models
- Agonistic: behaving like the natural ligand, agonist molecules bind to the receptor and trigger signaling. When a small molecule is simulated to bind in the same active site as a drug or mimetic with known biological effects, it is considered an agonist.
- Antagonistic: inhibiting the effect of the natural ligand or agonist by blocking the active site of the enzyme (competitive agonist), by binding elsewhere on the enzyme and altering the biological function (noncompetitive antagonist), or by covalently altering the binding site (irreversible agonist). A small molecule binding in this manner would inhibit the activity of the enzyme and alter downstream pathway outcomes.
5. Materials and Methods
Molecular Docking
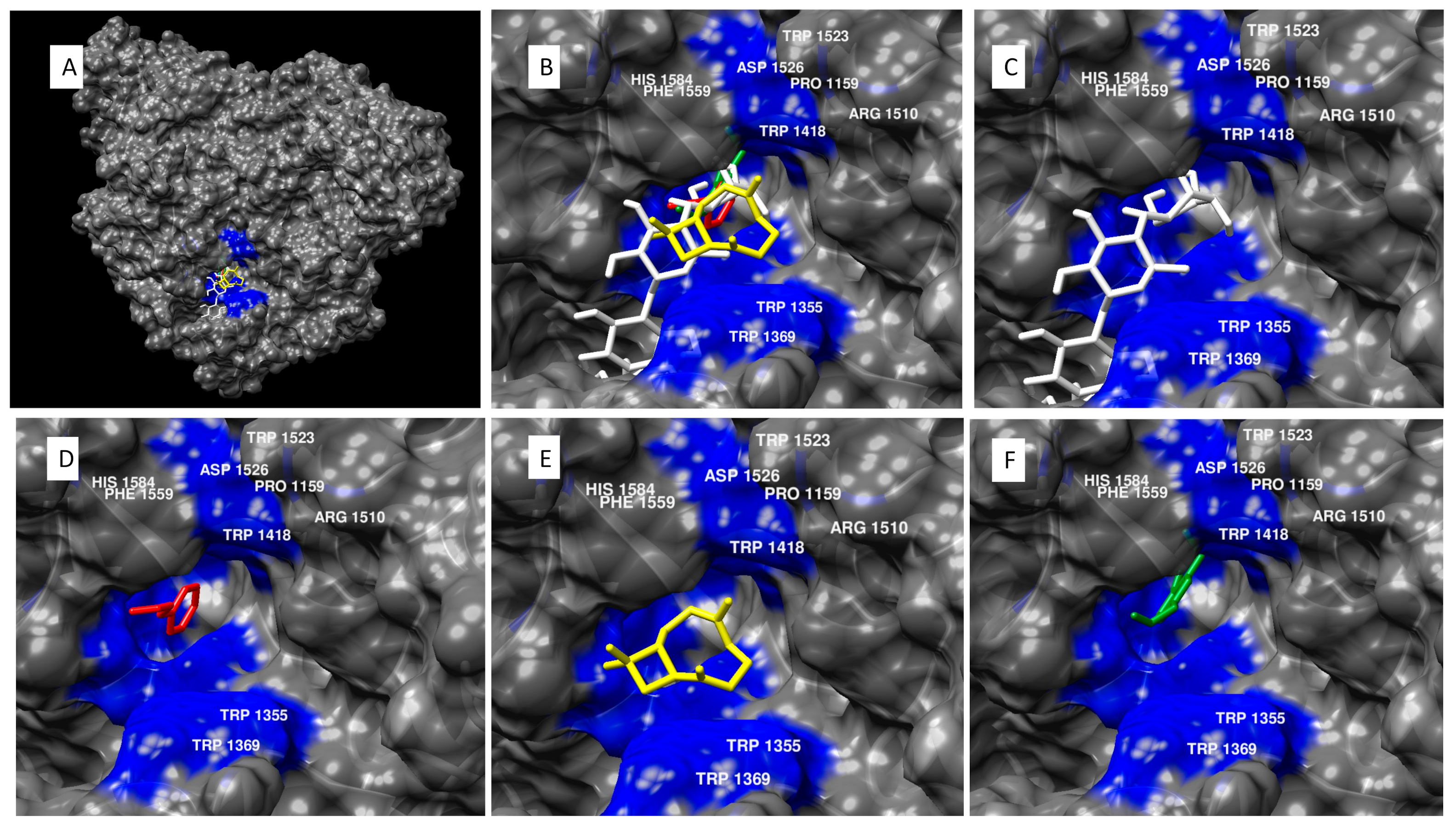
6. Modeling Results and Discussion
7. Limitations
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Santos-Sánchez, N.F.; Salas-Coronado, R.; Hernández-Carlos, B.; Villanueva-Cañongo, C. Shikimic acid pathway in biosynthesis of phenolic compounds. In Plant Physiological Aspects of Phenolic Compounds; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Dewick, P. The shikimate pathway: Aromatic amino acids and phenylpropanoids. In Medicinal Natural Products; John Wiley & Sons: Chichester, UK, 2009. [Google Scholar]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: A historical perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Sun, P.; Schuurink, R.C.; Caissard, J.-C.; Hugueney, P.; Baudino, S. My Way: Noncanonical Biosynthesis Pathways for Plant Volatiles. Trends Plant Sci. 2016, 21, 884–894. [Google Scholar] [CrossRef]
- Aharoni, A.; Jongsma, M.A.; Bouwmeester, H.J. Volatile science? Metabolic engineering of terpenoids in plants. Trends Plant Sci. 2005, 10, 594–602. [Google Scholar] [CrossRef]
- Berraaouan, A.; Abid, S.; Bnouham, M. Antidiabetic oils. Curr. Diabetes Rev. 2013, 9, 499–505. [Google Scholar] [CrossRef]
- Ping, H.; Zhang, G.; Ren, G. Antidiabetic effects of cinnamon oil in diabetic KK-Ay mice. Food Chem. Toxicol. 2010, 48, 2344–2349. [Google Scholar] [CrossRef]
- Bakirel, T.; Bakirel, U.; Keleş, O.U.; Ulgen, S.G.; Yardibi, H. In vivo assessment of antidiabetic and antioxidant activities of rosemary (Rosmarinus officinalis) in alloxan-diabetic rabbits. J. Ethnopharmacol. 2008, 116, 64–73. [Google Scholar] [CrossRef]
- El-Soud, N.A.; El-Laithy, N.; El-Saeed, G.; Wahby, M.S.; Khalil, M.; Morsy, F.; Shaffie, N. Antidiabetic activities of Foeniculum vulgare Mill. essential oil in streptozotocin-induced diabetic rats. Maced. J. Med. Sci. 2011, 4, 139–146. [Google Scholar]
- Verspohl, E.J.; Bauer, K.; Neddermann, E. Antidiabetic effect of Cinnamomum cassia and Cinnamomum zeylanicum In vivo and in vitro. Phytother. Res. 2005, 19, 203–206. [Google Scholar] [CrossRef]
- National Diabetes Statistics Report | Data & Statistics|Diabetes|CDC. Available online: https://www.cdc.gov/diabetes/data/statistics/statistics-report.html (accessed on 21 February 2020).
- Diabetes Infographics|Social Media|Press & Social Media|Diabetes|CDC. Available online: https://www.cdc.gov/diabetes/library/socialmedia/infographics.html (accessed on 21 February 2020).
- de Ruyck, J.; Brysbaert, G.; Blossey, R.; Lensink, M.F. Molecular docking as a popular tool in drug design, an In silico travel. Adv. Appl. Bioinform. Chem. 2016, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Barceloux, D.G. Cinnamon (Cinnamomum Species). Dis. A-Mon. 2009, 55, 327–335. [Google Scholar] [CrossRef]
- Felter, H.W.; Lloyd, J. Kings American Dispensatory, 3rd ed.; Eclectic Medical Publications: Cincinnati, OH, USA, 1898; Volume II. [Google Scholar]
- Moore, M. Specific Indications for Herbs in General Use, 2nd ed.; Southwest School of Botanical Medicine: Bisbee, ARI, USA, 1997; Available online: http://swsbm.henriettesherbal.com/ManualsMM/SpecIndic3.pdf (accessed on 29 November 2021).
- Scheid, V.; Bensky, D.; Ellis, A.; Barolet, R. Chinese Herbal Medicine: Formulas & Strategies, 2nd Revised Edition; Eastland Press: Vista, CA, USA, 2009; ISBN 978-0-939616-67-1. Available online: https://westminsterresearch.westminster.ac.uk/item/90z76/chinese-herbal-medicine-formulas-strategies-2nd-revised-edition (accessed on 29 November 2021).
- Hartwell, J.L. Plants used against cancer. A survey. [Continued]. Lloydia 1970, 33, 97–194. [Google Scholar] [PubMed]
- Abascal, K. The Medicinal Uses of Cinnamon. Integr. Med. A Clin. J. 2010, 9, 28–32. [Google Scholar]
- Baruah, A.; Nath, S.C. Systematics and diversities of Cinammomum species used as “cinnamon” spice in north-east India. J. Econ. Taxon. Bot. 2007, 31, 872–887. [Google Scholar]
- Anandaraj, M.; Devasahayam, B.; Krishnamoorthy, B.; Matthew, P.; Rema, J. Cinnamon (Extension Pamphlet); Indian Institute of Spices Research: Kozhikode, India, 2005; Available online: http://iisr.agropedias.iitk.ac.in/sites/default/files/cinnamon.pdf (accessed on 16 March 2020).
- Jeyaratnam, N.; Nour, A.H.; Kanthasamy, R.; Nour, A.H.; Yuvaraj, A.R.; Akindoyo, J.O. Essential oil from Cinnamomum cassia bark through hydrodistillation and advanced microwave assisted hydrodistillation. Ind. Crops Prod. 2016, 92, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Tisserand, R.; Young, R. Essential Oil Safety: A Guide for Health Care Professionals, 2nd ed.; Elsevier Ltd.: Edinburgh, UK, 2013; ISBN 978-0-443-06241-4. [Google Scholar]
- Issues in New Crops and New Uses; Janick, J.; Whipkey, A. (Eds.) ASHS Press: Alexandria, VA, USA, 2007; Available online: https://www.hort.purdue.edu/newcrop/ncnu07/pdfs/koroch338-341.pdf (accessed on 10 December 2021).
- PubChem Cassia Oil. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/6850781 (accessed on 10 December 2021).
- Averesch, N.J.H.; Krömer, J.O. Metabolic engineering of the shikimate pathway for production of aromatics and derived compounds—Present and future strain construction strategies. Front. Bioeng. Biotechnol. 2018, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- PubChem Cinnamaldehyde. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/637511 (accessed on 27 August 2019).
- Stevens, N. Cinnamon Bark Essential Oil and a Novel Essential Oil Blend as Potential Modulators of Glucose Metabolism. Ph.D. Thesis, University of Miami, Miami, FL, USA, 2020. Available online: https://scholarship.miami.edu/esploro/outputs/doctoral/Cinnamon-Bark-Essentialoil-and-a/991031454482202976 (accessed on 1 December 2021).
- PubChem Eugenol. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/3314 (accessed on 27 August 2019).
- PubChem Beta-Caryophyllene. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5281515 (accessed on 27 August 2019).
- Schmidt, E.; Jirovetz, L.; Buchbauer, G.; Eller, G.A.; Stoilova, I.; Krastanov, A.; Stoyanova, A.; Geissler, M. Composition and Antioxidant Activities of the Essential Oil of Cinnamon (Cinnamomum zeylanicum Blume) Leaves from Sri Lanka. J. Essent. Oil Bear. Plants 2006, 9, 170–182. [Google Scholar] [CrossRef]
- Jantan, I.B.; Karim Moharam, B.A.; Santhanam, J.; Jamal, J.A. Correlation Between Chemical Composition and Antifungal Activity of the Essential Oils of Eight Cinnamomum. Species. Pharm. Biol. 2008, 46, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-T.; Chang, W.-L.; Hsu, J.-C.; Shih, Y.; Chou, S.-T. Chemical composition and tyrosinase inhibitory activity of Cinnamomum cassia essential oil. Bot. Stud. 2013, 54, 10. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-S. Chemical Composition of Cinnamomum cassia Leaf Oils and Suppression Effect of Cinnamyl Alcohol on Nitric Oxide Synthase. J. Korean Soc. Appl. Biol. Chem. 2009, 52, 480–485. [Google Scholar] [CrossRef]
- Mohammed, K.A.A.; Ahmed, H.M.S.; Sharaf, H.A.; El-Nekeety, A.A.; Abdel-Aziem, S.H.; Mehaya, F.M.; Abdel-Wahhab, M.A. Encapsulation of cinnamon oil in whey protein counteracts the disturbances in biochemical parameters, gene expression, and histological picture of the liver and pancreas of diabetic rats. Env. Sci. Pollut. Res. Int. 2020, 27, 2829–2843. [Google Scholar] [CrossRef]
- Mohammed, K.A.A. Efficiency of encapsulated cinnamon oil in the treatment of diabetes in rats. CU Theses 2020. Available online: http://erepository.cu.edu.eg/index.php/cutheses/article/view/8618 (accessed on 1 December 2021).
- Mishra, A.; Bhatti, R.; Singh, A.; Singh Ishar, M.P. Ameliorative effect of the cinnamon oil from Cinnamomum zeylanicum upon early stage diabetic nephropathy. Planta Med. 2010, 76, 412–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budiastuti, B.; Safitri, Y.; Plumeriastuti, H.; Srianto, P.; Effendi, M.; Helmi, M. Effect of Cinnamon (Cinnamomum burmannii) Bark Oil on Testicular Histopathology in Streptozotocin Induced Diabetic Wistar Male Rats. J. Glob. Pharm. Technol. 2020, 12, 901–907. [Google Scholar]
- Kusmardi, K.; Tedjo, A.; Fadilah, F.; Arsianti, A.; Paramita, R. Identification by Docking Simulation and in vivo Effect of Essential Oil from Cinnamommum burmannii as Anti-obesity with Leptin Receptor in the Olfactory System of Mice Balb C. Pharmacogn. J. 2018, 10, 875–879. [Google Scholar] [CrossRef] [Green Version]
- Zari, T.A.; Al-Logmani, A.S. Long–term effects of Cinnamomum zeylanicum Blume oil on some physiological parameters in streptozotocin-diabetic and non-diabetic rats. Boletín Latinoam. Y Caribe Plantas Med. Y Aromáticas 2009, 8, 266–274. [Google Scholar]
- Bhatti, R.; Kaur, S.; Singh, J.; Ishar, M. Ameliorative effect of volatile oil from Cinnamomum zeylanicum on hyperalgesia in alloxan diabetic rats. Can. J. Pure Appl. Sci. 2009, 3, 887–895. [Google Scholar]
- Li, J.-E.; Futawaka, K.; Yamamoto, H.; Kasahara, M.; Tagami, T.; Liu, T.-H.; Moriyama, K. Cinnamaldehyde Contributes to Insulin Sensitivity by Activating PPARδ, PPARγ, and RXR. Am. J. Chin. Med. 2015, 43, 879–892. [Google Scholar] [CrossRef]
- Camacho, S.; Michlig, S.; de Senarclens-Bezençon, C.; Meylan, J.; Meystre, J.; Pezzoli, M.; Markram, H.; le Coutre, J. Anti-Obesity and Anti-Hyperglycemic Effects of Cinnamaldehyde via altered Ghrelin Secretion and Functional impact on Food Intake and Gastric Emptying. Sci. Rep. 2015, 5, 7919. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Yang, Y.; Wang, D.; Yang, Q.; Wan, J.; Liu, S.; Zhou, P.; Yang, Y. Cinnamaldehyde Ameliorates Vascular Dysfunction in Diabetic Mice by Activating Nrf2. Am. J. Hypertens. 2020, 33, 610–619. [Google Scholar] [CrossRef]
- Anand, P.; Murali, K.Y.; Tandon, V.; Murthy, P.S.; Chandra, R. Insulinotropic effect of cinnamaldehyde on transcriptional regulation of pyruvate kinase, phosphoenolpyruvate carboxykinase, and GLUT4 translocation in experimental diabetic rats. Chem. Biol. Interact. 2010, 186, 72–81. [Google Scholar] [CrossRef]
- Hosni, A.A.; Abdel-Moneim, A.A.; Abdel-Reheim, E.S.; Mohamed, S.M.; Helmy, H. Cinnamaldehyde potentially attenuates gestational hyperglycemia in rats through modulation of PPARγ, proinflammatory cytokines and oxidative stress. Biomed. Pharm. 2017, 88, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Hosni, A.; El-Twab, S.A.; Abdul-Hamid, M.; Prinsen, E.; AbdElgawad, H.; Abdel-Moneim, A.; Beemster, G.T.S. Cinnamaldehyde mitigates placental vascular dysfunction of gestational diabetes and protects from the associated fetal hypoxia by modulating placental angiogenesis, metabolic activity and oxidative stress. Pharm. Res. 2021, 105426. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wu, H.; Duan, M.; Liu, R.; Zhu, Q.; Zhang, K.; Wang, L. Cinnamaldehyde Improves Metabolic Functions in Streptozotocin-Induced Diabetic Mice by Regulating Gut Microbiota. Drug Des. Dev. 2021, 15, 2339–2355. [Google Scholar] [CrossRef]
- El-Bassossy, H.M.; Fahmy, A.; Badawy, D. Cinnamaldehyde protects from the hypertension associated with diabetes. Food Chem. Toxicol. 2011, 49, 3007–3012. [Google Scholar] [CrossRef] [PubMed]
- Jawale, A.; Datusalia, A.K.; Bishnoi, M.; Sharma, S.S. Reversal of diabetes-induced behavioral and neurochemical deficits by cinnamaldehyde. Phytomedicine 2016, 23, 923–930. [Google Scholar] [CrossRef]
- Zhao, H.; Xie, Y.; Yang, Q.; Cao, Y.; Tu, H.; Cao, W.; Wang, S. Pharmacokinetic study of cinnamaldehyde in rats by GC–MS after oral and intravenous administration. J. Pharm. Biomed. Anal. 2014, 89, 150–157. [Google Scholar] [CrossRef]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced glycation end products and diabetic complications. Korean J. Physiol. Pharm. 2014, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Mnafgui, K.; Kaanich, F.; Derbali, A.; Hamden, K.; Derbali, F.; Slama, S.; Allouche, N.; Elfeki, A. Inhibition of key enzymes related to diabetes and hypertension by Eugenol in vitro and in alloxan-induced diabetic rats. Arch. Physiol. Biochem. 2013, 119, 225–233. [Google Scholar] [CrossRef]
- Srinivasan, S.; Sathish, G.; Jayanthi, M.; Muthukumaran, J.; Muruganathan, U.; Ramachandran, V. Ameliorating effect of eugenol on hyperglycemia by attenuating the key enzymes of glucose metabolism in streptozotocin-induced diabetic rats. Mol. Cell. Biochem. 2014, 385, 159–168. [Google Scholar] [CrossRef]
- Al-Trad, B.; Alkhateeb, H.; Alsmadi, W.; Al-Zoubi, M. Eugenol ameliorates insulin resistance, oxidative stress and inflammation in high fat-diet/streptozotocin-induced diabetic rat. Life Sci. 2019, 216, 183–188. [Google Scholar] [CrossRef]
- Jeong, K.J.; Kim, D.Y.; Quan, H.-Y.; Jo, H.K.; Kim, G.W.; Chung, S.H. Effects of eugenol on hepatic glucose production and AMPK signaling pathway in hepatocytes and C57BL/6J mice. Fitoterapia 2014, 93, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Garud, M.S.; Kulkarni, Y.A. Eugenol ameliorates renal damage in streptozotocin-induced diabetic rats. Flavour Fragr. J. 2017, 32, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Prasad, S.N.; Bharath, M.M.S. Muralidhara Neurorestorative effects of eugenol, a spice bioactive: Evidence in cell model and its efficacy as an intervention molecule to abrogate brain oxidative dysfunctions in the streptozotocin diabetic rat. Neurochem. Int. 2016, 95, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Shareghi Boroujeni, A.; Jalali Dehkordi, K.; Sharifi, G.; Taghian, F.; Mazaheri, Z. Effect of Nano-eugenol and Aerobic Exercise Against the Streptozotocin Toxicity and Inflammatory Mediators P38-MAPK, NPY, and A-Rα2A in the Dorsal Root Ganglia of Diabetic Rats. Iran. J. Toxicol. 2021, 15, 205–214. [Google Scholar] [CrossRef]
- Oroojan, A.A.; Chenani, N.; An’aam, M. Antioxidant Effects of Eugenol on Oxidative Stress Induced by Hydrogen Peroxide in Islets of Langerhans Isolated from Male Mouse. Int. J. Hepatol. 2020, 2020, e5890378. [Google Scholar] [CrossRef]
- Hamdin, C.D.; Utami, S.W.; Muliasari, H.; Prasedya, E.S.; Sudarma, I.M. Histological pattern on pancreas and liver of diabetic rats after treatment of eugenol isolated from leaves of Syzygium aromaticum. AIP Conf. Proc. 2019, 2199, 060004. [Google Scholar] [CrossRef]
- Mani, V.; Balraj, M.; Venktsan, G.; Soundrapandiyan, J.; Kasturi, R.; Danavel, A.; Babu, S. Molecular docking analysis of beta-caryophyllene with IRS-1, cSrc and Akt. Bioinformation 2021, 17, 916–920. [Google Scholar] [CrossRef]
- Aguilar-Ávila, D.S.; Flores-Soto, M.E.; Tapia-Vázquez, C.; Pastor-Zarandona, O.A.; López-Roa, R.I.; Viveros-Paredes, J.M. β-Caryophyllene, a Natural Sesquiterpene, Attenuates Neuropathic Pain and Depressive-Like Behavior in Experimental Diabetic Mice. J. Med. Food 2019, 22, 460–468. [Google Scholar] [CrossRef]
- Basha, R.H.; Sankaranarayanan, C. β-Caryophyllene, a natural sesquiterpene lactone attenuates hyperglycemia mediated oxidative and inflammatory stress in experimental diabetic rats. Chem. Biol. Interact. 2016, 245, 50–58. [Google Scholar] [CrossRef]
- Basha, R.H.; Sankaranarayanan, C. Protective role of β-caryophyllene, a sesquiterpene lactone on plasma and tissue glycoprotein components in streptozotocin-induced hyperglycemic rats. J. Acute Med. 2015, 5, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Basha, R.H.; Sankaranarayanan, C. β-Caryophyllene, a natural sesquiterpene, modulates carbohydrate metabolism in streptozotocin-induced diabetic rats. Acta Histochem. 2014, 116, 1469–1479. [Google Scholar] [CrossRef] [PubMed]
- Youssef, D.A.; El-Fayoumi, H.M.; Mahmoud, M.F. Beta-caryophyllene alleviates diet-induced neurobehavioral changes in rats: The role of CB2 and PPAR-γ receptors. Biomed. Pharmacother. 2019, 110, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, D.; Chen, Y.; Yang, M. β-Caryophyllene inhibits high glucose-induced oxidative stress, inflammation and extracellular matrix accumulation in mesangial cells. Int. Immunopharmacol. 2020, 84, 106556. [Google Scholar] [CrossRef] [PubMed]
- Geddo, F.; Antoniotti, S.; Querio, G.; Salaroglio, I.C.; Costamagna, C.; Riganti, C.; Gallo, M.P. Plant-Derived Trans-β-Caryophyllene Boosts Glucose Metabolism and ATP Synthesis in Skeletal Muscle Cells through Cannabinoid Type 2 Receptor Stimulation. Nutrients 2021, 13, 916. [Google Scholar] [CrossRef]
- Galic, S.; Hauser, C.; Kahn, B.B.; Haj, F.G.; Neel, B.G.; Tonks, N.K.; Tiganis, T. Coordinated regulation of insulin signaling by the protein tyrosine phosphatases PTP1B and TCPTP. Mol. Cell. Biol. 2005, 25, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Sweet, I.R.; Li, G.; Najafi, H.; Berner, D.; Matschinsky, F.M. Effect of a glucokinase inhibitor on energy production and insulin release in pancreatic islets. Am. J. Physiol. 1996, 271, E606–E625. [Google Scholar] [CrossRef]
- Matschinsky, F.M.; Zelent, B.; Doliba, N.; Li, C.; Vanderkooi, J.M.; Naji, A.; Sarabu, R.; Grimsby, J. Glucokinase activators for diabetes therapy. Diabetes Care 2011, 34, S236–S243. [Google Scholar] [CrossRef] [Green Version]
- Marini, C.; Salani, B.; Massollo, M.; Amaro, A.; Esposito, A.I.; Orengo, A.M.; Capitanio, S.; Emionite, L.; Riondato, M.; Bottoni, G.; et al. Direct inhibition of hexokinase activity by metformin at least partially impairs glucose metabolism and tumor growth in experimental breast cancer. Cell Cycle 2013, 12, 3490–3499. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zheng, M.; Wu, S.; Gao, S.; Yang, M.; Li, Z.; Min, Q.; Sun, W.; Chen, L.; Xiang, G.; et al. Benserazide, a dopadecarboxylase inhibitor, suppresses tumor growth by targeting hexokinase 2. J. Exp. Clin. Cancer Res. 2017, 36, 58. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, G.; Huang, C.; Greenblatt, M.F. UCSF Chimera--a visualization system for exploratory research and analysis—PubMed. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brayer, G.D.; Luo, Y.; Withers, S.G. The structure of human pancreatic α-amylase at 1.8 Å resolution and comparisons with related enzymes. Protein Sci. 1995, 4, 1730–1742. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Qin, X.; Cao, X.; Wang, L.; Bai, F.; Bai, G.; Shen, Y. Structural insight into substrate specificity of human intestinal maltase-glucoamylase. Protein Cell 2011, 2, 827–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheruvallath, Z.S.; Gwaltney, S.L.; Sabat, M.; Tang, M.; Wang, H.; Jennings, A.; Hosfield, D.; Lee, B.; Wu, Y.; Halkowycz, P.; et al. Discovery of potent and orally active 1,4-disubstituted indazoles as novel allosteric glucokinase activators. Bioorganic Med. Chem. Lett. 2017, 27, 2678–2682. [Google Scholar] [CrossRef] [PubMed]
- Groves, M.R.; Yao, Z.-J.; Roller, P.P.; Burke, T.R.; Barford, D. Structural basis for inhibition of the protein tyrosine phosphatase 1B by phosphotyrosine peptide mimetics. Biochemistry 1998, 37, 17773–17783. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.H.; Ferreira, J.C.; Nedyalkova, L.; Zhu, H.; Carrasco-López, C.; Kirmizialtin, S.; Rabeh, W.M. The catalytic inactivation of the N-half of human hexokinase 2 and structural and biochemical characterization of its mitochondrial conformation. Biosci. Rep. 2018, 38, BSR20171666. [Google Scholar] [CrossRef] [Green Version]
- Shityakov, S.; Förster, C. In silico Predictive Model to Determine Vector-Mediated Transport Properties for the Blood-Brain Barrier Choline Transporter. Available online: https://www.dovepress.com/in-silico-predictive-model-to-determine-vector-mediated-transport-prop-peer-reviewed-article-AABC (accessed on 12 August 2019).
- Bischoff, H. The mechanism of alpha-glucosidase inhibition in the management of diabetes. Clin. Investig. Med. 1995, 18, 303–311. [Google Scholar]
- Sarabu, R.; Bizzarro, F.T.; Corbett, W.L.; Dvorozniak, M.T.; Geng, W.; Grippo, J.F.; Haynes, N.-E.; Hutchings, S.; Garofalo, L.; Guertin, K.R.; et al. Discovery of piragliatin—First glucokinase activator studied in Type 2 Diabetic patients. J. Med. Chem. 2012, 55, 7021–7036. [Google Scholar] [CrossRef]
- Bonadonna, R.C.; Heise, T.; Arbet-Engels, C.; Kapitza, C.; Avogaro, A.; Grimsby, J.; Zhi, J.; Grippo, J.F.; Balena, R. Piragliatin (RO4389620), a novel glucokinase activator, lowers plasma glucose both in the postabsorptive state and after a glucose challenge in patients with Type 2 Diabetes Mellitus: A mechanistic study. J. Clin. Endocrinol. Metab. 2010, 95, 5028–5036. [Google Scholar] [CrossRef] [Green Version]
- Zhi, J.; Zhai, S. Effects of piragliatin, a glucokinase activator, on fasting and postprandial plasma glucose in patients with Type 2 Diabetes Mellitus. J. Clin. Pharmacol. 2016, 56, 231–238. [Google Scholar] [CrossRef]
- Liu, S.; Ammirati, M.J.; Song, X.; Knafels, J.D.; Zhang, J.; Greasley, S.E.; Pfefferkorn, J.A.; Qiu, X. Insights into Mechanism of Glucokinase Activation: Observation of multiple distinct protein conformations. J. Biol. Chem. 2012, 287, 13598–13610. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Biochemical Pathway | Classes of Compounds Produced | Examples |
|---|---|---|
| Mevalonate | Terpenes and terpenoids | Beta-caryophyllene, limonene, pinenes, geraniol |
| Non-mevalonate, also called methylerythritol phosphate (MEP) | ||
| Shikimate | Alkaloids, phenylpropanoids, flavonoids, lignans, aromatic polyketides | Cinnamaldehyde, eugenol, coumarin |
| Derivation of fatty acids | Fatty alcohols | Octanol, decanol |
| Cinnamon Essential Oil | ||
|---|---|---|
| Physical Property | Typical Value | Range |
| Optical Rotation | −1 | −20–+20 |
| Specific Gravity | 1.02 | 1.01–1.07 |
| Refractive Index | 1.59 | 1.53–1.61 |
| Flash Point | 87 °C | 62–104 °C |
| Boiling Point | 248 °C | 248–249 °C |
| Color | Dark yellow | Yellow to light brown |
| Aroma | Spicy, sweet | Spicy, sharp, woody |
| C. zeylanicum Bark | Composition | C. zeylanicum Leaf | Composition | C. cassia Bark | Composition | C. cassia Leaf | Composition |
|---|---|---|---|---|---|---|---|
| (E)-Cinnamaldehyde | 44.2–75.7% | Eugenol | 68.6–87.0% | (E)-Cinnamaldehyde | 42.4–89.4% | (E)-Cinnamaldehyde | 54.6–90.1% |
| Eugenol | 1.6–13.3% | Eugenyl acetate | 1.0–8.1% | (Z)-Cinnamaldehyde | 0.6–12.3% | (E)-Cinnamyl acetate | 1.4–12.5% |
| (E)-Cinnamyl acetate | 0.3–10.6% | Linalool | 0.2–5.0% | (E)-Cinnamyl acetate | 0.1–5.4% | (Z)-Cinnamaldehyde | 0.4–10.5% |
| Linalool | 0.2–7.0% | (E)-Cinnamyl acetate | 0.8–4.6% | Benzaldehyde | 0.4–2.3% | Benzaldehyde | 1.1–6.3% |
| beta-Phellandrene | 1.5–8% | Benzyl benzoate | trace–4.1% | alpha-Terpineol | trace–2.0% | Eugenol | trace–5.8% |
| Beta-Caryophyllene | 1.3–6.9% | Beta-Caryophyllene | 1.9–4.1% | Coumarin | trace–1.9% | Cinnamyl alcohol | 0–5.7% |
| p-Cymene | 1.7–4.0% | Safrole | 0–1.3% | Salicylaldehyde | 0.04–1.8% | Salicylaldehyde | 0.05–3.1% |
| 1,8-Cineole | 0.4–2.3% | (E)-Cinnamaldehyde | 0.6–1.1% | Borneol | trace–1.3% | ⍺-copaene | trace–3.0% |
| Benzaldehyde | trace–2.2% | p-Cymene | 0.3–0.8% | Benzyl benzoate | trace–1.0% | Benzyl benzoate | trace–2.9% |
| alpha-Terpineol | 0.4–1.6% | Cinnamyl alcohol | 0–0.6% | Cinnamyl alcohol | 0–0.04% | Delta-Cadinene | trace–2.6% |
| Camphor | trace–1.4% | 1,8-Cineole | trace–0.6% | Coumarin | 0.03–2.5% | ||
| Beta-Phellandrene | 0.2–0.5% | Phenylpropanol | trace–1.6% | ||||
| ⍺-Amorphene | trace–1.1% | ||||||
| Anisaldehyde | 0–1.0% | ||||||
| (E)-Cinnamic acid | trace–0.9% | ||||||
| Methyl eugenol | trace–0.1% |
| Study Product | Study Type | Dosage | Effect | Reference |
|---|---|---|---|---|
| Cinnamon oil | Animal (Rat), KK-Ay | 25, 50, 100 mg/kg b.w. | Significant decrease in fasting blood glucose, plasma C-peptide, serum triglyceride, total cholesterol, and blood urea nitrogen levels, with significant increase in high-density lipoprotein after 35 days. Glucose tolerance was improved and pancreatic islet β-cells showed increased immunoreactivity. | Ping, Zhang, and Ren (2010) [7] |
| Cinnamon oil (encapsulated emulsion) | Animal (Rat), STZ | 200 or 400 mg/kg b.w. | Both doses improved levels of glucose, insulin, SOD, GSH, amylase, lipid profile, and hepatic MDA. Gene expression was modulated to favor antidiabetic outcomes. Positive histological changes seen in liver and pancreas. | Mohammed, Ahmed, Sharaf, El-Nekeety, Abdel-Aziem, Mehaya, Abdel-Wahhab (2020) [35] |
| Cinnamon oil (encapsulated) | Animal (Rat), STZ | 200 or 400 mg/kg b.w. | Treatment with encapsulated cinnamon oil showed improvement in all diabetes-related markers in STZ-treated rats, including liver and kidney function, insulin and glucose levels, lipid profile, and antioxidant enzymes. | Mohammed (2020) [36] |
| Cinnamon oil | Animal (Rat), Alloxan | 5, 10, 20 mg/kg b.w., i.p. | Decreases in fasting blood glucose, total cholesterol, markers of kidney damage and glutathione were observed in treated animals. Histological studies of kidney showed reduced glomerular expansion and tubular dilatations. | Mishra, Bhatti, Singh, Ishar (2010) [37] |
| Cinnamon oil | Animal (Rat), STZ | 100, 200, or 400 mg/kg b.w. | Treatment with cinnamon oil showed significant improvement in histopathology of testicular organs compared to untreated diabetic rats. | Budiastuti, Safitri, Plumeriastuti, Srianto, Effendi (2020) [38] |
| Cinnamon oil | Human | 400 mg/day | Fasting blood glucose levels and insulin levels, along with Quality-of-Life measures, showed improvement after treatment with cinnamon oil, although results were not statistically significant. Pharmacokinetic data indicated low bioavailability. | Stevens (2020) [28] |
| Cinnamon oil | Animal (Mouse), Balb C | 0.2 and 1.0 μL/cage, inhalation | Docking simulations showed interaction of cinnamon oil constituents with leptin receptor in olfactory bulb. In vivo studies confirmed interaction with leptin receptor resulting in decreased appetite and lower weight gain in treated mice. | Kusmardi, Tedjo, Fadilah, Arsianti, Paramita (2018) [39] |
| Cinnamon oil | Animal (Rat), STZ | 5% cinnamon oil in commercial chow | Treatment with cinnamon oil resulted in decreased blood glucose, triglycerides, LDL-cholesterol, and ALT, while levels of HDL-cholesterol were increased compared to diabetic rats. | Zari, Al-Logmani (2009) [40] |
| Cinnamon oil | Animal (Rat), Alloxan | 5, 10, 20 mg/kg b.w., i.p. | Cinnamon oil significantly ameliorated blood glucose levels and thermal hyperalgesia compared to untreated diabetic controls. | Bhatti, Kaur, Singh, Ishar (2009) [41] |
| Cinnamaldehyde | In vitro (HEK293 and 3T3-L1) | -- | Cinnamaldehyde induced expression of peroxisome proliferator-activated (PPAR) genes in 3T3-L1 adipocytes and increased target mRNA expression in HEK293- derived cells. | Li, Futakawa, Yamamoto, Kasahara, Tagami, Liu, and Moriyama (2015) [42] |
| Cinnamaldehyde | Animal (Mouse), DIO-mice | 250 mg/kg/day | Cinnamaldehyde induced significant reduction in cumulative food intake, gastric emptying rates, and ghrelin. Upregulation of genes involved in fatty-acid oxidation was observed in adipose tissue, and mice showed improved glucose tolerance over 5 weeks. | Camacho, Michlig, de Senarclens-Bezencon, Meylan, Meystre, Pezzoli, Markram, le Coutre (2015) [43] |
| Cinnamaldehyde | Animal (Mouse), db/db | 0.02% added to normal chow diet | Treatment with cinnamaldehyde improved aortic tone and function and normalized elevated kidney markers. Treatment also ameliorated glomerular fibrosis and renal dysfunction. Authors suggest a protective effect against vascular dysfunction by inhibiting oxidative stress via Nrf2 signaling pathway activation. | Wang, Yang, Wang, Yang, Wan, Liu, Zhou, Yang (2020) [44] |
| Cinnamaldehyde | Animal (Rat), STZ | 20 mg/kg b.w. | Oral administration led to insulinotropic effects, with increased glucose uptake through GLUT4 receptors and improved function of pyruvate kinase and phosphoenolpyruvate carboxykinase. | Anand, Murali, Tandon, Murthy, Chandra (2010) [45] |
| Cinnamaldehyde | Animal (Rat), FSD/STZ | 20 mg/kg b.w. | Gestating rats treated with cinnamaldehyde showed numerous improvements in health markers compared to diabetic controls, including improved lipid panels and glucose tolerance, more viable fetuses, and improved fetal glucose and insulin levels. | Hosni, Abdel-Moneim, Abdel-Reheim, Mohamed, Helmy (2017) [46] |
| Cinnamaldehyde | Animal (Rat), FSD/STZ | 20 mg/kg b.w. | In rats with gestational diabetes, treatment with cinnamaldehyde prevented development of placental vasculopathy and fetal hypoxia while also alleviating maternal and fetal hyperglycemia. | Hosni, El-Twab, Abdul- Hamid, Prinsen, AbdElgwad, Abdel-Moneim, Beemster (2021) [47] |
| Cinnamaldehyde | Animal (Mouse), STZ | 20 mg/kg/day | Treated mice showed significantly improved insulin sensitivity and glucose metabolism, as well as positive changes in gut microbiota. Authors suggest that modulating host metabolomics may directly or indirectly affect expression levels of genes related to glucose metabolism. | Zhao, Wu, Duan, Liu, Zhu, Zhang, Wang (2021) [48] |
| Cinnamaldehyde | Animal (Rat), STZ | 20 mg/kg/day | Treatment with cinnamaldehyde prevented development of hyperglycemia and insulin resistance following STZ administration. | El-Bassossy, Fahmy, Dadawy (2011) [49] |
| Cinnamaldehyde | Animal (Rat), STZ | 10, 20, 40 mg/kg b.w., p.o. | Rats treated with cinnamaldehyde showed reduced blood glucose levels and amelioration of neurochemical and behavioral deficits seen in diabetic rats. Reductions in IL-2 and TNF-⍺ levels were also noted. | Jawale, Datusalia, Bishnoi, Sharma (2016) [50] |
| Cinnamaldehyde | Rat | 125, 250, 500 mg/kg b.w. | Pharmacokinetic determination of Cmax in rats administered 125, 250, and 500 mg/kg b.w. cinnamaldehyde was 249, 121, and 82 ng/mL serum, respectively. Estimated half-life of cinnamaldehyde was 6.2–6.9 h. | Zhao, Xie, Yang, Cao, Tu, Cao, Wang (2014) [51] |
| Eugenol | Animal (Mouse), STZ | 100 mg/kg b.w. i.p., 2× per week for 2 weeks | Significant reduction in advanced glycation end-products (AGE) and blood glucose levels. | Singh et al. (2014) [52] |
| Eugenol | Animal (Rat), STZ | Treatment with eugenol produced lower blood glucose, decrease in serum glycosylated hemoglobin (HbA1C), lipase, and angiotensin-converting enzyme. Lipid panel levels were also positively affected. | Mnafgui et al. (2013) [53] | |
| Eugenol | Animal (Rat), STZ | 2.5, 5, 10 mg/kg b.w. | Eugenol improved blood glucose and HbA1C levels in diabetic rats and returned glucose metabolism enzyme levels to near normal. Body weight and liver function also improved. | Srinivasan et al. (2013) [54] |
| Eugenol | Animal (Rat), FSD/STZ | 10 mg/kg b.w. | Levels of fasting blood glucose, insulin, triglyceride, cholesterol, and low-density lipoprotein were all improved. Glutathione levels were increased, as were GLUT4 and AMPK levels in skeletal muscle. Homeostasis model assessment of insulin resistance (HOMA-IR) was significantly lower in rats treated with eugenol compared to diabetic controls. | Al-Trad, Alkhateeb, Alsmadi, Al-Zoubi (2019) [55] |
| Eugenol | Animal (Rat), HFD | 20, 40 mg/kg b.w. | Plasma glucose and insulin levels decreased in a dose-dependent manner, and hepatic gluconeogenesis was inhibited via the CAMKK-AMPK-CREB signaling pathway. | Jeong, Kim, Quan, Jo, Kim, Chung (2014) [56] |
| Eugenol | Animal (Rat), STZ | 5, 10 mg/kg b.w. | Diabetic neuropathy parameters (both blood markers and histological changes) were ameliorated in diabetic rats treated with eugenol. Overexpression of TGF-β1 associated with diabetes was also reduced. | Garud, Kulkarni (2017) [57] |
| Eugenol | Animal (Rat), STZ | 10 mg/kg b.w. | Diabetic rats treated with eugenol showed diminished oxidative stress markers and increased antioxidants. In the brain, levels of acetylcholinesterase and calcium were attenuated. Authors postulate that eugenol may help ameliorate diabetic complications due to oxidative stress. | Prasad, Bharath, Muralidhara (2016) [58] |
| Eugenol | Animal (Rat), STZ | 2 mL/day of a 10% nanoemulsion | Oxidative damage was attenuated, and levels of antioxidants were returned to near-normal levels in diabetic rats compared to untreated controls. | Boroujeni, Dehkordi, Sharifi, Taghian, Mazaheri (2021) [59] |
| Eugenol | In vitro (Islets of Langerhans cells from male mouse) | 50, 100, 200 μM | Total antioxidant capacity, superoxide dismutase, and catalase levels increased in cells treated with eugenol following exposure to hydrogen peroxide to induce oxidative stress. Eugenol can bolster antioxidant systems in islet cells that are particularly vulnerable to oxidative stress in diabetics. | Oroojan, Chenani, An’aam (2020) [60] |
| Eugenol | Animal (Rat), Alloxan | 5, 10, 15 mg/kg b.w. | Diabetic rats treated with eugenol showed lower fasting blood glucose, and improved morphology of liver and islet of Langerhans cells. | Hamdin, Utami, Muliasari, Prasedya, Sudarma (2019) [61] |
| β-caryophyllene | In silico | -- | β-caryophyllene showed affinity for interaction with insulin downstream signaling molecules such as IRS-1, cSrc, and Akt. | Mani, Balraj, Venktsan, Soundrapandiyan, Kasthuri, Danavel, Babu (2021) [62] |
| β-caryophyllene | Animal (Rat), STZ | 10 mg/kg b.w. | Diabetic neuropathy was attenuated in rats treated with β-caryophyllene. Depression behavior and cytokine markers of diabetes were also reduced. | Aguilar-Ávila, Flores-Soto, Tapia-Vázquez, Pastor-Zarandona, López-Roa, Viveros-Paredes (2019) [63] |
| β-caryophyllene | Animal (Rat), STZ | 200 mg/kg b.w. | Hyperglycemia was attenuated by treatment with β-caryophyllene, and oxidative stress was avoided through increased activity of antioxidant enzymes. | Basha, Sankaranarayanan (2016) [64] |
| β-caryophyllene | Animal (Rat), STZ | 200 mg/kg b.w. | Plasma insulin levels were rescued to near-normal levels in diabetic rats treated with β-caryophyllene. | Basha, Sankaranarayanan (2015) [65] |
| β-caryophyllene | Animal (Rat), STZ | 100, 200, 400 mg/kg b.w. | Administration of β-caryophyllene ameliorated STZ-induced changes in blood glucose, insulin levels, and glucose metabolism enzymes. The antidiabetic and insulinotropic effects were most pronounced at the 200 mg/kg dose. | Basha, Sankaranarayanan (2014) [66] |
| β-caryophyllene | Animal (Rat), HFD | 30 mg/kg b.w. | Treatment with β-caryophyllene improved glycemic and lipidemic markers and reduced vascular oxidative stress and inflammation. | Youssef, El-Fayoumi, Mahmoud (2019) [67] |
| β-caryophyllene | In vitro (mesangial cells) | -- | β-caryophyllene modulated NF-κB and Nrf pathways and exhibited anti-inflammatory and nephroprotective activity in mesangial cells under high-glucose conditions. | Li, Wang, Chen, Yang (2020) [68] |
| β-caryophyllene | In vitro (C2C12 myotubes) | -- | β-caryophyllene significantly increased skeletal muscle uptake of glucose and glycolytic production of ATP through cannabinoid receptor-2-mediated pathways. | Geddo et al. (2021) [69] |
| Essential Oil Ligand | PTP1B | α-Glucosidase | Glucokinase | α-Amylase | HK-II |
| β-caryophyllene | −5.6 | −6.6 | −6.3 | −6.9 | −6.6 |
| Cinnamaldehyde | −6.5 | −6.1 | −5.8 | −5.4 | −5.5 |
| Eugenol | −6.0 | −6.7 | −6.0 | −5.6 | −5.6 |
| Positive Controls | PTP1B | α-Glucosidase | Glucokinase | α-Amylase | HK-II |
| Acarbose | -- | −8.3 | -- | −7.3 | -- |
| Piragliatin | -- | -- | −9.5 | -- | -- |
| TPICOOH | −9.7 | -- | -- | -- | -- |
| Benserazide | -- | -- | -- | -- | −7.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stevens, N.; Allred, K. Antidiabetic Potential of Volatile Cinnamon Oil: A Review and Exploration of Mechanisms Using In Silico Molecular Docking Simulations. Molecules 2022, 27, 853. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27030853
Stevens N, Allred K. Antidiabetic Potential of Volatile Cinnamon Oil: A Review and Exploration of Mechanisms Using In Silico Molecular Docking Simulations. Molecules. 2022; 27(3):853. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27030853
Chicago/Turabian StyleStevens, Nicole, and Kathryn Allred. 2022. "Antidiabetic Potential of Volatile Cinnamon Oil: A Review and Exploration of Mechanisms Using In Silico Molecular Docking Simulations" Molecules 27, no. 3: 853. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27030853