
Anti-Obesity Effects of a Mixture of Atractylodes macrocephala and Amomum villosum Extracts on 3T3-L1 Adipocytes and High-Fat Diet-Induced Obesity in Mice



Abstract
:1. Introduction
2. Results
2.1. Cytotoxic Effects of the Mixture of Atractylodes macrocephala and Amomum villosum Extracts (3:1) in 3T3-L1 Preadipocytes
2.2. Effects of Mixtures of Atractylodes macrocephala and Amomum villosum Extracts (3:1) on Adipocyte Differentiation in 3T3-L1 Preadipocytes
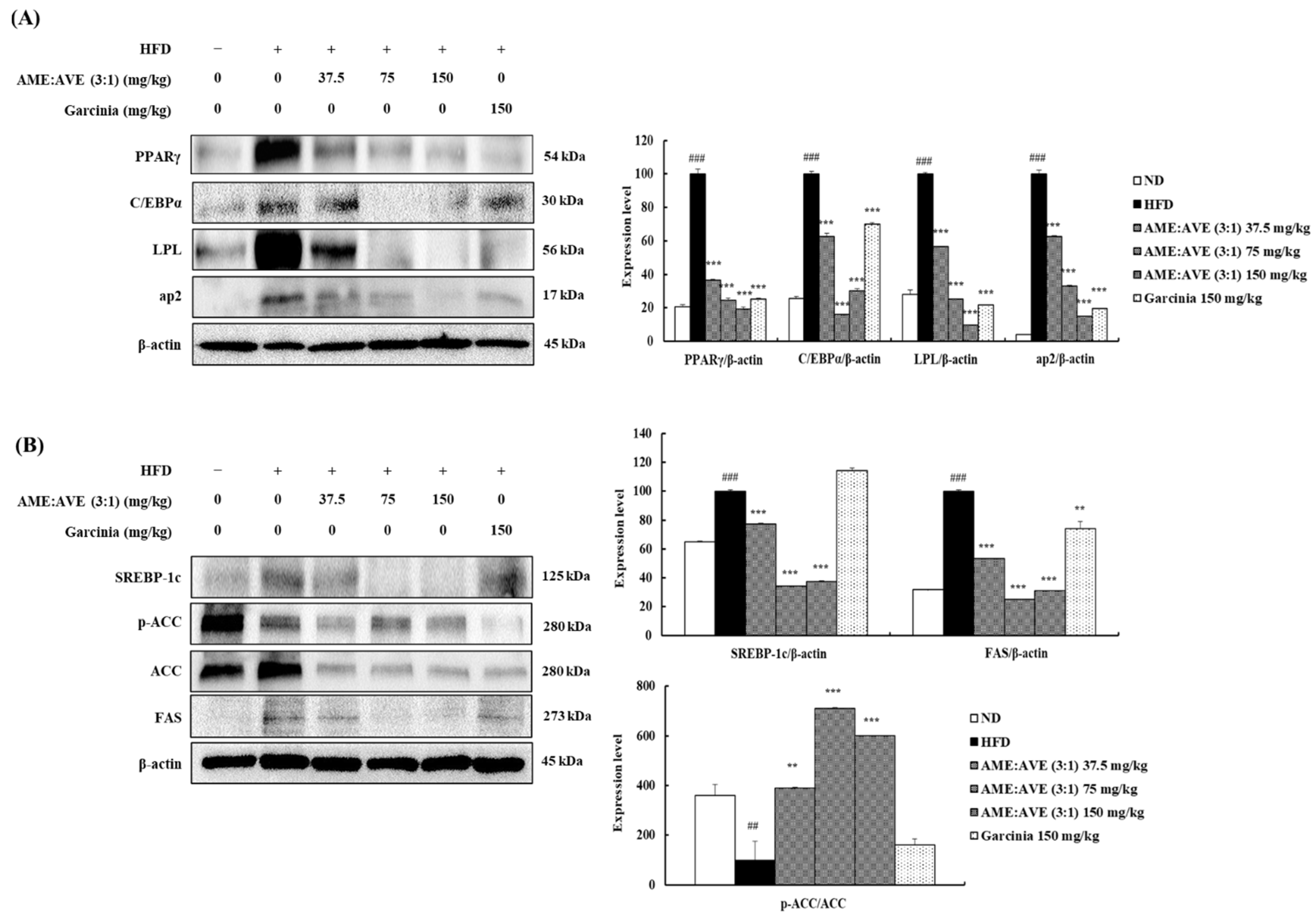
2.3. Effects of Mixtures of Atractylodes macrocephala and Amomum villosum Extracts (3:1) on Proteins in Adipogenesis and Lipogenesis Downstream in 3T3-L1 Adipocytes
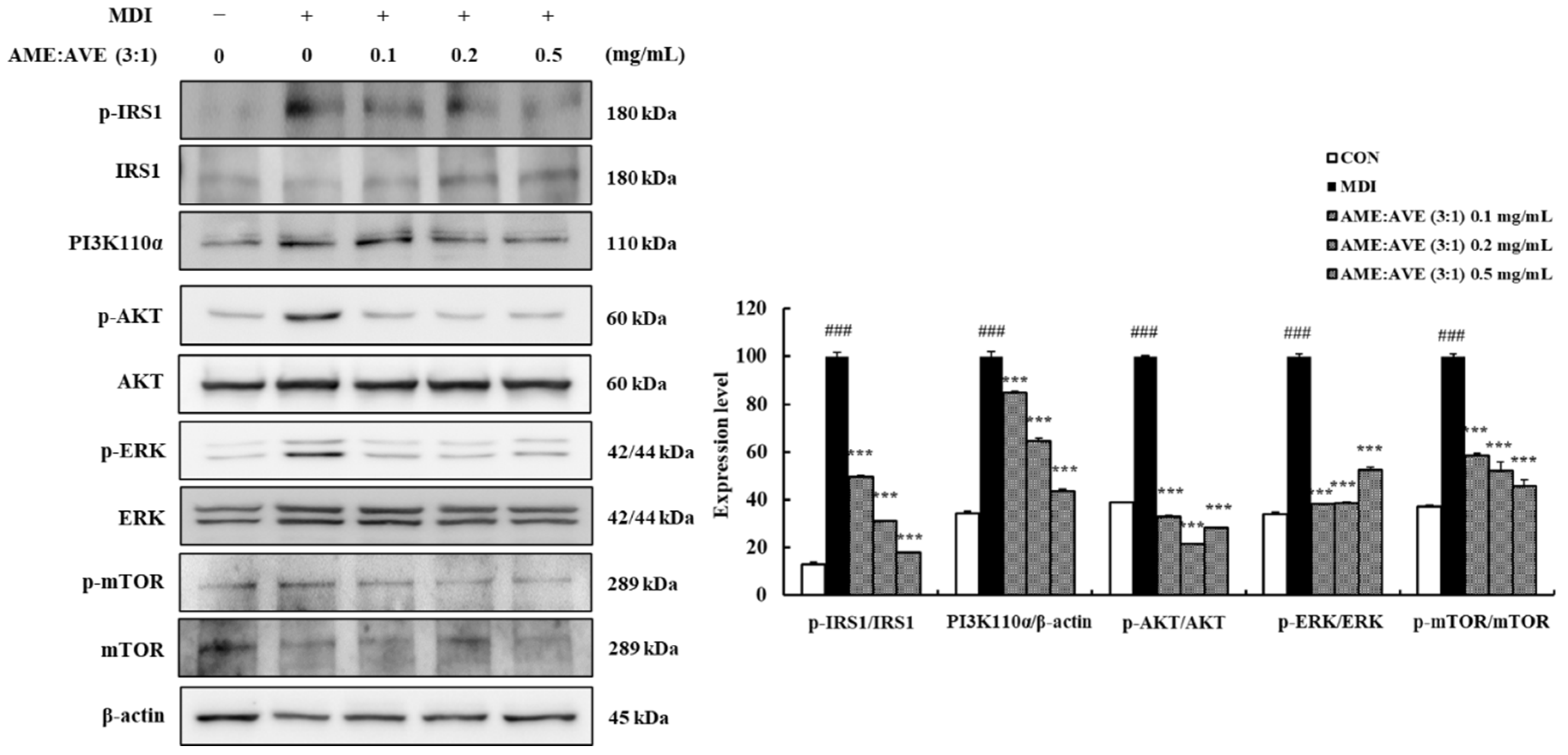
2.4. Effects of Atractylodes macrocephala and Amomum villosum Extracts (3:1) on Upstream Proteins in Adipogenesis and Lipogenesis in 3T3-L1 Adipocytes
2.5. Effects of Atractylodes macrocephala and Amomum villosum Extracts (3:1) on Body and Organ Weights of High-Fat Diet-Induced Obesity in Mice
2.6. Effects of Mixtures of Atractylodes macrocephala and Amomum villosum Extracts (3:1) on Liver and Epididymal Fat Morphology in High-Fat Diet-Induced Obesity in Mice
2.7. Effects of Atractylodes macrocephala and Amomum villosum Extracts (3:1) on Plasma Biochemical Parameters in High-Fat Diet-Induced Obesity in Mice
2.8. Effects of Mixtures of Atractylodes macrocephala and Amomum villosum Extracts (3:1) on Adipogenesis- and Lipogenesis-Related Protein Levels in High-Fat Diet-Induced Obesity in Mice
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of AME:AVE (3:1) Extracts
4.3. Cell Culture
4.4. MTT Assay for Cell Viability
4.5. Oil-Red O Staining
4.6. Animals and Diets
4.7. Biochemical Analysis
4.8. Histological Analysis
4.9. Protein Extraction and Western Blot Analysis
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Giri, S.; Rattan, R.; Haq, E.; Khan, M.; Yasmin, R.; Won, J.S.; Key, L.; Singh, A.K.; Singh, I. AICAR inhibit adipocyte differentiation in 3T3L1 and restores metabolic alterations in diet-induced obesity mice model. Nutr. Metab. 2006, 3, 31–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.K.; Kong, C.S. Anti-adipogenic effect of dioxinodehydoeckol via AMPK activation in 3T3-L1 adipocytes. Chem. Biol. 2010, 186, 24–29. [Google Scholar]
- Spiegelman, M.B.; Flier, J.S. Adipogenesis and obesity: Rounding out the big picture. Cell 1996, 87, 377–389. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, K. Management of obesity as a chronic disease: Non- pharmacologic, pharmacologic, and surgical options. Obes. Res. 2002, 10, 116S–123S. [Google Scholar] [CrossRef] [Green Version]
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [CrossRef]
- Song, Y.; Park, H.J.; Kang, S.N.; Jang, S.-H.; Lee, S.-J.; Ko, Y.-G.; Kim, G.-S.; Cho, J.-H. Blueberry Peel Extracts Inhibit Adipogenesis in 3T3-L1 Cells and Reduce High-Fat Diet-Induced Obesity. PLoS ONE 2013, 8, e69925. [Google Scholar] [CrossRef] [Green Version]
- Jeon, T.; Hwang, S.G.; Hirai, S.; Matsui, T.; Yano, H.; Kawada, T.; Lim, B.O.; Park, D.K. Red yeast rice extracts suppress adipogenesis by down-regulating adipogenic transcription factors and gene expression in 3T3-L1 cells. Life Sci. 2004, 75, 3195–3203. [Google Scholar] [CrossRef]
- Hong, H.; Park, J.; Lumbera, W.L.; Hwang, S.G. Monascus ruber-fermented buckwheat (Red Yeast Buckwheat) suppresses adipo-genesis in 3T3-L1 cells. J. Med. Food 2017, 20, 352–359. [Google Scholar]
- Lee, D.-R.; Lee, Y.-S.; Choi, B.-K.; Lee, H.J.; Park, S.-B.; Kim, T.-M.; Oh, H.J.; Yang, S.H.; Suh, J.-W. Roots extracts of Adenophora triphylla var. japonica improve obesity in 3T3-L1 adipocytes and high-fat diet-induced obese mice. Asian Pac. J. Trop. Med. 2015, 8, 898–906. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.L.; Lee, H.J.; Choi, B.K.; Park, S.B.; Woo, S.M.; Lee, D.R. Roots Extract of Adenophora triphylla var. japonica Inhibits Ad-ipogenesis in 3T3-L1 Cells through the Downregulation of IRS1. J. Physiol. Pathol. Korean Med. 2020, 34, 136–141. [Google Scholar] [CrossRef]
- Derosa, G.; Maffifioli, P. Anti-obesity drugs: A review about their effects and their safety. Expert Opin. Drug Saf. 2012, 11, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.A.; Hocht, C.L.; Puyo, A.; Taira, C.A. Recent advances in obesity pharmacotheraphy. Curr. Clin. Pharmacol. 2009, 4, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.W. Possible anti-obesity therapeutics from nature–a review. Phytochemistry 2010, 71, 1625–1641. [Google Scholar] [CrossRef] [PubMed]
- China Pharmacopoeia Committee. Pharmacopoeia of the People’s Republic of China (Part 1); China Chemical Industry Press: Beijing, China, 2005; p. 58.
- Shan, Z.J. Check and Punctuate on Shennong’s Classic of Herbology; Wannan Medial College Press: Hefei, China, 1981; p. 48. [Google Scholar]
- Chen, Q.; Li, P.; Yang, H.; Zhu, J. Sensitive capillary GC-MS-SIM determination of Atractylenolide I and Atractylenolide III in At-ractylodes macrocephala. Anal. Lett. 2009, 42, 2547–2555. [Google Scholar] [CrossRef]
- Sun, X.; Jian, G. Simultaneous determination of atractylenolide II and III in Rhizoma Atractylodes macrocephalae and Chinese medicinal preparation by reverse-phase high-performance liquid chromatography. Lect. Notes Electr. Eng. 2014, 269, 1283–1289. [Google Scholar]
- Li, X.; Lin, J.; Han, W.; Mai, W.; Wang, L.; Li, Q.; Lin, M.; Bai, M.; Zhang, L.; Chen, D. Antioxidant Ability and Mechanism of Rhizoma Atractylodes macrocephala. Molecules 2012, 17, 13457–13472. [Google Scholar] [CrossRef] [Green Version]
- Li, C.-Q.; He, L.-C.; Dong, H.-Y.; Jin, J.-Q. Screening for the anti-inflammatory activity of fractions and compounds from Atractylodes macrocephala Koidz. J. Ethnopharmacol. 2007, 114, 212–217. [Google Scholar] [CrossRef]
- Huang, Q.S.; Li, H.Z.; Zhang, Y.L.; Qiu, S.L. The research on the effect of abstraction of Atractylodes on the ability of resisting mutation of human peripheral blood lymphocytes for Cr 6+. Prog. Mod. Biomed. 2006, 6, 20–21. [Google Scholar]
- Wen, Z.; Liu, B.; Zheng, Z.; You, X.; Pu, Y.; Li, Q. Preparation of liposomes entrapping essential oil from Atractylodes macrocephala Koidz by modified RESS technique. Chem. Eng. Res. Des. 2010, 88, 1102–1107. [Google Scholar] [CrossRef]
- Zhou, S. Cultivation of Amomum villosum in tropical forests. For. Ecol. Manag. 1993, 60, 157–162. [Google Scholar] [CrossRef]
- Huang, Q.; Duan, Z.; Yang, J.; Ma, X.; Zhan, R.; Xu, H.; Chen, W. SNP Typing for Germplasm Identification of Amomum villosum Lour. Based on DNA Barcoding Markers. PLoS ONE 2014, 9, e114940. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.R.; Kwon, Y.K.; Choi, B.K.; Baek, D.G. Inhibitiory Effects of Mixture of Atractylodes macrocephala and Amomum villosum Extracts on Adipocyte Differentiation in OP9 Cells. J. Physiol. Pathol. Korean Med. 2020, 34, 24–29. [Google Scholar] [CrossRef]
- Kim, H.R.; Choi, B.K.; Jung, H.J. Anti-Obesity Effects of Mixture of Atractylodes macrocephala and Amomum villosum Extracts. J. Physiol. Pathol. Korean Med. 2019, 33, 282–287. [Google Scholar] [CrossRef]
- Kim, H.R.; Kwon, Y.K.; Choi, B.K.; Jung, H.J.; Baek, D.G. Effect of Mixture of Atractylodes macrocephala and Amomum villosum Extracts on Body Weight and Lipid Metabolism in High Fat Diet-Induced Obesity Model. J. Physiol. Pathol. Korean Med. 2020, 34, 75–80. [Google Scholar] [CrossRef]
- Song, M.Y.; Lim, S.K.; Wang, J.H.; Kim, H. The Root of Atractylodes macrocephala Koidzumi Prevents Obesity and Glucose Intol-erance and Increases Energy Metabolism in Mice. Int. J. Mol. Sci. 2018, 19, 278. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Zhang, T.; Gu, W.; Yang, X.; Lu, J.; Zhao, R.; Yu, J. Volatile Oil of Amomum villosum Inhibits Nonalcoholic Fatty Liver Disease via the Gut-Liver Axis. BioMed Res. Int. 2018, 2018, 3589874. [Google Scholar] [CrossRef] [Green Version]
- Ducharme, N.A.; Bickel, P.E. Minireview: Lipid Droplets in Lipogenesis and Lipolysis. Endocrinology 2008, 149, 942–949. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, M.; Leibel, R.L.; Hirsch, J. Obesity. N. Engl. J. Med. 1997, 337, 396–407. [Google Scholar] [CrossRef]
- Hsieh, Y.H.; Wang, S.Y. Lucidone from Linera erythrocarpa Makino fruits suppresses adipogenesis in 3T3-L1 cells and attenuates obesity and consequent metabolic disorders in high-fat diet C57BL/6 mice. Phytomedicine 2013, 20, 394–400. [Google Scholar] [CrossRef]
- Browning, J.D.; Horton, J.D. Molecular mediators of hepatic steatosis and liver injury. J. Clin. Investig. 2004, 114, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Marchesini, G.; Moscatiello, S.; Di Domizio, S.; Forlani, G. Obesity-Associated Liver Disease. J. Clin. Endocrinol. Metab. 2008, 93, s74–s80. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, K.; Shibata, R.; Murohara, T.; Ouchi, N. Role of anti-inflammatory adipokines in obesity-related diseases. Trends Endocrinol. Metab. 2014, 25, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Miki, H.; Yamauchi, T.; Suzuki, R.Y.O.; Komeda, K.; Tsuchida, A.; Kubota, N.; Terauchi, Y.; Kamon, J.; Kaburagi, Y.; Matsui, J.; et al. Essential role of insulin receptor substrate 1 (IRS-1) and IRS-2 in adipocyte differentiation. Mol. Cell. Biol. 2001, 21, 2521–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef]
- Petersen, K.F.; Shulman, G. Etiology of Insulin Resistance. Am. J. Med. 2006, 119, S10–S16. [Google Scholar] [CrossRef] [Green Version]
- Boucher, J.; Kleinridders, A.; Kahn, C.R. Insulin Receptor Signaling in Normal and Insulin-Resistant States. Cold Spring Harb. Perspect. Biol. 2014, 6, a009191. [Google Scholar] [CrossRef] [Green Version]
- Zoncu, R.; Efeyan, A.; Sabatini, D.M. mTOR: From growth signal integration to cancer, diabetes and ageing. Nat. Rev. Mol. Cell Biol. 2011, 12, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.B.; Spiegelman, B.M. ADD1/SREBP1 promotes adipocyte differentiation and gene expression linked to fatty acid metabolism. Genes Dev. 1996, 10, 1096–1107. [Google Scholar] [CrossRef] [Green Version]
- Farmer, S.R. Regulation of PPAR γ activity during adipogenesis. Int. J. Obes. 2005, 29, S13–S16. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar] [CrossRef]
- Park, J.; Rho, H.K.; Kim, K.H.; Choe, S.S.; Lee, Y.S.; Kim, J.B. Overexpression of Glucose-6-Phosphate Dehydrogenase Is Associated with Lipid Dysregulation and Insulin Resistance in Obesity. Mol. Cell. Biol. 2005, 25, 5146–5157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, J.; Bihain, B.E.; Bengtsson-Olivecrona, G.; Deckelbaum, R.J.; Carpentier, Y.A.; Olivecrona, T. Fatty acid control of lipo-protein lipase: A link between energy metabolism and lipid transport. Proc. Natl. Acad. Sci. USA 1990, 87, 909–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.-H. Regulation of Mammalian Acetyl-Coenzyme A Carboxylase. Annu. Rev. Nutr. 1997, 17, 77–99. [Google Scholar] [CrossRef] [PubMed]
- Nepali, S.; Kim, D.-K.; Lee, H.-Y.; Ki, H.-H.; Kim, B.-R.; Hwang, S.-W.; Park, M.; Kim, D.-K.; Lee, Y.-M. Euphorbia supina extract results in inhibition of high-fat-diet-induced obesity in mice. Int. J. Mol. Med. 2018, 41, 2952–2960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Choi, Y.H.; Lee, J.H.; Lee, S.Y.; Kang, I.J. Anti-Obesity Effect of Erigeron annuus (L.) Pers. Extract Containing Phenolic Acids. Foods 2021, 10, 1266. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Lee, J.; Lee, E.-H.; In, G.; Kim, J.; Lee, M.-H.; Lee, O.-H.; Kang, I.-J. A Combination of Korean Red Ginseng Extract and Glycyrrhiza glabra L. Extract Enhances Their Individual Anti-Obesity Properties in 3T3-L1 Adipocytes and C57BL/6J Obese Mice. J. Med. Food 2020, 23, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Campitelli, M.; Desiderio, A.; Cacace, G.; Nigro, C.; Prevenzano, I.; Leone, A.; de Simone, S.; Campiglia, P.; Formisano, P.; Raciti, G.A.; et al. Citrus aurantium L. Dry Extracts Ameliorate Adipocyte Differentiation of 3T3-L1 Cells Exposed to TNFα by Down-Regulating miR-155 Expression. Nutrients 2020, 12, 1587. [Google Scholar]
- Ilmiawati, C.; Fitri, F.; Rofinda, Z.D.; Reza, M. Green coffee extract modifies body weight, serum lipids and TNF-α in high-fat di-et-induced obese rats. BMC Res. Notes 2020, 13, 208. [Google Scholar] [CrossRef] [Green Version]
- Ji-Ping, L.; Ren-Chao, T.; Xiao-Meng, S.; Hao-Yue, Z.; Shuai, S.; Ai-Zhen, X.; Zheng-Tao, W.; Li, Y. Comparison of main chemical composition of Plantago asiatica L. and P. depressa Willd. seed extracts and their anti-obesity effects in high-fat diet-induced obese mice. Phytomedicine 2020, 81, 153362. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indexes | ND | HFD | HFD | |||
|---|---|---|---|---|---|---|
| AME:AVE (3:1) 37.5 | AME:AVE (3:1) 75 | AME:AVE (3:1) 150 | G 150 | |||
| Glucose (mg/dL) | 251.0 ± 15.1 | 374.5 ± 13.9 ### | 305.9 ± 9.8 ** | 325.3 ± 2.8 ** | 268.6 ± 6.0 *** | 301.8 ± 8.7 ** |
| T-CHO (mg/dL) | 97.5 ± 3.7 | 205.5 ± 19.5 ### | 186.0 ± 5.1 | 160.2 ± 3.1 * | 159.5 ± 4.0 | 165.5 ± 4.9 |
| TG (mg/dL) | 73.0 ± 2.6 | 98.3 ± 5.3 ## | 106.3 ± 3.2 | 92.5 ± 3.1 | 85.6 ± 4.4 | 83.5 ± 2.0 * |
| LDL-c (mg/dL) | 9.4 ± 0.2 | 37.7 ± 7.0 ## | 32.7 ± 0.8 | 29.3 ± 0.9 | 31.5 ± 1.3 | 31.9 ± 1.3 |
| HDL-c/LDL-c | 11.3 ± 0.5 | 4.7 ± 0.4 ## | 4.9 ± 0.1 | 5.3 ± 0.2 | 4.9 ± 0.3 | 4.8 ± 0.3 |
| ALT (U/L) | 29.3 ± 3.9 | 50.6 ± 6.9 ## | 40.6 ± 4.5 | 32.6 ± 1.4 * | 32.7 ± 2.6 * | 29.8 ± 0.9 * |
| AST (U/L) | 129.4 ± 8.6 | 185.9 ± 20.2 # | 144.6 ± 9.3 | 140.2 ± 3.6 * | 125.1 ± 8.5 * | 134.7± 2.9 * |
| Leptin (ng/mL) | 1.5 ± 0.1 | 48.8 ± 9.1 ### | 35.4 ± 3.1 | 26.0 ± 1.4 * | 26.7 ± 6.7 | 33.6 ± 3.7 |
| Adiponectin (ng/mL) | 14.2 ± 1.0 | 13.9 ± 0.5 | 14.1 ± 0.6 | 14.4 ± 1.1 | 14.7 ± 0.3 | 12.0 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-L.; Lee, S.-K.; Min, D.-E.; Choi, B.-K.; Lee, D.-R. Anti-Obesity Effects of a Mixture of Atractylodes macrocephala and Amomum villosum Extracts on 3T3-L1 Adipocytes and High-Fat Diet-Induced Obesity in Mice. Molecules 2022, 27, 906. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27030906
Kim H-L, Lee S-K, Min D-E, Choi B-K, Lee D-R. Anti-Obesity Effects of a Mixture of Atractylodes macrocephala and Amomum villosum Extracts on 3T3-L1 Adipocytes and High-Fat Diet-Induced Obesity in Mice. Molecules. 2022; 27(3):906. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27030906
Chicago/Turabian StyleKim, Hae-Lim, Sung-Kwon Lee, Da-Eun Min, Bong-Keun Choi, and Dong-Ryung Lee. 2022. "Anti-Obesity Effects of a Mixture of Atractylodes macrocephala and Amomum villosum Extracts on 3T3-L1 Adipocytes and High-Fat Diet-Induced Obesity in Mice" Molecules 27, no. 3: 906. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27030906