
Essential Oils Derived from Cistus Species Activate Mitochondria by Inducing SIRT1 Expression in Human Keratinocytes, Leading to Senescence Inhibition

 and
and 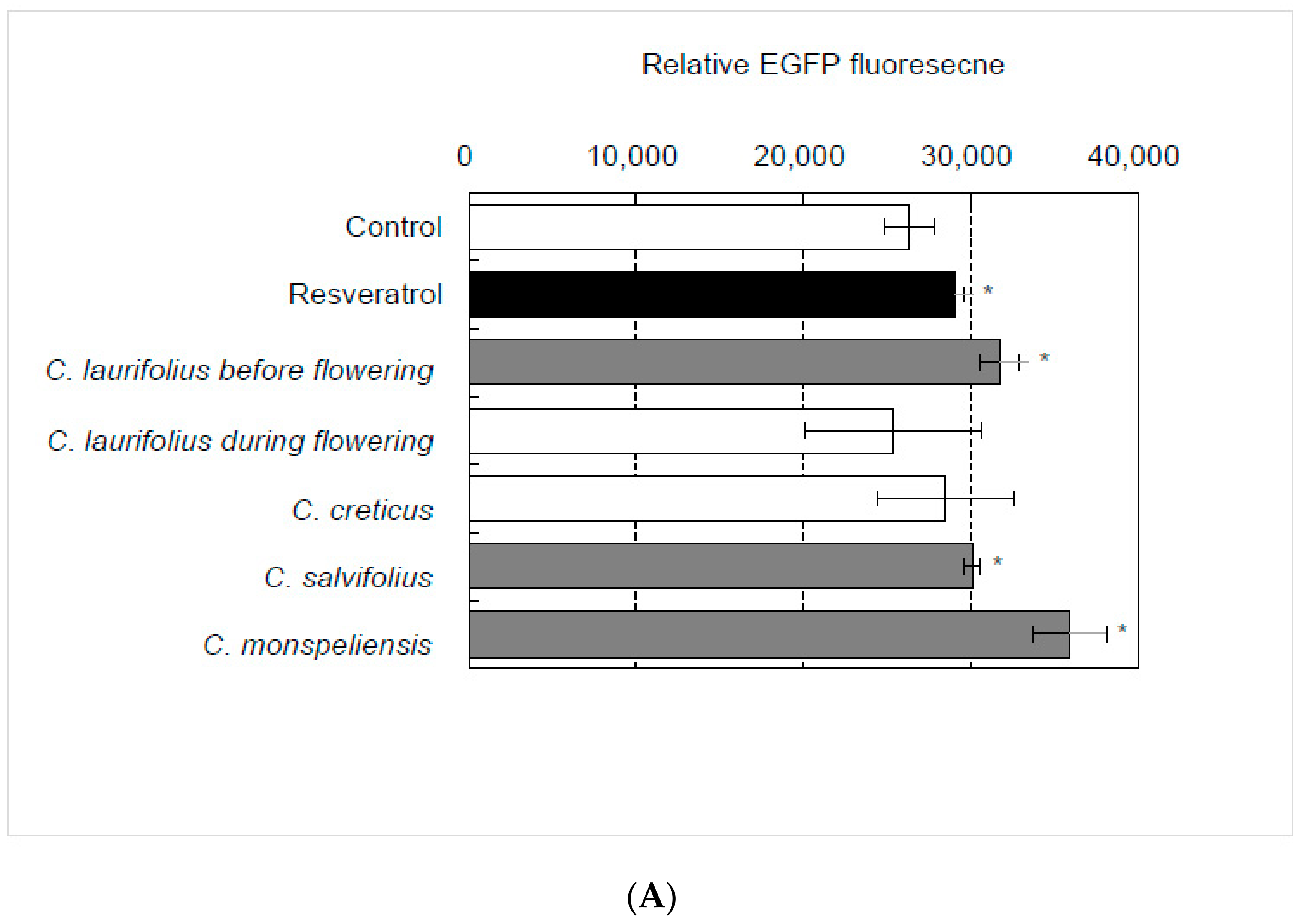{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of Essential Oil That Activates SIRT1 Transcription
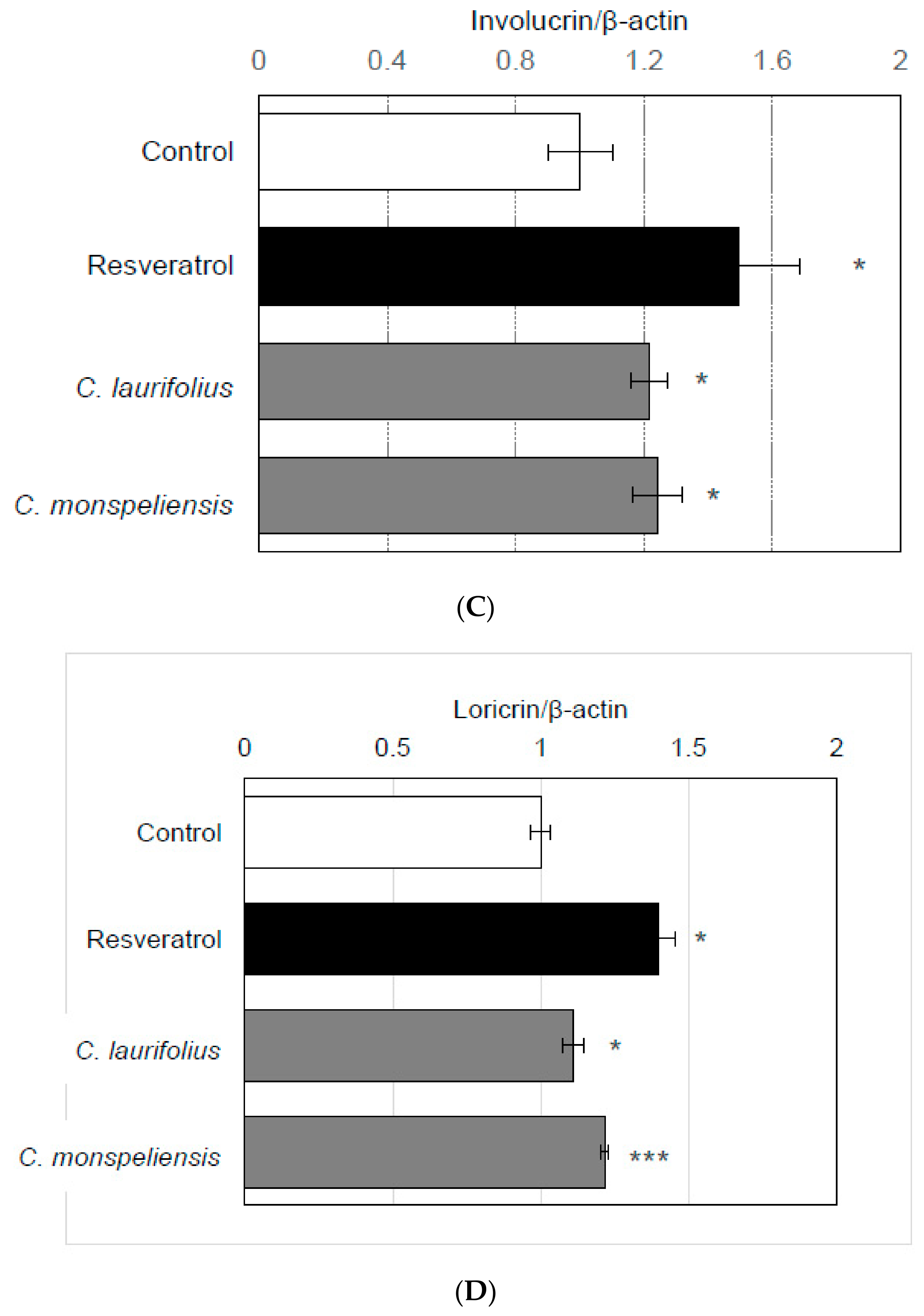
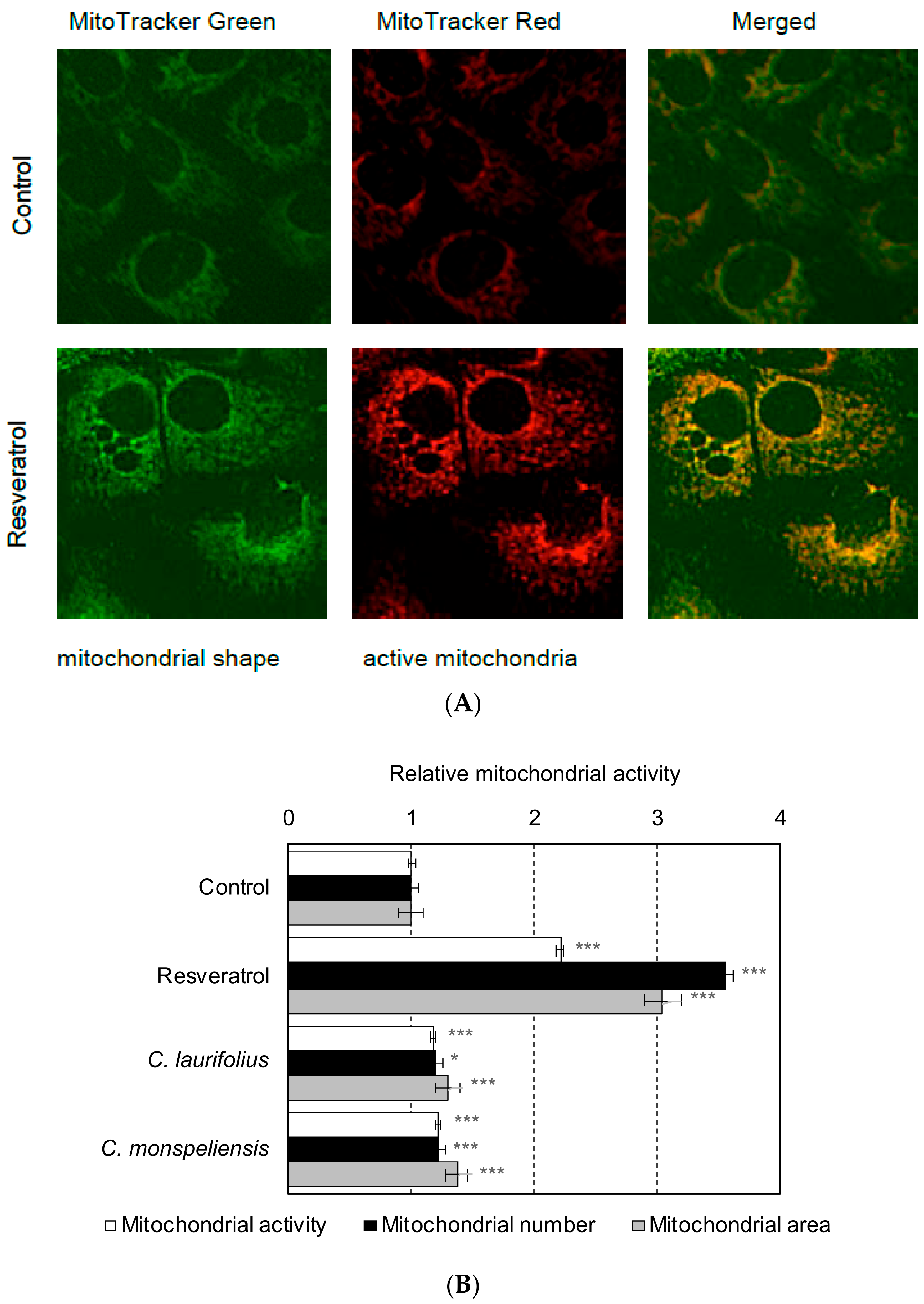
2.2. Effects of Cistus Essential Oils on the Mitochondrial Biogenesis and Differentiation
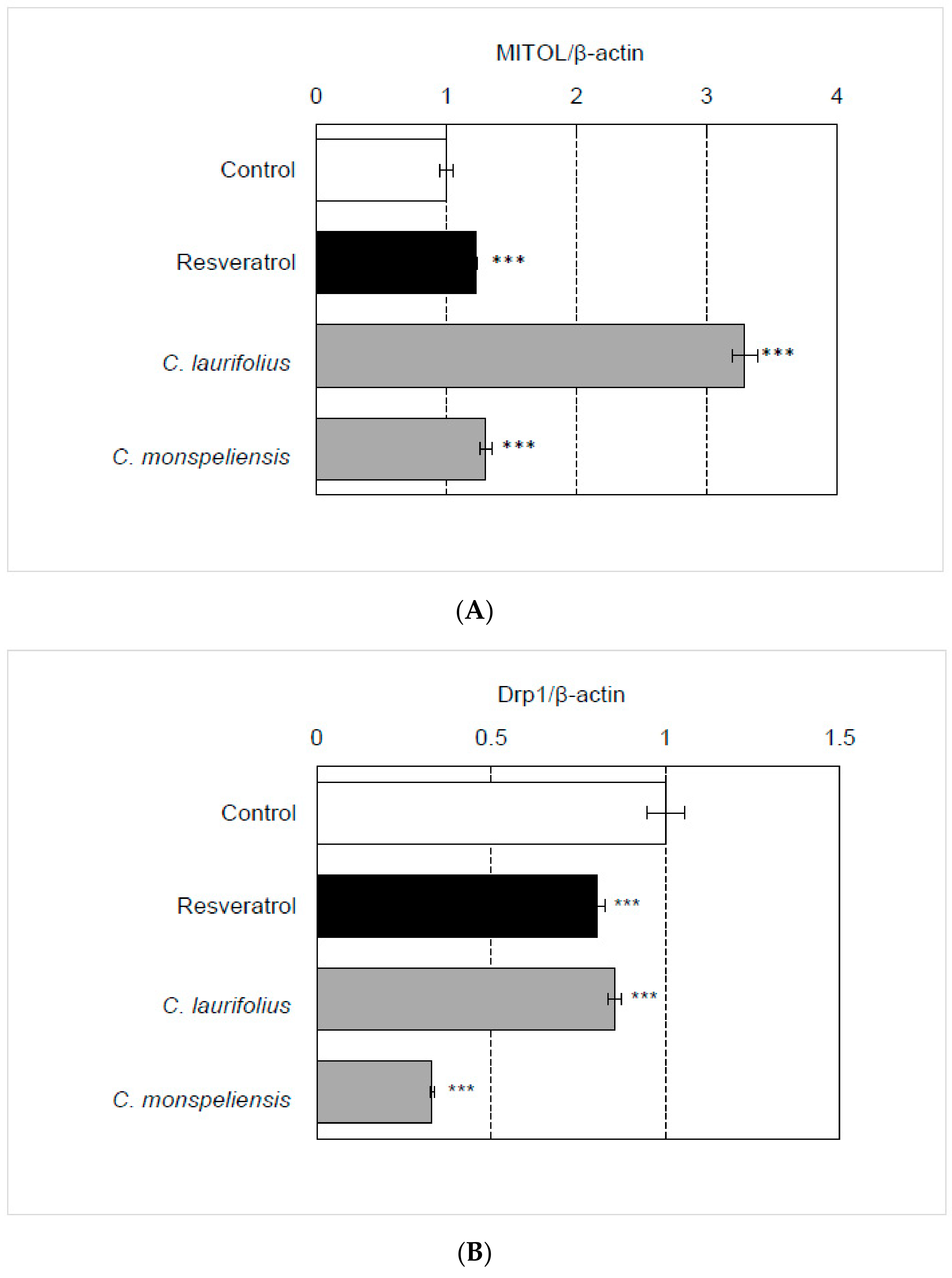
2.3. Effects of Cistus Essential Oils on UVB-Induced Mitochondrial Damage and Senescence
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Essential Oil Extraction
4.2. Cell Line and Treatment
4.3. SIRT1 Promoter Reporter Assay
4.4. Quantitative Reverse Transcription Polymerase Chain Reaction
4.5. Western Blot
4.6. DNA Isolation
4.7. Determination of Mitochondrial DNA (mtDNA) Copy Number
4.8. Mitochondrial Imaging
4.9. Ultraviolet B Irradiation
4.10. Fluorescent Senescence-Associated β-Galactosidase Assay
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Papaefthimiou, D.; Papanikolaou, A.; Falara, V.; Givanoudi, S.; Kostas, S.; Kanellis, A.K. Genus Cistus: A model for exploring labdane-type diterpenes’ biosynthesis and a natural source of high value products with biological, aromatic, and pharmacological properties. Front. Chem. 2014, 2, 198. [Google Scholar] [CrossRef] [Green Version]
- Bellakhdar, J. La Pharmacopée Marocaine Traditionnelle; Ibis Press: Paris, France, 1997. [Google Scholar]
- Zalegh, I.; Akssira, M.; Bourhia, M.; Mellouki, F.; Rhallabi, N.; Salamatullah, A.M.; Alkaltham, M.S.; Alyahya, H.K.; Mhand, R.A. A Review on Cistus sp.: Phytochemical and Antimicrobial Activities. Plants 2021, 10, 1214. [Google Scholar] [CrossRef]
- Preedy, V.R. Essential Oils in Food Preservation, Flavor and Safety; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Baur, J.A.; Ungvari, Z.; Minor, R.K.; Le Couteur, D.G.; de Cabo, R. Are sirtuins viable targets for improving healthspan and lifespan? Nat. Rev. Drug Discov. 2012, 11, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, B.L. Sirt1 and the Mitochondria. Mol. Cells 2016, 39, 87. [Google Scholar] [PubMed] [Green Version]
- Chong, Z.; Matsuo, H.; Onoue, S.; Yamamoto, H.; Ito, H.; Katakura, Y. Identification of polyphenols that repair the ultraviolet-B-induced DNA damage via SIRT1-dependent XPC/XPA activation. J. Funct. Foods 2019, 54, 119–127. [Google Scholar] [CrossRef]
- Zhao, C.; Sakaguchi, T.; Fujita, K.; Ito, H.; Nishida, N.; Nagatomo, A.; Tanaka-Azuma, Y.; Katakura, Y. Pomegranate-derived polyphenols reduce reactive oxygen species production via SIRT3-mediated SOD2 Activation. Oxid. Med. Cell Longev. 2016, 2016, 2927131. [Google Scholar] [CrossRef] [Green Version]
- Pendergrass, W.; Wolf, N.; Poot, M. Efficacy of MitoTracker Green and CMXrosamine to measure changes in mitochondrial membrane potentials in living cells and tissues. Cytometry A 2004, 61, 162–169. [Google Scholar] [CrossRef]
- Nagashima, S.; Tokuyama, T.; Yonashiro, R.; Inatome, R.; Yanagi, S. Roles of mitochondrial ubiquitin ligase MITOL/MARCH5 in mitochondrial dynamics and diseases. J. Biochem. 2014, 155, 273–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Küpeli, E.; Orhan, D.D.; Yesilada, E. Effect of Cistus laurifolius L. leaf extracts and flavonoids on acetaminophen-induced hepatotoxicity in mice. J. Ethnopharmacol. 2006, 103, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Ustun, O.; Ozçelik, B.; Akyön, Y.; Abbasoglu, U.; Yesilada, E. Flavonoids with anti-Helicobacter pylori activity from Cistus laurifolius leaves. J. Ethnopharmacol. 2006, 108, 457–461. [Google Scholar] [CrossRef]
- Orhan, N.; Aslan, M.; Süküroğlu, M.; Deliorman Orhan, D. In vivo and in vitro antidiabetic effect of Cistus laurifolius L. and detection of major phenolic compounds by UPLC-TOF-MS analysis. J. Ethnopharmacol. 2013, 146, 859–865. [Google Scholar] [CrossRef]
- Sadhu, S.K.; Okuyama, E.; Fujimoto, H.; Ishibashi, M.; Yesilada, E. Prostaglandin inhibitory and antioxidant components of Cistus laurifolius, a Turkish medicinal plant. J. Ethnopharmacol. 2006, 108, 371–378. [Google Scholar] [CrossRef]
- Hong, S.-P.; Seo, H.-S.; Shin, K.-O.; Park, K.; Park, B.C.; Kim, M.H.; Park, M.; Kim, C.D.; Seo, S.J. Adiponectin enhances human keratinocyte lipid synthesis via SIRT1 and nuclear hormone receptor signaling. J. Investig. Dermatol. 2019, 139, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Shin, K.-O.; Lim, C.-J.; Park, H.-Y.; Kim, S.; Kim, B.; Lee, Y.; Chung, H.; Jeong, S.-K.; Park, K.; Park, K. Activation of SIRT1 enhances epidermal permeability barrier formation through ceramide synthase 2- and 3-dependent mechanisms. J. Investig. Dermatol. 2020, 140, 1435–1438.e5. [Google Scholar] [CrossRef] [PubMed]
- Robles, C.; Garzino, S. Infraspecific variability in the essential oil composition of Cistus monspeliensis leaves. Phytochemistry 2000, 53, 71–75. [Google Scholar] [CrossRef]
- Mastino, P.M.; Marchetti, M.; Costa, J.; Usai, M. Interpopulation variability in the essential oil composition of Cistus creticus subsp. eriocephalus from Sardinia. Chem. Biodivers. 2018, 15, e1800151. [Google Scholar] [CrossRef] [PubMed]
- Ben Jemia, M.; Kchouk, M.E.; Senatore, F.; Autore, G.; Marzocco, S.; De Feo, V.; Bruno, M. Antiproliferative activity of hexane extract from Tunisian Cistus libanotis, Cistus monspeliensis and Cistus villosus. Chem. Cent. J. 2013, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Harada, G.; Pattarawat, P.; Ito, K.; Matsumoto, T.; Hasegawa, T.; Katakura, Y. Lactobacillus brevis T2102 suppresses the growth of colorectal cancer cells by activating SIRT1. J. Funct. Foods 2016, 23, 444–452. [Google Scholar] [CrossRef]
- Kubo, C.; Ogawa, M.; Uehara, N.; Katakura, Y. Fisetin promotes hair growth by augmenting TERT expression. Front. Cell. Dev. Biol. 2020, 8, 566617. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Udono, M.; Kadooka, K.; Yamashita, S.; Katakura, Y. Quantitative analysis of cellular senescence phenotypes using an imaging cytometer. Methods 2012, 56, 383–388. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ledrhem, M.; Nakamura, M.; Obitsu, M.; Hirae, K.; Kameyama, J.; Bouamama, H.; Gadhi, C.; Katakura, Y. Essential Oils Derived from Cistus Species Activate Mitochondria by Inducing SIRT1 Expression in Human Keratinocytes, Leading to Senescence Inhibition. Molecules 2022, 27, 2053. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27072053
Ledrhem M, Nakamura M, Obitsu M, Hirae K, Kameyama J, Bouamama H, Gadhi C, Katakura Y. Essential Oils Derived from Cistus Species Activate Mitochondria by Inducing SIRT1 Expression in Human Keratinocytes, Leading to Senescence Inhibition. Molecules. 2022; 27(7):2053. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27072053
Chicago/Turabian StyleLedrhem, Merieme, Miku Nakamura, Miyu Obitsu, Kinue Hirae, Jun Kameyama, Hafida Bouamama, Chemseddoha Gadhi, and Yoshinori Katakura. 2022. "Essential Oils Derived from Cistus Species Activate Mitochondria by Inducing SIRT1 Expression in Human Keratinocytes, Leading to Senescence Inhibition" Molecules 27, no. 7: 2053. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27072053