
Suppression of Oxidative Stress and Proinflammatory Cytokines Is a Potential Therapeutic Action of Ficus lepicarpa B. (Moraceae) against Carbon Tetrachloride (CCl4)-Induced Hepatotoxicity in Rats


Abstract
:1. Introduction
2. Results
2.1. F. lepicarpa’s In Vitro Antioxidative Activity
2.2. F. lepicarpa Preliminary Phytochemical Screening
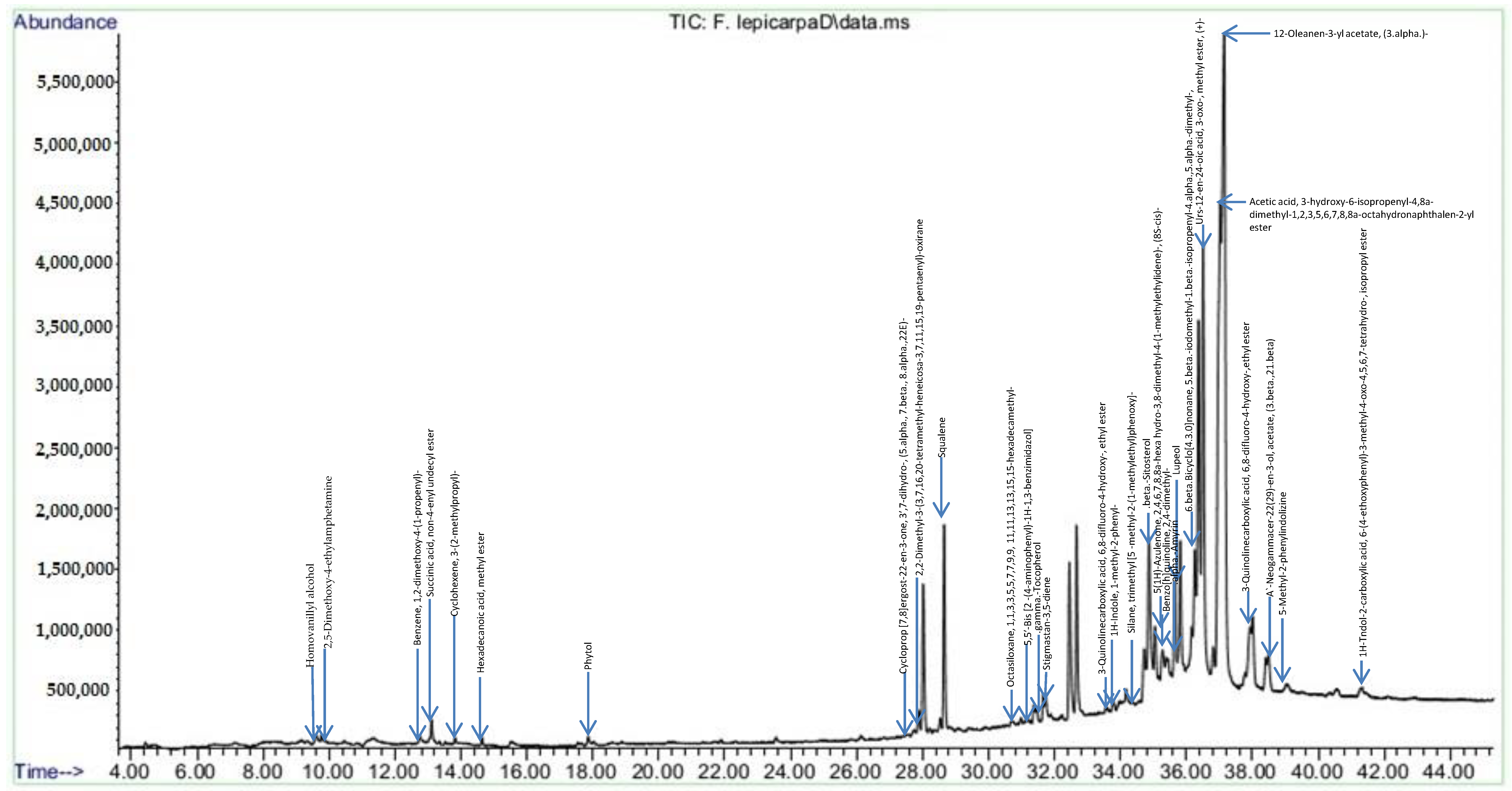
2.3. F. lepicarpa Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
2.4. Liver Index and Body Weight
2.5. F. lepicarpa’s Effect on Serum Transaminases: Alanine Aminotransferase (ALT) and Aspartate Aminotransferase (AST)
2.6. F. lepicarpa’s Effect on Liver Reduced Glutathione (GSH)
2.7. F. lepicarpa’s Effect on Liver Lipid Peroxidation (LPO)
2.8. F. lepicarpa’s Effect on Liver Antioxidant Enzymes: Glutathione Peroxidase (GPx), Glutathione Reductase (GR), Glutathione-S-Transferase (GST) and Quinone Reductase (QR)
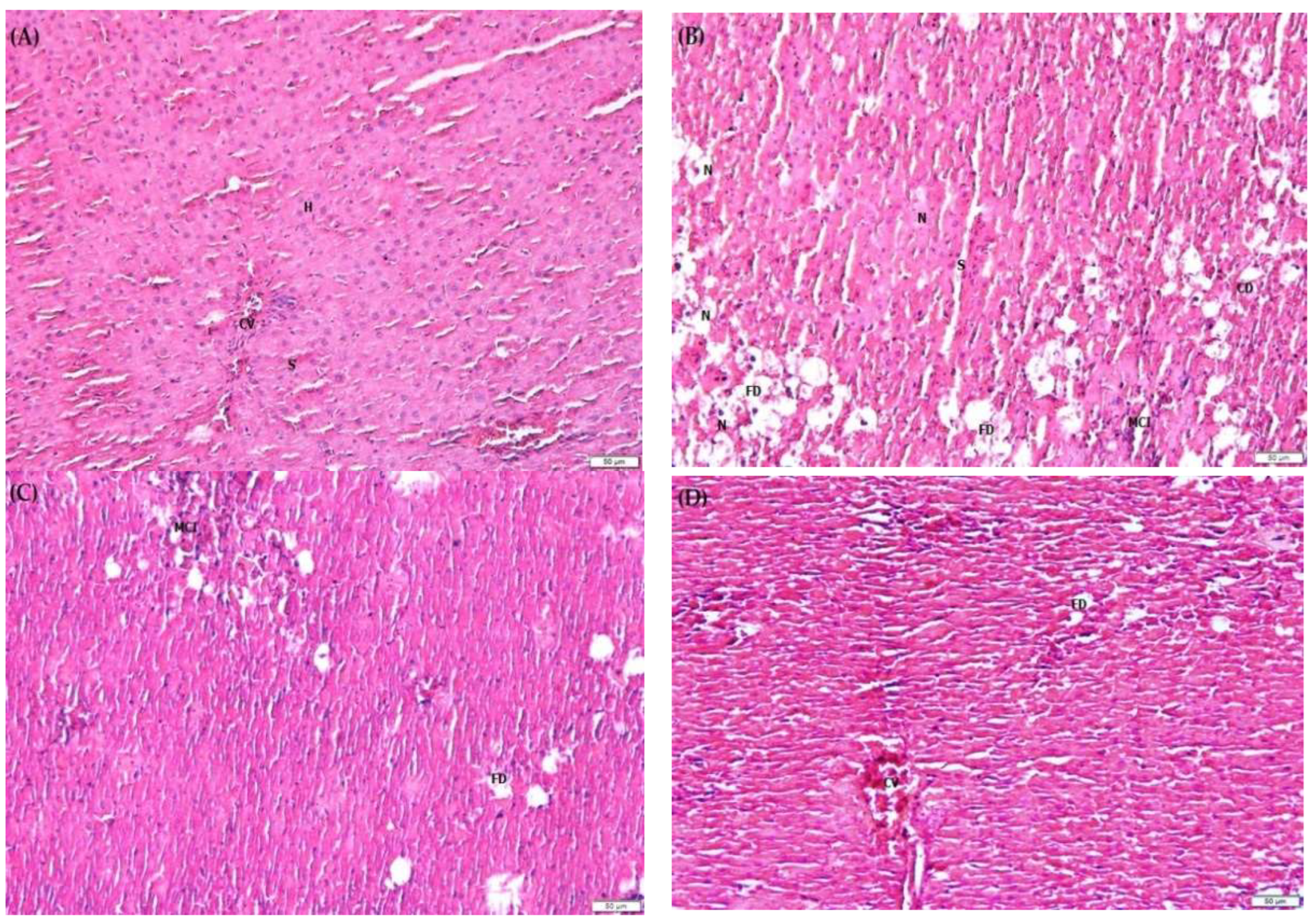
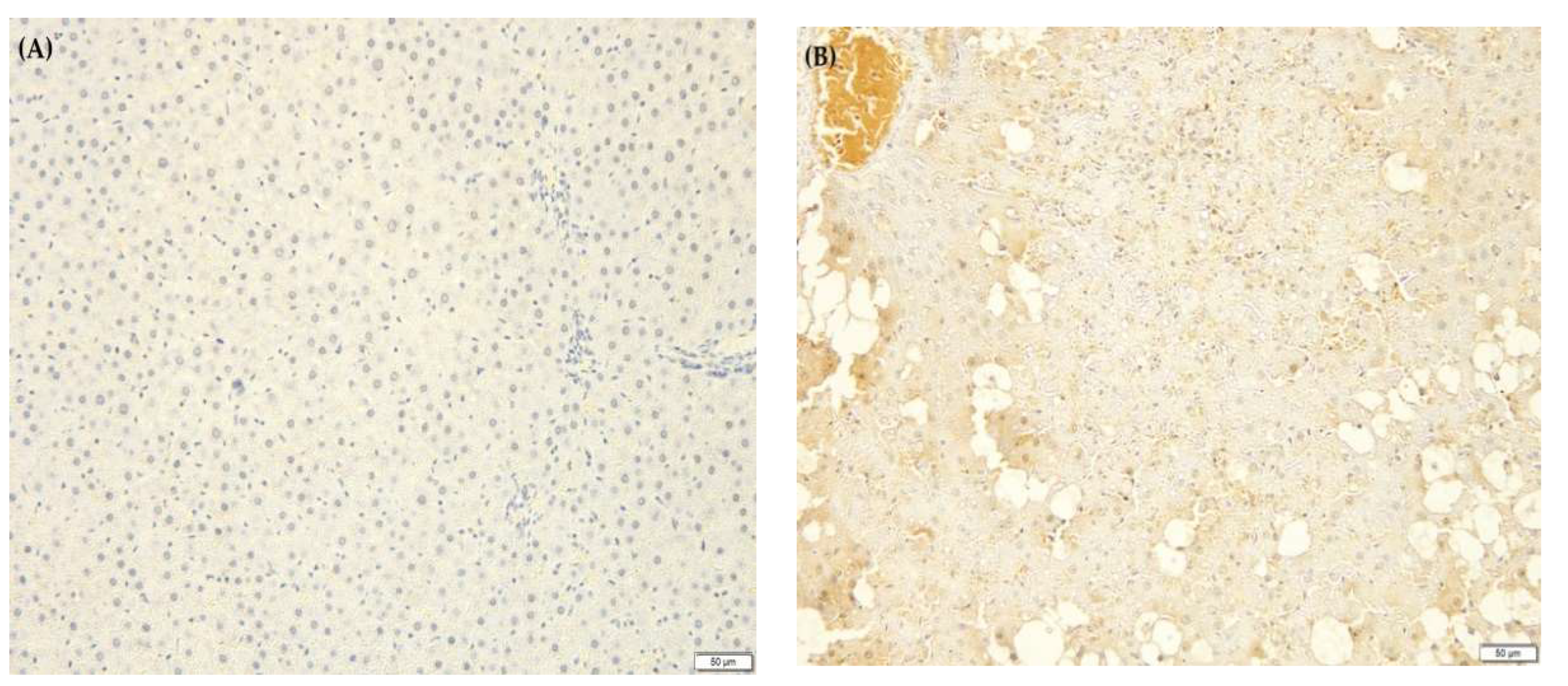
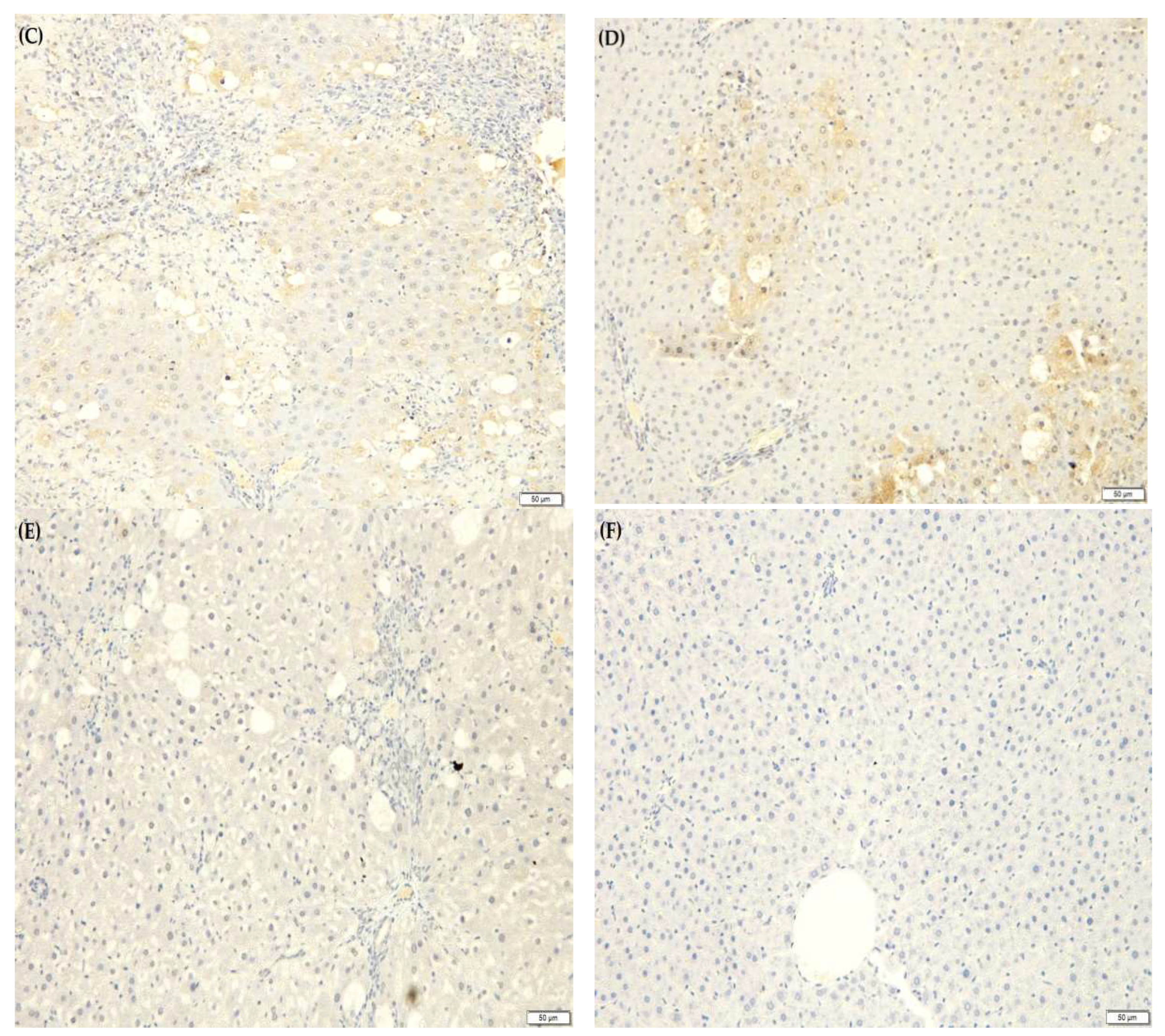
2.9. F. lepicarpa’s Effect on Liver Histopathology
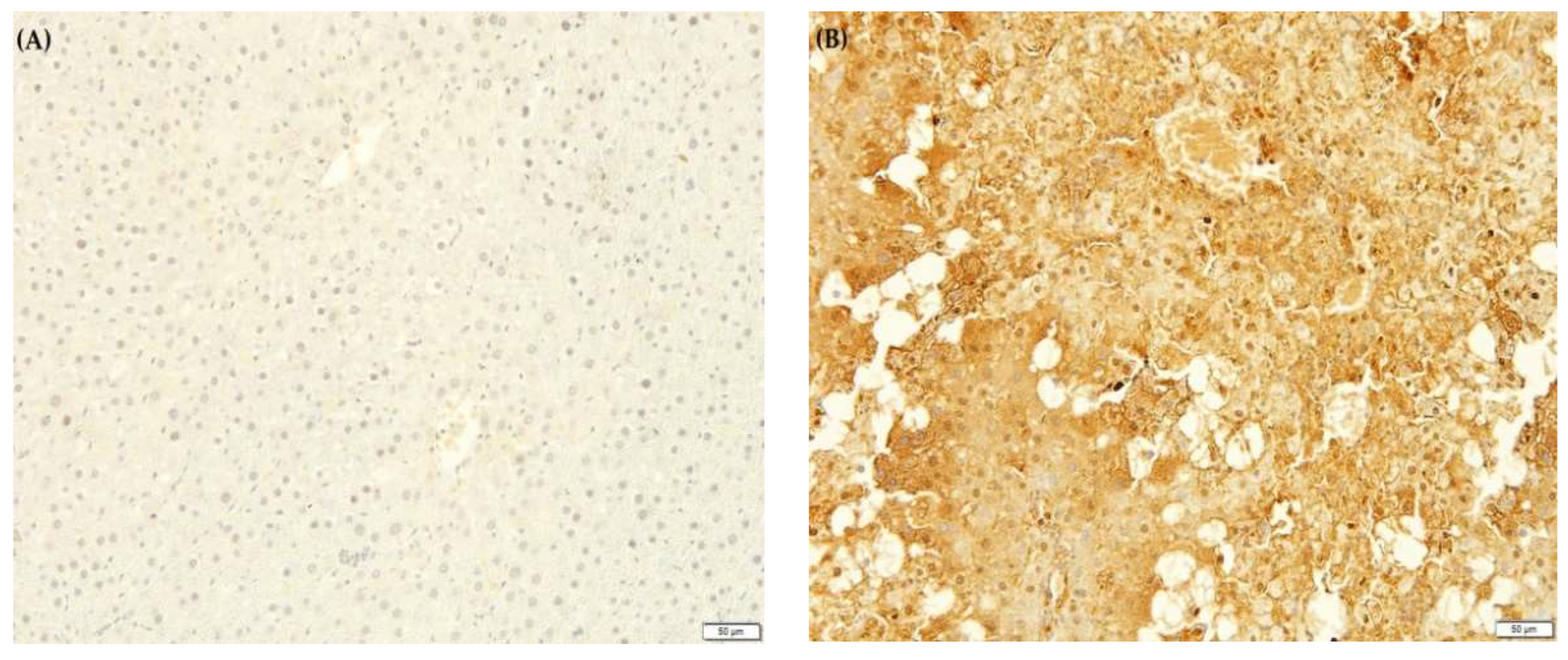
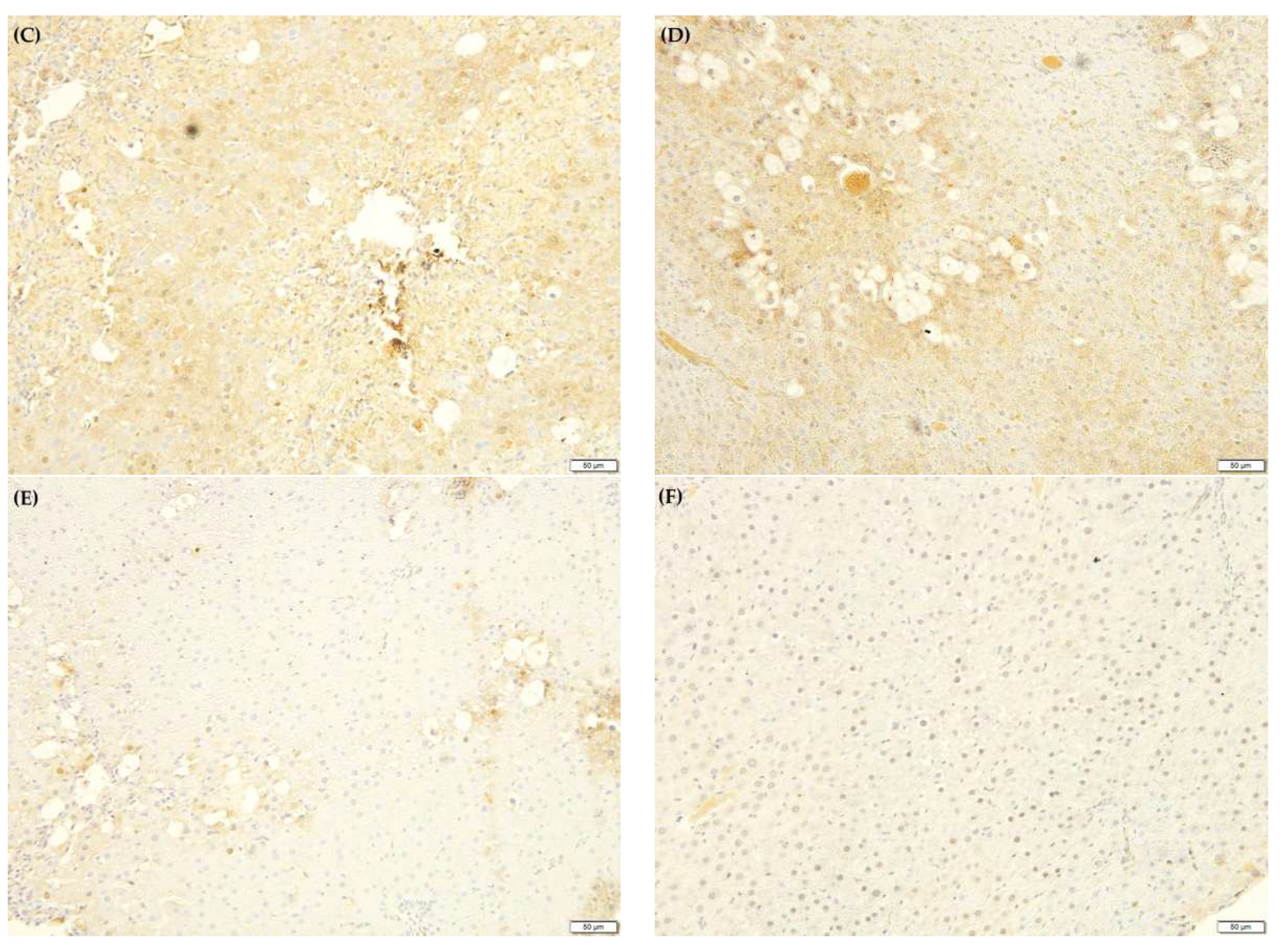
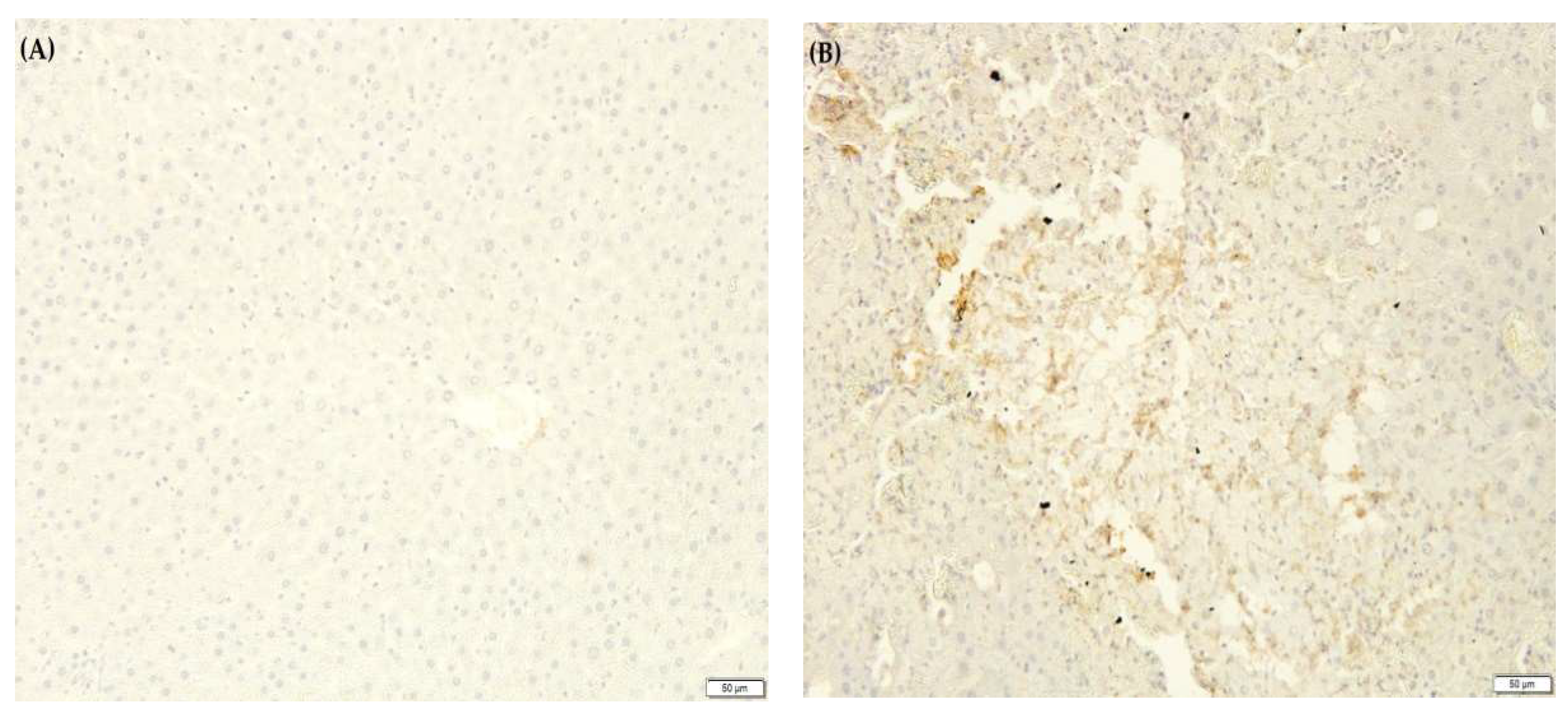
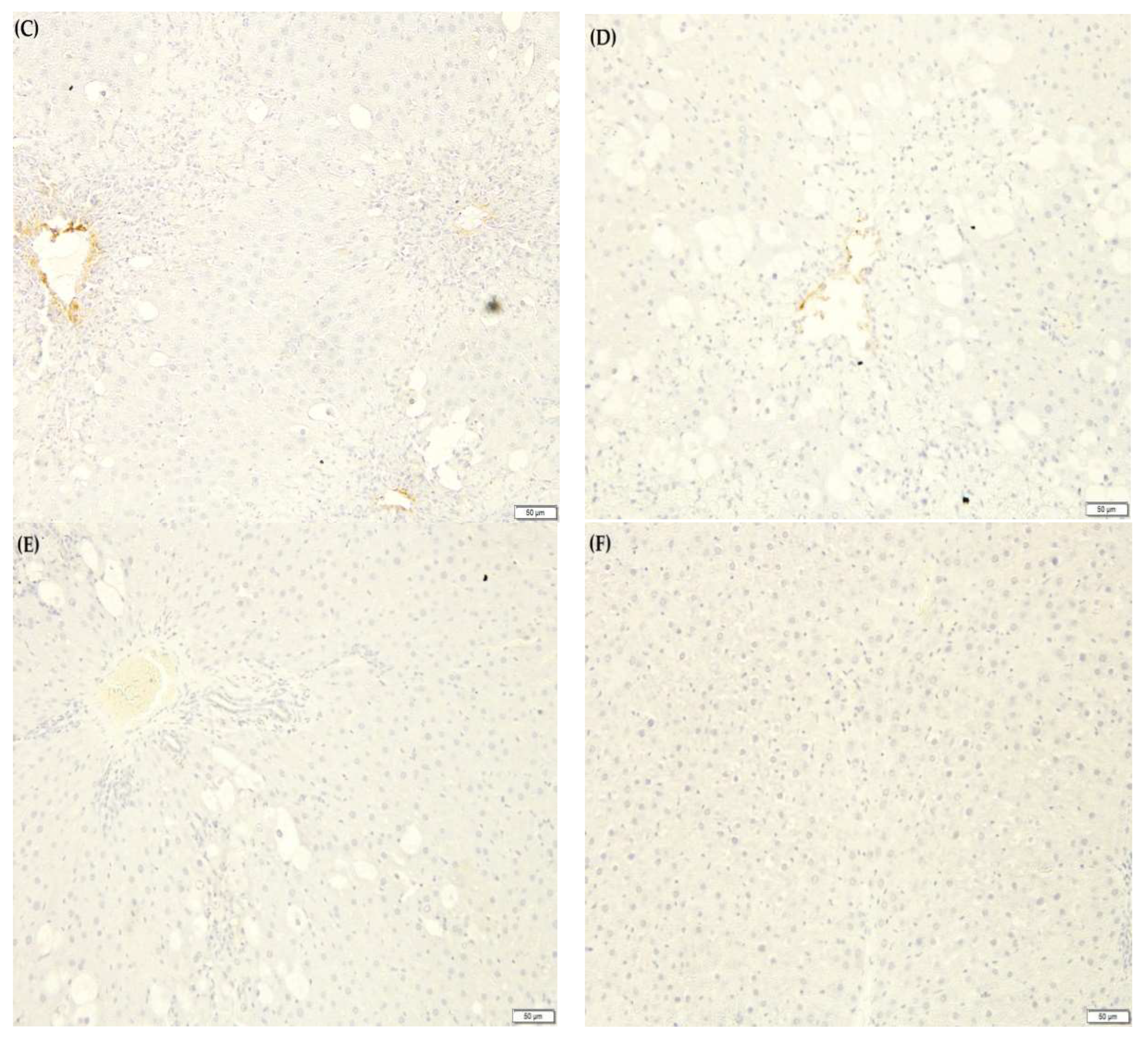
2.10. F. lepicarpa’s Effect on Liver Proinflammatory Cytokines
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Sample Collection and Preparation
4.3. Total Phenolic Content (TPC)
4.4. Total Flavonoid Content (TFC)
4.5. Radical Scavenging Assay (DPPH)
4.6. Phytochemical Screening
4.7. Gas Chromatography-Mass Spectrometry Analysis (GC-MS)
4.8. Animal Experiments
- Group 1: Normal control (were not given any treatment).
- Group 2: CCl4 (1.0 mL/kg bwt).
- Group 3: CCl4 + F. lepicarpa (100 mg/kg bwt).
- Group 4: CCl4 + F. lepicarpa (200 mg/kg bwt).
- Group 5: CCl4 + F. lepicarpa (400 mg/kg bwt).
- Group 6: F. lepicarpa (400 mg/kg bwt) (plant control alone).
4.9. Serum Transaminases (AST and ALT)
4.10. Reduced Glutathione (GSH)
4.11. Lipid Peroxidation (TBARS Content)
4.12. Glutathione Peroxidase (GPx)
4.13. Glutathione Reductase (GR)
4.14. Glutathione S-Transferase (GST)
4.15. Quinone Reductase (QR)
4.16. Histopathological Assessment
4.17. Immunohistochemical Assessment
4.18. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bogdanos, D.P.; Gao, B.; Gershwin, M.E. Liver immunology. Compr. Physiol. 2013, 3, 567–598. [Google Scholar] [PubMed] [Green Version]
- Arias, I.M.; Alter, H.J.; Boyer, J.L.; Cohen, D.E.; Shafritz, D.A.; Thorgeirsson, S.S.; Wolkoff, A.W. The Liver: Biology and Pathobiology, 6th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 3–13. [Google Scholar]
- Thanebal, S.P.; Vun-Sang, S.; Iqbal, M. Hepatoprotective effects of Pandanus amaryllifolius against carbon tetrachloride (CCl4) induced toxicity: A biochemical and histopathological study. Arab. J. Chem. 2021, 14, 103390. [Google Scholar] [CrossRef]
- Asrani, S.K.; Denarbhavi, D.H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Aydin, M.M.; Akcali, K.C. Liver fibrosis. Turk J Gastroenterol. 2018, 29, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Global Health Estimates 2019: Deaths by Cause, Age, Sex, by Country and by Region, 2000–2019; World Health Organization: Geneva, Switzerland, 2020; Available online: https://www.who.int/data/gho/data/themes/mortality-and-global-health-estimates (accessed on 18 August 2021).
- Tsochatzis, E.A.; Bosch, J.; Burroughs, A.K. Liver cirrhosis. Lancet 2014, 383, 1749–1761. [Google Scholar] [CrossRef]
- Shah, M.D.; D’Souza, U.J.A.; Iqbal, M. The potential protective effect of Commelina nudiflora L. against carbon tetrachloride (CCl4)—Induced hepatotoxicity in rats, mediated by suppression of oxidative stress and inflammation. Environ. Health Prev. Med. 2017, 22, 66. [Google Scholar] [CrossRef]
- Ilyas, U.; Katare, D.P.; Aeri, V.; Naseef, P.P. A review on hepatoprotective and immunomodulatory herbal plants. Pharmacogn. Rev. 2016, 10, 66–70. [Google Scholar]
- Shirani, M.; Raeisi, R.; Heidari-Soureshjani, S.; Asadi-Samani, M.; Luther, T. A review for discovering hepatoprotective herbal drugs with least side effects on kidney. J. Nephropharmacology 2017, 6, 38–48. [Google Scholar] [CrossRef]
- Mohamed Saleem, T.S.; Madhusudhana Chetty, C.; Ramkanth, S.; Rajan, V.S.T.; Mahesh Kumar, K.; Gauthaman, K. Hepatoprotective herbs—A review. Int. J. Res. Pharm. Sci. 2010, 1, 1–5. [Google Scholar]
- Iqbal, M.; Shah, M.D.; Vun-Sang, S.; Awang Saman, R. Hepatoprotective activity of Merremia borneensis against carbon tetrachloride (CCl4)–Induced acute liver damage in rats: A biochemical and histopathological evaluation. Int. J. Pharm. Pharm. Sci. 2020, 12, 41–45. [Google Scholar] [CrossRef]
- Corner, E.J.H. New Taxa of Ficus (Moraceae) 2. Blumea 1975, 22, 299–309. [Google Scholar]
- Berg, C.C.; Corner, E.J.H. Flora Malesiana, Series I: Moraceae–Ficus. In Flora Malesiana, Series I: Spermatophyta (Seed Plants); Nooteboom, H.P., Ed.; National Herbarium of Netherlands: Leiden, The Netherlands, 2005; Volume 17/2, pp. 393–431. [Google Scholar]
- Jansen, P.C.M.; Jukema, J.; Oyen, L.P.A.; van Lingen, T.G. Ficus lepicarpa Blume. In Plant Resources of South-East Asia No 2: Edible Fruits and Nuts; Verheij, E.W.M., Coronel, R.E., Eds.; PROSEA Foundation: Bogor, Indonesia, 1991; Available online: https://prota4u.org/prosea/view.aspx?id=1656 (accessed on 28 November 2021).
- Milow, P.; Malek, S.B.; Edo, J.; Ong, H.C. Malaysian Species of Plants with Edible Fruits or Seeds and Their Valuation. Int. J. Fruit Sci. 2014, 14, 1–27. [Google Scholar] [CrossRef]
- Ahmad, F.B.; Holdsworth, D.K. Traditional medicinal plants of Sabah, Malaysia—Part III. The Rungus People of Kudat. Int. J. Pharmacogn. 1995, 33, 262–264. [Google Scholar] [CrossRef]
- Vun-Sang, S.; Teoh, P.L.; Rodriques, K.F.; Iqbal, M. Phytochemical analysis and antioxidant activity of Ficus lepicarpa leaves from Sabah, Malaysia. Trans. Sci. Technol. 2017, 4, 360–365. [Google Scholar]
- Kulip, J. An enthnobotanical survey of medicinal and other useful plants of Muruts in Sabah, Malaysia. Telopea 2003, 10, 81–98. [Google Scholar] [CrossRef]
- Ibrahim, F.H.; Hamzah, N.H. The use of medicinal plant species by the Temuan Tribe of Ayer Hitam Forest, Selangor, Peninsular Malaysia. J. Trop. Agric. Sci. 1999, 22, 85–94. [Google Scholar]
- Pandit, D.N.; Priyanka; Gupta, M.L. Heapto-Somatic Index, Gonado-Somatic Index and Condition Factor of Anabas testudineus as bio-monitoring tools of nickel and chromium toxicity. Int. J. Innov. Eng. Technol. (IJIET) 2019, 12, 25–28. [Google Scholar]
- Rao, J.P. A novel and economic method to assess clinical transaminase assays. Indian J. Appl. Res. 2014, 4, 44–46. [Google Scholar] [CrossRef]
- Prabha, D.C.; Annapoorani, S. Hepatoprotective effect of Cynodon dactylon on CCl4 induced experimental mice. J. Biol. Sci. 2009, 17, 17–24. [Google Scholar]
- Liu, F.; Ong, E.S.; Li, Y.S.F. Drying is generally regarded as the most important step in the post-harvest process due to its importance in limiting enzymatic degradation and microbial growth while preserving the plant’s beneficial properties. Food Chem. 2013, 141, 1807–1813. [Google Scholar] [CrossRef]
- Zhu, Y.; Pu, B.Q.; Xie, G.Y.; Tian, M.; Chen, Y.J.; Qin, M.J. Dynamic changes of flavonoids content in the different parts of rhizome of Belamcanda chinensis during thermal drying process. Molecules 2014, 19, 10440–10454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Hao, L.J.; Wu, G.; Wang, S.; Duan, J.A.; Xie, G.Y.; Qin, M.J. Effects of drying methods on the phytochemicals contents and antioxidant properties of chrysanthemum flower heads harvested at two developmental stages. J. Funct. Foods 2015, 19, 786–795. [Google Scholar] [CrossRef]
- Tarvainen, M.; Suomela, J.P.; Kallio, H.; Yang, B. Triterpene acids in plantago major. Identification, quantification and comparison of different extraction methods. Chromatographia 2010, 71, 279–284. [Google Scholar] [CrossRef]
- Wójciak-Kosior, M.; Sowa, I.; Kocjan, R.; Nowak, R. Effect of different extraction techniques on quantification of oleanolic and ursolic acid in Lamii albi flos. Ind. Crops Prod. 2013, 44, 373–377. [Google Scholar] [CrossRef]
- Bae, H.; Jayaprakasha, G.K.; Jifon, J.; Patil, B.S. Variation of antioxidant activity and the levels of bioactive compounds in lipophilic and hydrophilic extracts from hot pepper (Capsicum spp.) cultivars. Food Chem. 2012, 134, 1912–1918. [Google Scholar] [CrossRef]
- Khan, J.; Deb, P.K.; Priya, S.; Medina, K.D.; Devi, R.; Walode, S.G.; Rudrapal, M. Dietary Flavonoids: Cardioprotective potential with antioxidant effects and their pharmacokinetic, toxicological and therapeutic concerns. Molecules 2021, 26, 4021. [Google Scholar] [CrossRef]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total phenolic content, flavonoid content and antioxidant potential of wild vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Sang, W.; Zhou, M.; Ren, G. Phenolic composition and antioxidant activities of 11 celery cultivars. J. Food Sci. 2010, 75, C9–C13. [Google Scholar] [CrossRef]
- Hossain, M.A.; Nagooru, M.R. Biochemical profiling and total flavonoids contents of leaves crude extract of endemic medical plant Cordydyline terminals L. Kunth. Pharmacogn. J. 2011, 3, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Dillard, C.J.; German, J.B. Phytochemicals: Nutraceuticals and human health. J. Sci. Food Agric. 2000, 80, 1744–1756. [Google Scholar] [CrossRef]
- Vilegas, J.H.Y.; Lanças, F.M.; Vilegas, W.; Pozetti, G.L. Further triterpenes, steroids and furocoumarins from Brazilian medicinal plants of Dorstenia genus (Moraceae). J. Braz. Chem. Soc. 1997, 8, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Abubakar, U.S.; Danmalam, U.H.; Ahmed, A.; Abdullahi, S.; Abba, A.; Rukayya, N. GC-MS analysis of ethyl acetate extract of Ficus sycomorus root Linn. (Moraceae). Bayero J. Pure Appl. Sci. (BAJOPAS) 2016, 9, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, F.A.; Chaves, M.H.; Almeida, F.R.; Lima, R.C.J.; Silva, R.M.; Maia, J.L.; Brito, G.A.; Santos, F.A.; Rao, V.S. Protective effect of alpha- and beta- amyrin, a triterpene mixture from Protium heptaphyllum (Aubl.) March. Trunk wood resin, against acetaminophen-induced liver injury in mice. J. Ethnopharmacolology 2005, 98, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Okoye, N.N.; Ajaghaku, D.L.; Okeke, H.N.; Ilodigwe, E.E.; Nworu, C.S.; Okoye, F.B. Beta-Amyrin and alpha-Amyrin acetate isolated from the stem bark of Alstinia boonei display profound anti-inflammatory activity. Pharm. Biol. 2014, 52, 1478–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, L.W.; Boll, M.; Stamfl, A. Hepatotoxicity and mechanism of action of haloalkanes: Carbon tetrachloride as a toxicological model. Crit. Rev. Toxicol. 2003, 33, 105–136. [Google Scholar] [CrossRef]
- Eidi, A.; Moghadam, J.Z.; Mortazavi, P.; Rezazadeh, S.; Olamafar, S. Hepatoprotective effects of Juglans regia extract against CCl4-induced oxidative damage in rats. Pharm. Biol. 2013, 51, 558–565. [Google Scholar] [CrossRef] [Green Version]
- Seyer, J.M.; Hutcheson, E.T.; Kang, A.H. Collagen polymorphism in normal and cirrhotic human liver. J. Clin. Investig. 1977, 59, 241–248. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, Z.J.; Lao, L.; Wong, C.W.; Feng, Y. The role of oxidative stress and antioxidants in liver diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Dey, A.; Dutta, R.; Dey, S.; Acharya, K. Hepatoprotective effect of the ethanolic extract of Calocybe indica on mice with CCl4 hepatic intoxication. Int. J. PharmTech Res. 2011, 3, 2162–2168. [Google Scholar]
- Shah, M.D.; Gnanaraj, C.; Emdadul Haque, A.T.M.; Iqbal, M. Antioxidative and chemopreventive effects of Nephrolepis biserrata against carbon tetrachloride (CCl4)-induced oxidative stress and hepatic dysfunction in rats. Pharm. Biol. 2015, 53, 31–39. [Google Scholar] [CrossRef]
- Gnanaraj, C.; Shah, M.D.; Emdadul Haque, A.T.M.; Makki, J.S.; Iqbal, M. Hepatoprotective and immunosuppressive effect of Synedrella nodiflora L. on carbon tetrachloride (CCl4)-intoxicated rats. J. Environ. Pathol. Toxicol. Oncol. 2016, 35, 29–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amzar, N.; Iqbal, M. The Hepatoprotective effect of Clidemia hirta against carbon tetrachloride (CCl4)–induced oxidative stress and hepatic damage in mice. J. Environ. Pathol. Toxicol. Oncol. 2017, 36, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.D.; Gnanaraj, C.; Khan, M.S.; Iqbal, M. Dillenia suffruticosa L. impedes carbon tetrachloride-induced hepatic damage by modulating oxidative stress and inflammatory markers in rats. J. Environ. Pathol. Toxicol. Oncol. 2015, 34, 133–152. [Google Scholar] [CrossRef] [Green Version]
- Gnanaraj, C.; Shah, M.D.; Song, T.T.; Iqbal, M. Hepatoprotective mechanism of Lygodium microphyllum (Cav.) R.Br. through ultrastructural signalling prevention against carbon tetrachloride (CCl4)-mediated oxidative stress. Biomed. Pharmacother. 2017, 92, 1010–1022. [Google Scholar] [CrossRef]
- Fard, J.K.; Hamzely, H.; Sattari, M.; Eftkhari, A.; Ahmadian, E.; Eghbal, M.A. Triazole rizatriptan induces liver toxicity through lysosomal/mitochondrial dysfuction. Drug Res. 2016, 66, 470–478. [Google Scholar]
- Ahmadian, E.; Eftekhari, A.; Fard, J.K.; Babaei, H.; Nayebi, A.M.; Mohammadnejad, D.; Eghbal, M.A. In vitro and in vivo evaluation of the mechanisms of citalopram-induced hepatotoxicity. Arch. Pharmacal Res. 2017, 40, 1296–1313. [Google Scholar] [CrossRef] [PubMed]
- Eftekhari, A.; Ahmadian, E.; Azarmi, Y.; Parvizpur, A.; Fard, J.K.; Eghbal, M.A. The effects of cimetidine, n-acetylcysteine, and taurine on thioridazine metabolic activation and induction of oxidative stress in isolated rat hepatocytes. Pharm. Chem. J. 2018, 51, 965–969. [Google Scholar] [CrossRef]
- Macdonald, I.O.; Olusola, O.J.; Osaigbovo, U.A. Effects of chronic ethanol administration on body weight, reduced glutathione (GSH), malondialdehyde (MDA) levels and glutathione-s-transferase activity (GST) in rats. N. Y. Sci. J. 2010, 3, 39–47. [Google Scholar]
- Rehman, M.U.; Tahir, M.; Khan, A.Q.; Khan, R.; Lateef, A.; Hamiza, O.O.; Qamar, W.; Ali, F.; Sultana, S. Chrysin suppresses renal carcinogenesis via amelioration of hyperproliferation, oxidative stress and inflammation: Plausible role of NF-kb. Toxicol. Lett. 2013, 216, 146–158. [Google Scholar] [CrossRef]
- Knockaert, L.; Berson, A.; Ribault, C.; Prost, P.E.; Fautrel, A.; Pajaud, J.; Lepage, S.; Lucas-Clerc, C.; Bégué, J.M.; Fromenty, B.; et al. Carbon tetrachloride-mediated lipid peroxidation induces early mitochondrial alterations in mouse liver. Lab. Investig. 2012, 92, 396–410. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.Y.; Chao, J.; Chiu, C.S.; Hsieh, I.C.; Huang, H.C.; Wu, L.Y.; Peng, W.H. Hepatoprotective and antioxidant effects of Wu-Zi-Yuan-Chung-Wan against CCl4-induced oxidative damage in rats. Eur. J. Inflamm. 2021, 19, 1–13. [Google Scholar] [CrossRef]
- Althnaian, T.; Albokhadaim, I.; El-Bahr, S.M. Biochemical and histopathological study in rats intoxicated with carbon tetrachloride and treated with camel milk. Springer Plus 2013, 2, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Muñoz, I.; de la Torre, P.; Sánchez-Alcázar, J.A.; García, I.; Santiago, E.; Muñoz-Yagüe, M.T.; Solís-Herruzo, J.A. Tumor necrosis factor alpha inhibits collagen alpha 1(I) gene expression in rat hepatic stellate cells through a G protein. Gastroenterology 1997, 113, 625–640. [Google Scholar] [CrossRef]
- Yang, T.; Li, Y.; Lyu, Z.; Huang, K.; Corrigan, C.J.; Ying, S.; Wang, W.; Wang, C. Characteristics of proinflammatory cytokines and chemokines in airways of asthmatics: Relationships with disease severity and infiltration of inflammatory cells. Chin. Med. J. 2017, 130, 2033–2040. [Google Scholar] [CrossRef] [PubMed]
- Jantan, I.; Jumuddin, F.A.; Saputri, F.C.; Rahman, K. Inhibitory effects of the extracts of Garcinia species on human low density lipoprotein peroxidation and platelet aggregation in relation to their total phenolic contents. J. Med. Plants Res. 2014, 5, 2699–2709. [Google Scholar]
- Velioglu, Y.S.; Mazza, G.; Gao, L.; Oomah, B.D. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Zou, Y.P.; Lu, Y.H.; Wei, D.Z. Antioxidant activity of flavonoid-rich extracts of Hypericum perforatum L. in-vitro. J. Agric. Food Chem. 2004, 52, 5032–5039. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Vimalkumar, C.S.; Hosagaudar, V.B.; Suja, S.R.; Vilash, V.; Krishnakumar, N.M.; Latha, P.G. Comparative preliminary phytochemical analysis of ethanolic extracts of leaves of Olea dioica Roxb., infected with the rust fungus Zaghouania oleae (E.J. Butler) Cummins and non-infected plants. J. Pharmacogn. Phytochem. 2014, 3, 69–72. [Google Scholar]
- Hossain, M.A.; AL-Raqmi, K.A.; AL-Mijizy, Z.H.; Weli, A.M.; Al-Riyami, Q. Study of total phenol, flavonoids contents and phytochemical screening of various leaves crude extracts of locally grown Thymus vulgaris. Asian Pac. J. Trop. Biomed. 2013, 3, 705–710. [Google Scholar] [CrossRef] [Green Version]
- Ugochukwu, S.C.; Uche, I.A.; Ifeanyi, O. Preliminary phytochemical screening of different solvent extracts of stem bark and roots of Dennetia tripetala G. Baker. Asian J. Plant Sci. Res. 2013, 3, 10–13. [Google Scholar]
- Firdouse, S.; Alam, P. Phytochemical investigation of extract of Amorphophallus campanulatus tubers. Int. J. Phytomed. 2011, 3, 32–35. [Google Scholar]
- Philip, D.; Kaleena, P.K.; Valivittan, K.; Girish Kumar, C.P. Phytochemical screening and antimicrobial activity of Sansevieria roxburghiana Schult. and Schult. F. Middle-East J. Sci. Res. 2011, 10, 512–518. [Google Scholar]
- Harborne, J.B. Phytochemical methods. In A Guide to Modern techniques of Plant Analysis, 3rd ed.; Chapman and Hall: London, UK, 1998; pp. 100–101. [Google Scholar]
- Clark, J.D.; Gebhart, G.F.; Gonder, J.C.; Keeling, M.E.; Kohn, D.F. Special Report: The 1996 Guide for the Care and Use of Laboratory Animals. ILAR (Inst. Lab. Anim. Resour.) J. 1997, 38, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum oxaloacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef]
- Jollow, D.J.; Mitchell, J.R.; Zampaglione, N.; Gillette, J.R. Bromobenzene-induced liver necrosis. Protective role of glutathione and evidence for 3, 4-bromobenzene oxide as the hepatotoxic metabolite. Pharmacology 1974, 11, 151–169. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar]
- Iqbal, M.; Giri, U.; Giri, D.K.; Alam, M.S.; Athar, M. Age-dependent renal accumulation of 4-hydroxy-2-nonenal (HNE)-modified proteins following parenteral administration of ferric nitrilotriacetate commensurate with its differential toxicity: Implications for the involvement of HNE-protein adducts in oxidative stress and carcinogenesis. Arch. Biochem. Biophys. 1999, 365, 101–112. [Google Scholar]
- Mohandas, J.; Marshall, J.J.; Duggin, G.G.; Horvath, J.S.; Tiller, D.J. Differential distribution of glutathione and glutathione-related enzymes in rabbit kidney. Possible implications in analgesic nephropathy. Biochem. Pharmacol. 1984, 33, 1801–1807. [Google Scholar] [CrossRef]
- Iqbal, M.; Rezazadeh, H.; Ansar, S.; Athar, M. Alpha-Tocopherol (vitamin-E) ameliorates ferric nitrilotriacetate (Fe-NTA)-dependent renal proliferative response and toxicity: Diminution of oxidative stress. Hum. Exp. Toxicol. 1998, 17, 163–171. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. Purification and characterization of the flavoenzyme glutathione reductase from rat liver. J. Biol. Chem. 1975, 250, 5475–5480. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Athar, M.; Iqbal, M. Ferric nitrilotriacetate promotes N-diethylnitrosamine-induced renal tumorigenesis in the rat: Implications for the involvement of oxidative stress. Carcinogenesis 1998, 19, 1133–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, A.M.; Hunkeler, A.J.; Talalay, P. Increase of NAD(P)H: Quinone reductase by dietary antioxidants: Possible role in protection against carcinogenesis and toxicity. Proc. Natl. Acad. Sci. USA 1980, 77, 5216–5220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, M.; Sharma, S.D.; Rahman, A.; Trikha, P.; Athar, M. Evidence that ferric nitrilotriacetate mediates oxidative stress by down-regulating DT-diaphorase activity: Implications for carcinogenesis. Cancer Lett. 1999, 141, 151–157. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemical Constituents | Presence (+) or Absence (−) |
|---|---|
| Alkaloids (Wagner’s test) | − |
| Flavonoids (alkaline reagent test) | + |
| Tannins (Braymer’s test) | − |
| Saponin (foam test) | + |
| Phenols (ferric chloride test) | + |
| Steroids (Liebermann–Burchard test) | + |
| Anthraquinones | − |
| Phytosterols | + |
| Triterpenoids (Salkowki’s test) | + |
| No. | Ret. Time (min) | Compound | Area (%) |
|---|---|---|---|
| 1. | 9.595 | Homovanillyl alcohol | 0.24 |
| 2. | 9.773 | 2,5-Dimethoxy-4-ethylamphetamine | 0.16 |
| 3. | 12.756 | Benzene, 1,2-dimethoxy-4-(1-propenyl)- | 0.16 |
| 4. | 13.111 | Succinic acid, non-4-enyl undecyl ester | 0.42 |
| 5. | 13.832 | Cyclohexene, 3-(2-methylpropyl)- | 0.07 |
| 6. | 14.637 | Hexadecanoic acid, methyl ester | 0.11 |
| 7. | 17.865 | Phytol | 0.13 |
| 8. | 27.762 | Cycloprop [7,8]ergost-22-en-3-one, 3′,7-dihydro-, (5.alpha., 7.beta., 8.alpha.,22E)- | 0.13 |
| 9. | 27.880 | 2,2-Dimethyl-3-(3,7,16,20-tetramethyl-heneicosa-3,7,11,15,19-pentaenyl)-oxirane | 0.44 |
| 10. | 28.660 | Squalene | 2.85 |
| 11. | 30.727 | Octasiloxane, 1,1,3,3,5,5,7,7,9,9, 11,11,13,13,15,15-hexadecamethyl- | 0.16 |
| 12. | 31.176 | 5,5′-Bis [2 -(4-aminophenyl)-1H-1,3-benzimidazol] | 0.18 |
| 13. | 31.371 | .gamma.-Tocopherol | 0.27 |
| 14. | 31.736 | Stigmastan-3,5-diene | 0.55 |
| 15. | 33.778 | 3-Quinolinecarboxylic acid, 6,8-difluoro-4-hydroxy-, ethyl ester | 0.20 |
| 16. | 33.981 | 1H-Indole, 1-methyl-2-phenyl- | 0.19 |
| 17. | 34.083 | Silane, trimethyl [5 -methyl-2-(1-methylethyl)phenoxy]- | 0.167 |
| 18. | 34.862 | .beta.-Sitosterol | 4.44 |
| 19. | 35.159 | 5(1H)-Azulenone, 2,4,6,7,8,8a-hexa hydro-3,8-dimethyl-4-(1-methylethylidene)-, (8S-cis)- | 0.46 |
| 20. | 35.278 | Benzo[h]quinoline, 2,4-dimethyl- | 1.67 |
| 21. | 35.650 | .alpha.-Amyrin | 2.62 |
| 22. | 35.820 | Lupeol | 4.16 |
| 23. | 36.260 | 6.beta.Bicyclo[4.3.0]nonane, 5.beta.-iodomethyl-1.beta.-isopropenyl-4.alpha.,5.alpha.-dimethyl-, | 3.02 |
| 24. | 36.506 | Urs-12-en-24-oic acid, 3-oxo-, methyl ester, (+)- | 9.80 |
| 25. | 37.032 | Acetic acid, 3-hydroxy-6-isopropenyl-4,8a-dimethyl-1,2,3,5,6,7,8,8a-octahydronaphthalen-2-yl ester | 13.69 |
| 26. | 37.150 | 12-Oleanen-3-yl acetate, (3.alpha.)- | 21.09 |
| 27. | 38.006 | 3-Quinolinecarboxylic acid, 6,8-difluoro-4-hydroxy-,ethyl ester | 1.75 |
| 28. | 38.489 | A`-Neogammacer-22(29)-en-3-ol, acetate, (3.beta.,21.beta) | 0.77 |
| 29. | 39.048 | 5-Methyl-2-phenylindolizine | 0.06 |
| 30. | 41.302 | 1H-Tndol-2-carboxylic acid, 6-(4-ethoxyphenyl)-3-methyl-4-oxo-4,5,6,7-tetrahydro-, isopropyl ester | 0.15 |
| Treatment Group | Initial Body Weight (g) | Final Body Weight (g) | Percentage Increase of Body Weight (%) | Liver Index (%) |
|---|---|---|---|---|
| Control | 227.77 ± 6.61 | 265.33 ± 13.63 | 16.49 | 3.08 ± 0.29 |
| CCl4 (1 mL/kg bwt) | 229.17 ± 7.24 | 237.50 ± 2.55 | 3.64 | 4.93 ± 0.37 |
| F. lepicarpa (100 mg/kg bwt + CCl4) | 228.63 ± 5.11 | 241.81 ± 2.93 | 5.76 | 4.15 ± 0.29 |
| F. lepicarpa (200 mg/kg bwt + CCl4) | 230.83 ± 5.75 | 247.77 ± 1.98 | 7.34 | 4.10 ± 0.20 |
| F. lepicarpa (400 mg/kg bwt + CCl4) | 232.58 ± 6.41 | 252.33 ± 3.11 | 8.49 | 4.03 ± 0.29 |
| F. lepicarpa (400 mg/kg bwt) (plant control) | 229.67 ± 7.20 | 265.75 ± 14.04 | 15.71 | 3.05 ± 0.23 |
| Treatment Group | ALT Enzyme Activity (U/L) | AST Enzyme Activity (U/L) |
|---|---|---|
| Control | 6.56 ± 0.06 | 11.58 ± 0.01 |
| CCl4 (1 mL/kg bwt) | 47.03 ± 0.33 * | 42.28 ± 0.59 * |
| F. lepicarpa (100 mg/kg bwt + CCl4) | 23.75 ± 0.41 | 24.20 ± 0.33 |
| F. lepicarpa (200 mg/kg bwt + CCl4) | 14.63 ± 0.37 ** | 19.21 ± 0.34 |
| F. lepicarpa (400 mg/kg bwt + CCl4) | 11.26 ± 0.06 ** | 14.41 ± 0.02 ** |
| F. lepicarpa (400 mg/kg bwt) (plant control) | 6.64 ± 0.07 ** | 11.21 ± 0.02 ** |
| Treatment Group | GSH (µmol/g Tissue) | MDA Formation (nmol MDA/g Tissue) |
|---|---|---|
| Control | 9.18 ± 0.05 | 22.22 ± 0.01 |
| CCl4 (1 mL/kg bwt) | 2.33 ± 0.04 * | 69.51 ± 0.12 * |
| F. lepicarpa (100 mg/kg bwt + CCl4) | 3.87 ± 0.04 | 59.18 ± 0.04 |
| F. lepicarpa (200 mg/kg bwt + CCl4) | 4.03 ± 0.05 ** | 49.34 ± 0.07 |
| F. lepicarpa (400 mg/kg bwt + CCl4) | 5.41 ± 0.07 ** | 36.74 ± 0.02 ** |
| F. lepicarpa (400 mg/kg bwt) (plant control) | 8.32 ± 0.05 ** | 21.31 ± 0.02 ** |
| Treatment Group | GPx (nmol NADPH Oxidized/min/mg Protein) | GR (nmol NADPH Oxidized/min/mg Protein) | GST (nmol CDNB Conjugate Formed/min/mg Protein) | QR (nmol Dichloroindophenol Reduced/min/mg Protein) |
|---|---|---|---|---|
| Control | 865.25 ± 0.14 | 215.46 ± 0.30 | 330.18 ± 0.04 | 114.06 ± 0.12 |
| CCl4 (1 mL/kg bwt) | 389.77 ± 0.20 * | 71.71 ± 0.24 * | 111.85 ± 0.04 * | 59.08 ± 0.10 * |
| F. lepicarpa (100 mg/kg bwt + CCl4) | 400.86 ± 0.11 | 104.86 ± 0.25 | 177.68 ± 0.08 | 80.89 ± 0.11 |
| F. lepicarpa (200 mg/kg bwt + CCl4) | 523.58 ± 0.14 ** | 145.89 ± 0.28 ** | 222.04 ± 0.04 ** | 100.30 ± 0.11 ** |
| F. lepicarpa (400 mg/kg bwt + CCl4) | 542.32 ± 0.06 ** | 182.36 ± 0.35 ** | 249.04 ± 0.03 ** | 102.04 ± 0.12 ** |
| F. lepicarpa (400 mg/kg bwt) (plant control) | 760.50 ± 0.06 ** | 203.84 ± 0.27 ** | 296.49 ± 0.02 ** | 106.38 ± 0.12 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vun-Sang, S.; Rodrigues, K.F.; Dsouza, U.J.A.; Iqbal, M. Suppression of Oxidative Stress and Proinflammatory Cytokines Is a Potential Therapeutic Action of Ficus lepicarpa B. (Moraceae) against Carbon Tetrachloride (CCl4)-Induced Hepatotoxicity in Rats. Molecules 2022, 27, 2593. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27082593
Vun-Sang S, Rodrigues KF, Dsouza UJA, Iqbal M. Suppression of Oxidative Stress and Proinflammatory Cytokines Is a Potential Therapeutic Action of Ficus lepicarpa B. (Moraceae) against Carbon Tetrachloride (CCl4)-Induced Hepatotoxicity in Rats. Molecules. 2022; 27(8):2593. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27082593
Chicago/Turabian StyleVun-Sang, Senty, Kenneth Francis Rodrigues, Urban J. A. Dsouza, and Mohammad Iqbal. 2022. "Suppression of Oxidative Stress and Proinflammatory Cytokines Is a Potential Therapeutic Action of Ficus lepicarpa B. (Moraceae) against Carbon Tetrachloride (CCl4)-Induced Hepatotoxicity in Rats" Molecules 27, no. 8: 2593. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27082593