

Why Do These Yeasts Smell So Good? Volatile Organic Compounds (VOCs) Produced by Malassezia Species in the Exponential and Stationary Growth Phases

 , and
, and
Abstract
:
1. Introduction
2. Results
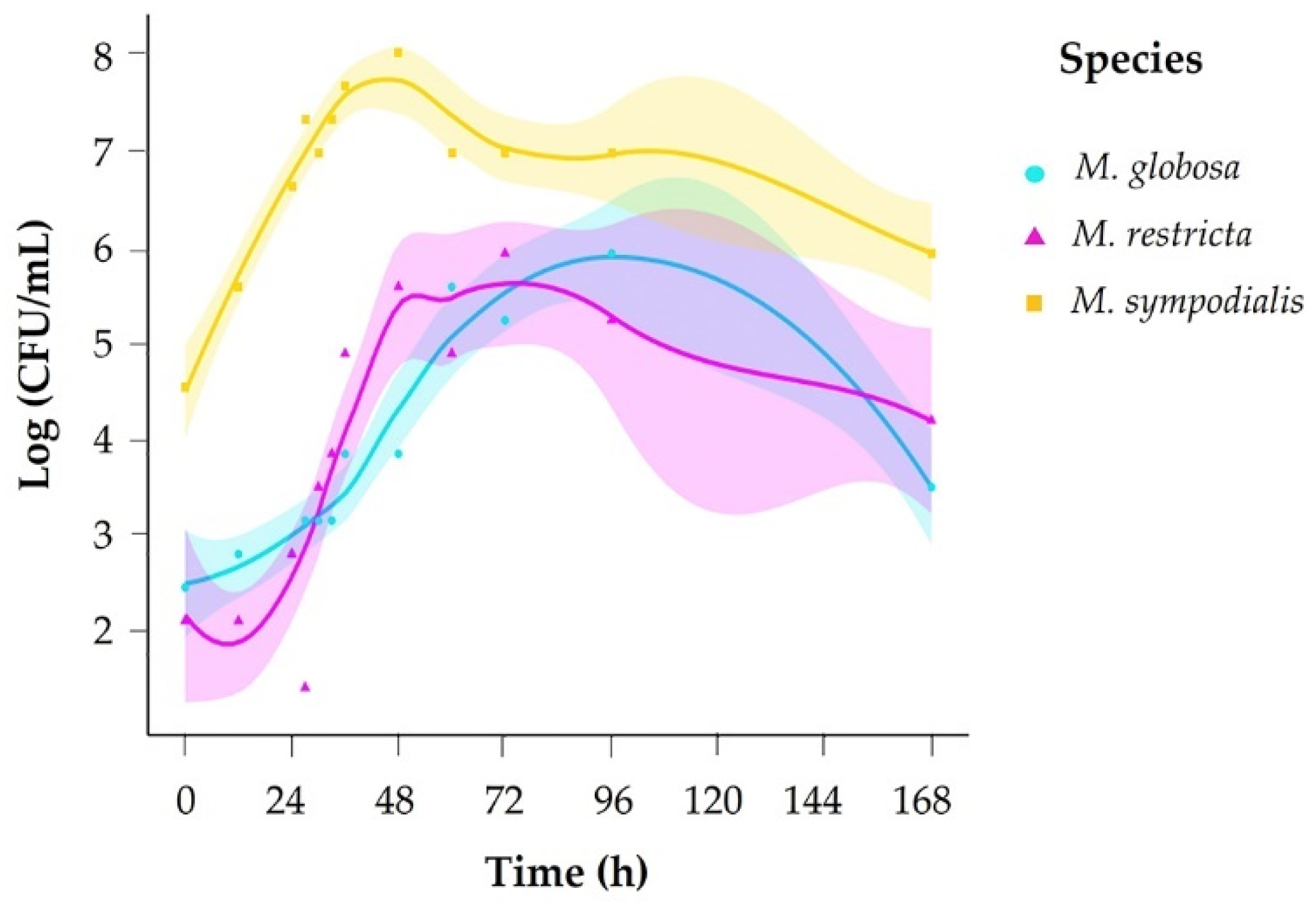
2.1. Growth Phases Differ between Malassezia Species
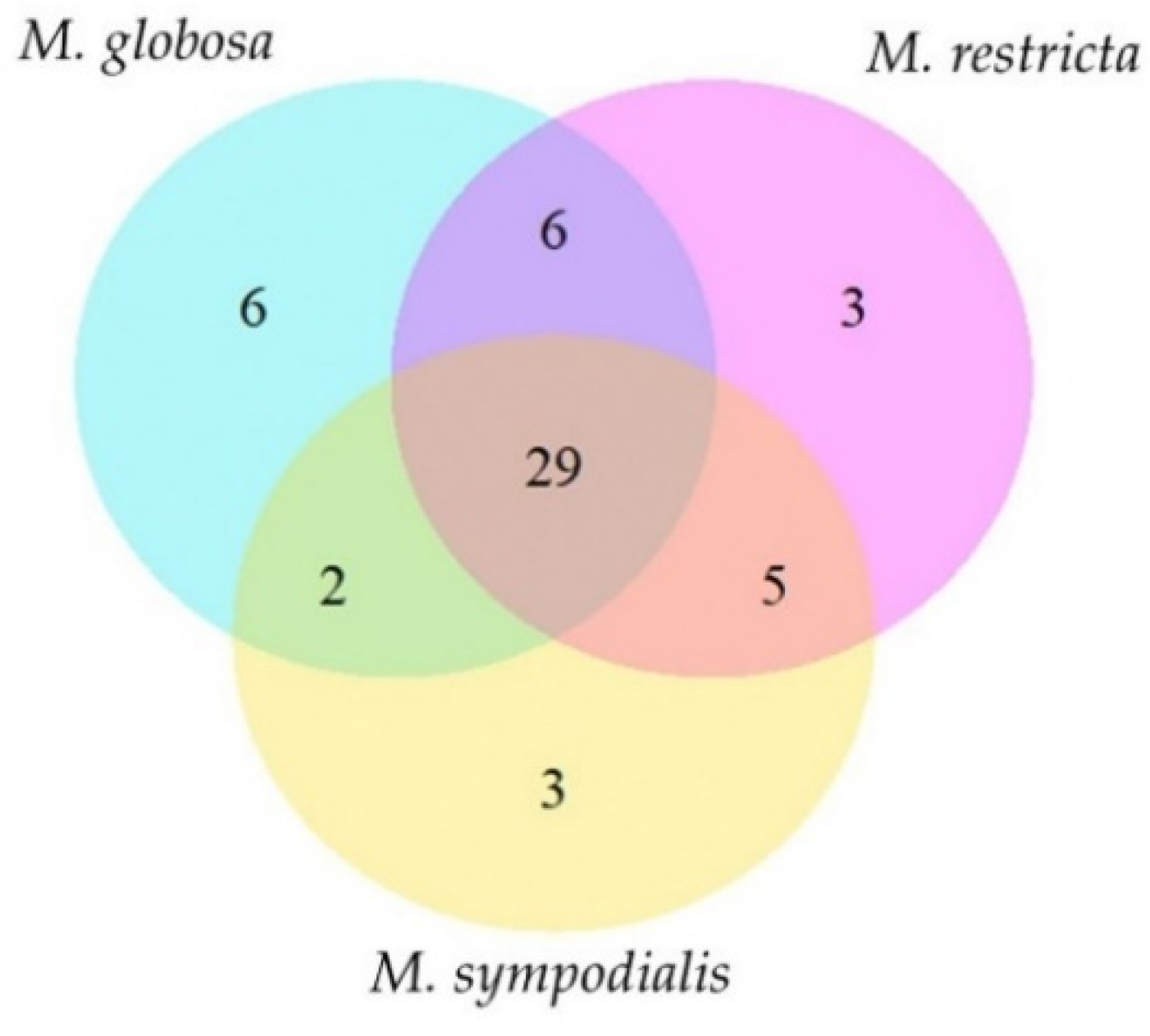
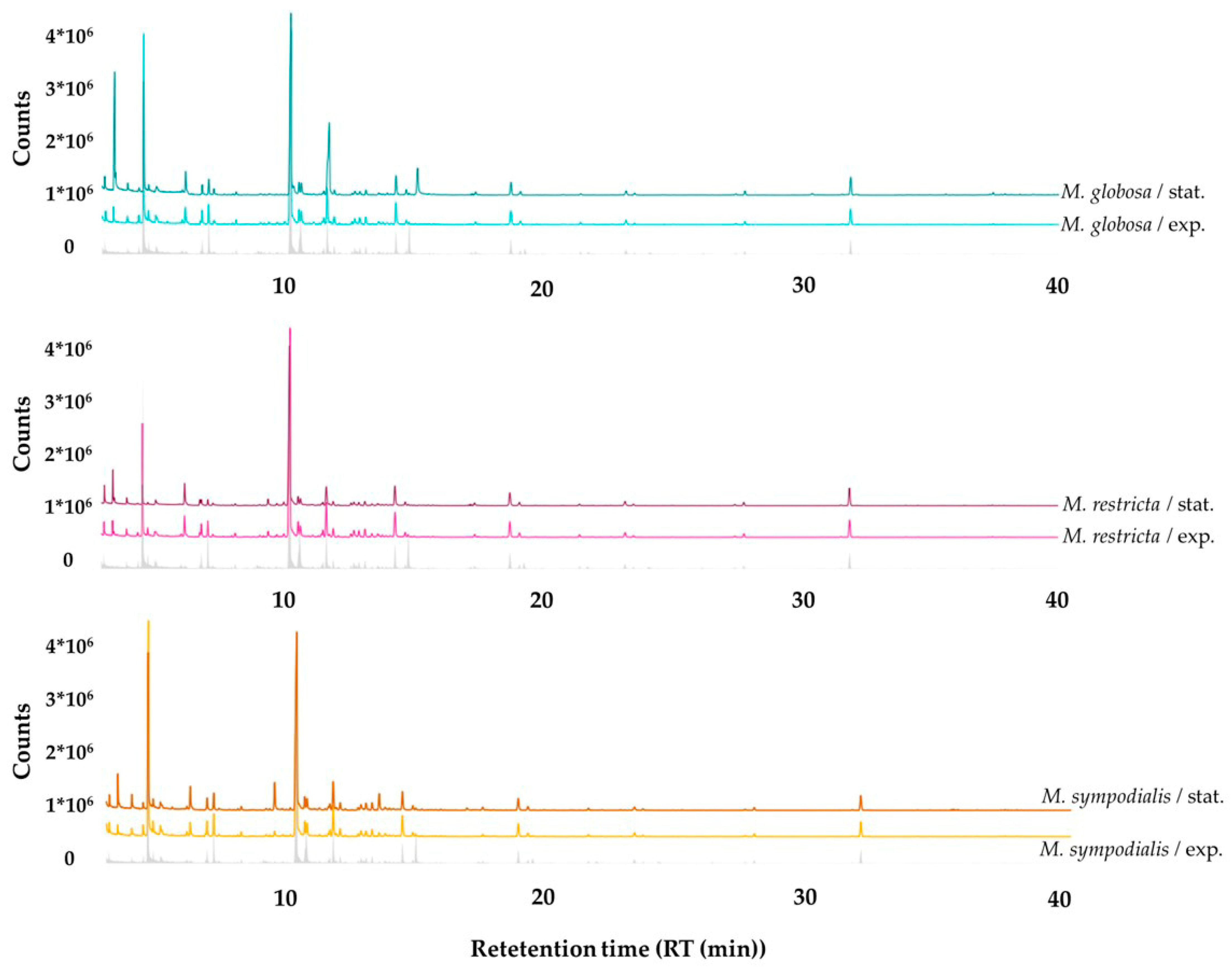
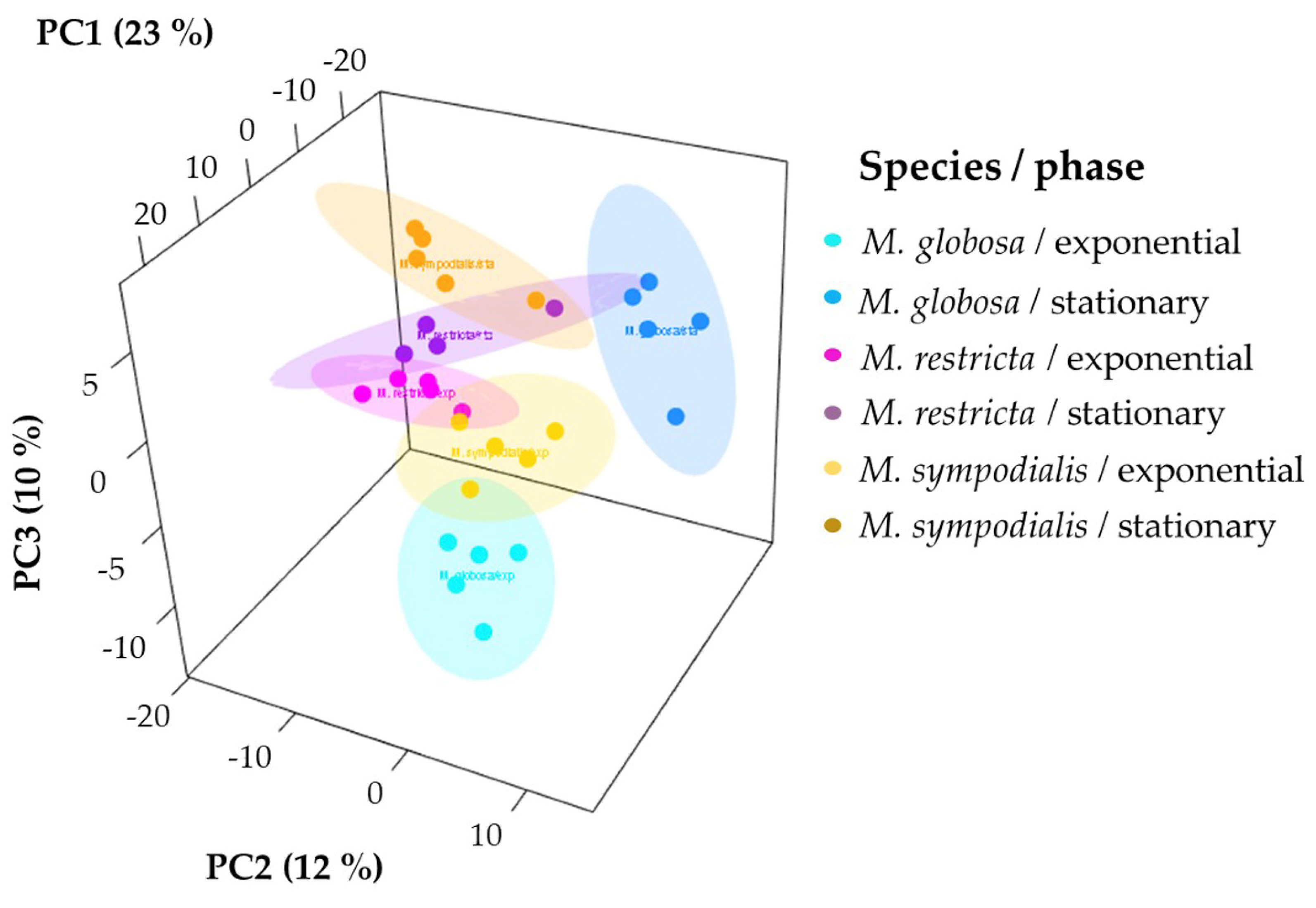
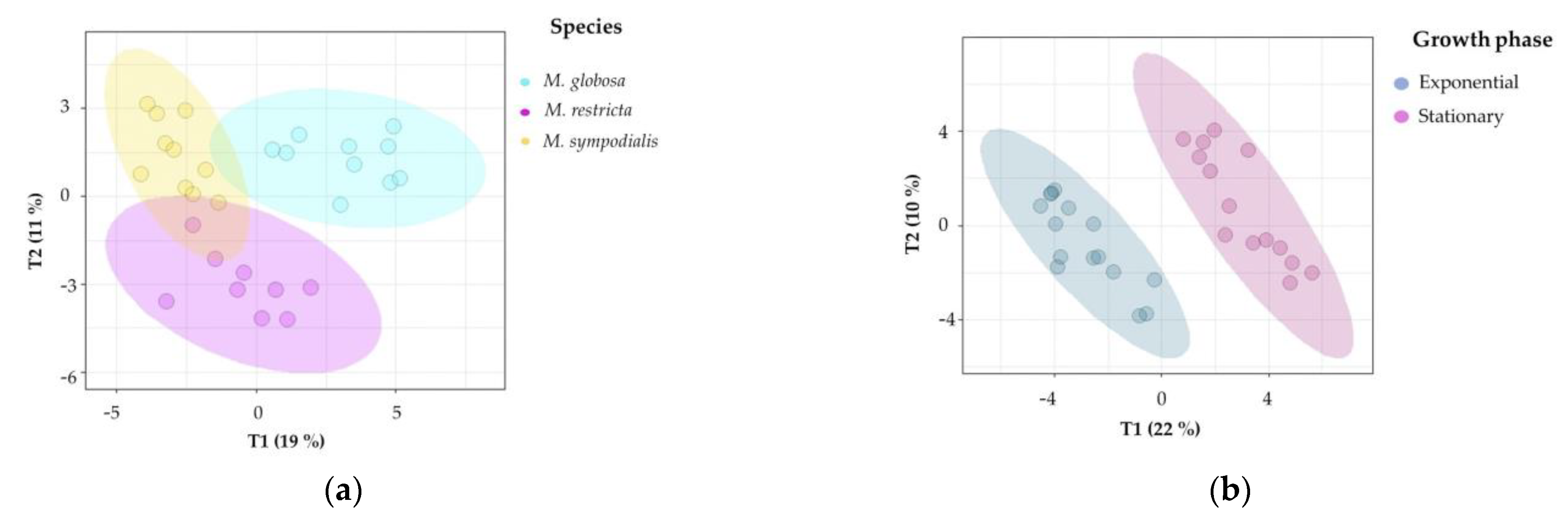
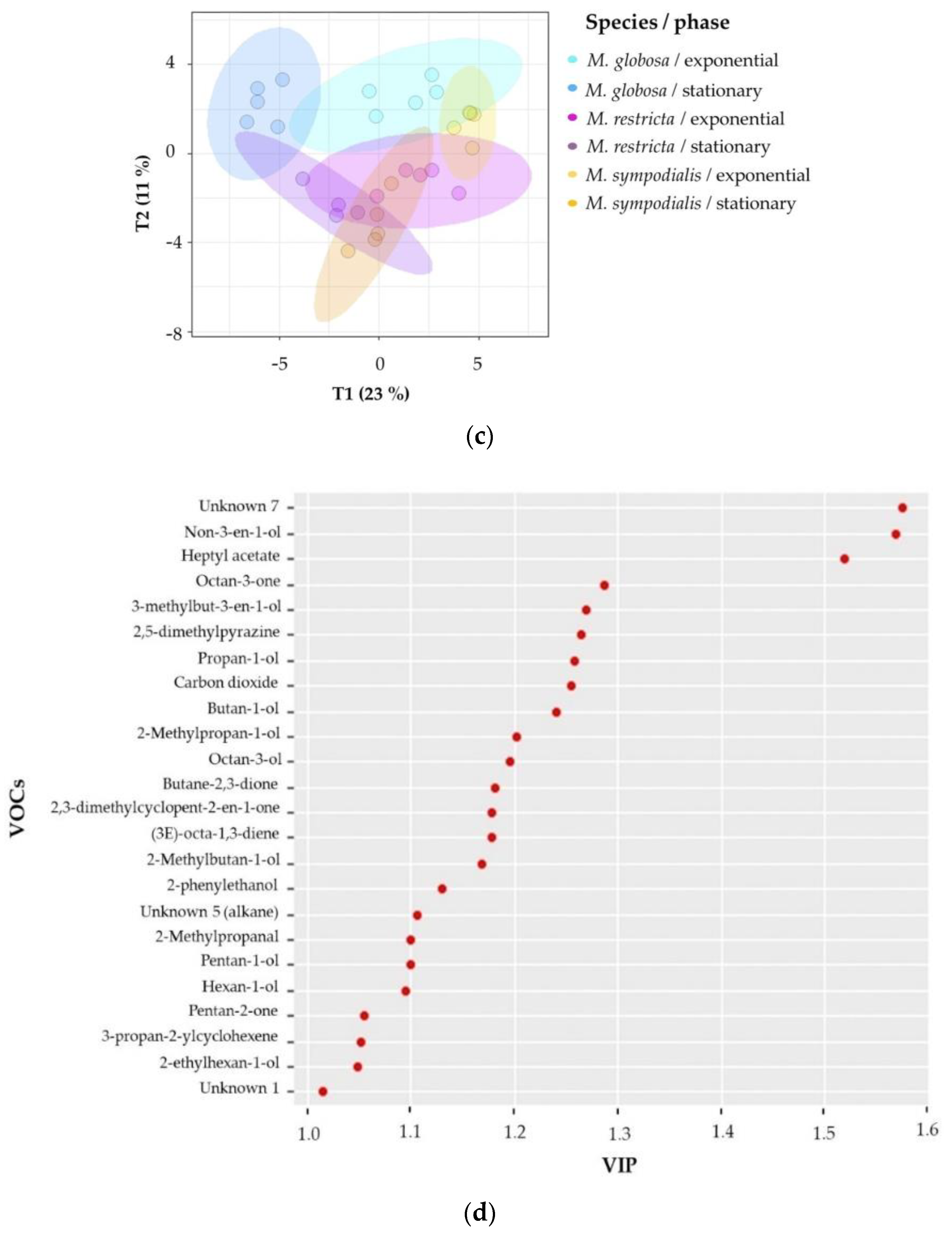
2.2. The Volatile Profile Allows the Separation of Malassezia Species and Growth Phases
3. Discussion
4. Materials and Methods
4.1. Strains and Growth Conditions
4.2. Growth Curve
4.3. VOC Production
4.4. VOC Sampling and GC–MS Analysis
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Chandra, S.H.V.; Srinivas, R.; Dawson, T.L.J.; Common, J.E. Cutaneous Malassezia: Commensal, Pathogen, or Protector? Front. Cell. Infect. Microbiol. 2021, 10, 614446. [Google Scholar] [CrossRef]
- Kashaf, S.S.; Proctor, D.M.; Deming, C.; Saary, P.; Hölzer, M.; Mullikin, J.; Thomas, J.; Young, A.; Bouffard, G.; Barnabas, B.; et al. Integrating cultivation and metagenomics for a multi-kingdom view of skin microbiome diversity and functions. Nat. Microbiol. 2021, 7, 169–179. [Google Scholar] [CrossRef]
- Celis, A.M.; Wösten, H.A.B.; Triana, S.; Restrepo, S.; de Cock, H. Malassezia spp. beyond the Mycobiota. SM Dermatol. J. 2017, 3, 1–10. [Google Scholar]
- Grice, E.A.; Dawson, T.L., Jr. Host–microbe interactions: Malassezia and human skin. Curr. Opin. Microbiol. 2017, 40, 81–87. [Google Scholar] [CrossRef]
- Theelen, B.; Cafarchia, C.; Gaitanis, G.; Bassukas, I.D.; Boekhout, T.; Dawson, T.L. Malassezia ecology, pathophysiology, and treatment. Med. Mycol. 2018, 56, 10–25. [Google Scholar] [CrossRef] [Green Version]
- Triana, S.; de Cock, H.; Ohm, R.A.; Danies, G.; Wösten, H.A.B.; Restrepo, S.; Barrios, A.F.G.; Celis, A. Lipid Metabolic Versatility in Malassezia spp. Yeasts Studied through Metabolic Modeling. Front. Microbiol. 2017, 8, 1772. [Google Scholar] [CrossRef]
- Nowicka, D.; Nawrot, U. Contribution of Malassezia spp. to the development of atopic dermatitis. Mycoses 2019, 62, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Limon, J.J.; Tang, J.; Li, D.; Wolf, A.J.; Michelsen, K.S.; Funari, V.; Gargus, M.; Nguyen, C.; Sharma, P.; Maymi, V.I.; et al. Malassezia Is Associated with Crohn’s Disease and Exacerbates Colitis in Mouse Models. Cell Host Microbe 2019, 25, 377–388.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurence, M.; Benito-León, J.; Calon, F. Malassezia and Parkinson’s Disease. Front. Neurol. 2019, 10, 758. [Google Scholar] [CrossRef] [Green Version]
- Ehemann, K.; Mantilla, M.J.; Mora-Restrepo, F.; Rios-Navarro, A.; Torres, M.; Ramírez, A.M.C. Many ways, one microorganism: Several approaches to study Malassezia in interactions with model hosts. PLoS Pathog. 2022, 18, e1010784. [Google Scholar] [CrossRef]
- Davis, T.S.; Crippen, T.L.; Hofstetter, R.W.; Tomberlin, J.K. Microbial Volatile Emissions as Insect Semiochemicals. J. Chem. Ecol. 2013, 39, 840–859. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.; Celis, A.M.; Guevara-Suarez, M.I.; Molina, J.; Carazzone, C. Yeast Smell Like What They Eat: Analysis of Volatile Organic Compounds of Malassezia furfur in Growth Media Supplemented with Different Lipids. Molecules 2019, 24, 419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, R.; Cordovez, V.; de Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz-Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial Volatiles: Small Molecules with an Important Role in Intra- and Inter-Kingdom Interactions. Front. Microbiol. 2017, 8, 2484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, J.W.; Inamdar, A. Are some fungal volatile organic compounds (VOCs) mycotoxins? Toxins 2015, 9, 3785–3804. [Google Scholar] [CrossRef] [Green Version]
- Veselova, M.A.; Plyuta, V.A.; Khmel, I.A. Volatile Compounds of Bacterial Origin: Structure, Biosynthesis, and Biological Activity. Microbiology 2019, 88, 261–274. [Google Scholar] [CrossRef]
- Bennett, J.W.; Hung, R.; Lee, S.; Padhi, S. 18 Fungal and Bacterial Volatile Organic Compounds: An Overview and Their Role as Ecological Signaling Agents. Fungal Assoc. 2012, 9, 373–393. [Google Scholar] [CrossRef]
- Inamdar, A.A.; Morath, S.; Bennett, J.W. Fungal Volatile Organic Compounds: More Than Just a Funky Smell? Annu. Rev. Microbiol. 2020, 74, 101–116. [Google Scholar] [CrossRef]
- Kai, M.; Elmassry, M.; Farag, M.A. Sampling, Detection, Identification, and Analysis of Bacterial Volatile Organic Compounds (VOCs). In Bacterial Volatile Compounds as Mediators of Airborne Interactions; Ryu, C.M., Weisskopf, L., Piechulla, B., Eds.; Springer: Singapore, 2020. [Google Scholar]
- Alpha, C.J.; Campos, M.; Jacobs-Wagner, C.; Strobel, S.A. Mycofumigation by the Volatile Organic Compound-Producing Fungus Muscodor albus Induces Bacterial Cell Death through DNA Damage. Appl. Environ. Microbiol. 2015, 81, 1147–1156. [Google Scholar] [CrossRef] [Green Version]
- Contarino, R.; Brighina, S.; Fallico, B.; Cirvilleri, G.; Parafati, L.; Restuccia, C. Volatile organic compounds (VOCs) produced by biocontrol yeasts. Food Microbiol. 2019, 82, 70–74. [Google Scholar] [CrossRef]
- Briard, B.; Heddergott, C.; Latgé, J.-P. Volatile Compounds Emitted by Pseudomonas aeruginosa Stimulate Growth of the Fungal Pathogen Aspergillus fumigatus. MBio 2016, 7, e00219-16. [Google Scholar] [CrossRef] [Green Version]
- Juarez, G.E.; Mateyca, C.; Galvan, E.M. Proteus mirabilis outcompetes Klebsiella pneumoniae in artificial urine medium through secretion of ammonia and other volatile compounds. Heliyon 2020, 6, e03361. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.M.; Lawal, O.; Nijsen, T.M.; Goodacre, R.; Fowler, S.J. Exhaled Volatile Organic Compounds of Infection: A Systematic Review. ACS Infect. Dis. 2017, 3, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Elmassry, M.M.; Piechulla, B. Volatilomes of Bacterial Infections in Humans. Front. Neurosci. 2020, 14, 257. [Google Scholar] [CrossRef] [PubMed]
- Gerritsen, M.G.; Brinkman, P.; Escobar, N.; Bos, L.D.; de Heer, K.; Meijer, M.; Janssen, H.-G.; de Cock, H.; Wösten, H.A.B.; Visser, C.E.; et al. Profiling of volatile organic compounds produced by clinical Aspergillus isolates using gas chromatography–mass spectrometry. Med. Mycol. 2017, 56, 253–256. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, D.K.; Sinclair, E.; Xu, Y.; Sarkar, D.; Walton-Doyle, C.; Liscio, C.; Banks, P.; Milne, J.; Silverdale, M.; Kunath, T.; et al. Discovery of Volatile Biomarkers of Parkinson’s Disease from Sebum. ACS Cent. Sci. 2019, 5, 599–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabares, M.; Ortiz, M.; González, M.; Carazzone, C.; Florez, M.J.V.; Molina, J. Behavioral responses of Rhodnius prolixus to volatile organic compounds released in vitro by bacteria isolated from human facial skin. PLoS Negl. Trop. Dis. 2018, 12, e0006423. [Google Scholar] [CrossRef] [Green Version]
- Verhulst, N.O.; Beijleveld, H.; Knols, B.G.; Takken, W.; Schraa, G.; Bouwmeester, H.J.; Smallegange, R.C. Cultured skin microbiota attracts malaria mosquitoes. Malar. J. 2009, 8, 302. [Google Scholar] [CrossRef] [Green Version]
- Almaliki, H.S.; Angela, A.; Goraya, N.J.; Yin, G.; Bennett, J.W. Volatile Organic Compounds Produced by Human Pathogenic Fungi Are Toxic to Drosophila melanogaster. Front. Fungal Biol. 2021, 1, 629510. [Google Scholar] [CrossRef]
- Lemfack, M.C.; Gohlke, B.-O.; Toguem, S.M.T.; Preissner, S.; Piechulla, B.; Preissner, R. mVOC 2.0: A database of microbial volatiles. Nucleic Acids Res. 2017, 46, D1261–D1265. [Google Scholar] [CrossRef] [Green Version]
- Rios-Navarro, A.; Gonzalez, M.; Carazzone, C.; Ramírez, A.M.C. Learning about microbial language: Possible interactions mediated by microbial volatile organic compounds (VOCs) and relevance to understanding Malassezia spp. metabolism. Metabolomics 2021, 17, 39. [Google Scholar] [CrossRef]
- Labows, J.N.; McGinley, K.J.; Leyden, J.J.; Webster, G.F. Characteristic gamma-lactone odor production of the genus Pityrosporum. Appl. Environ. Microbiol. 1979, 38, 412–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feunang, Y.D.; Eisner, R.; Knox, C.; Chepelev, L.; Hastings, J.; Owen, G.; Fahy, E.; Steinbeck, C.; Subramanian, S.; Bolton, E.; et al. ClassyFire: Automated chemical classification with a comprehensive, computable taxonomy. J. Cheminform. 2016, 8, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akarachantachote, N.; Chadcham, S.; Saithanu, K. Cutoff threshold of variable importance in projection for variable selection. Int. J. Pure Appl. Math. 2014, 94, 307–322. [Google Scholar] [CrossRef]
- Schaechter, M. From growth physiology to systems biology. Int. Microbiol. 2006, 9, 157–161. [Google Scholar] [PubMed]
- Tonner, P.D.; Darnell, C.L.; Bushell, F.M.L.; Lund, P.A.; Schmid, A.K.; Schmidler, S.C. A Bayesian non-parametric mixed-effects model of microbial growth curves. PLoS Comput. Biol. 2020, 16, e1008366. [Google Scholar] [CrossRef] [PubMed]
- Combet, E.; Eastwood, D.C.; Burton, K.S.; Henderson, J. Eight-carbon volatiles in mushrooms and fungi: Properties, analysis, and biosynthesis. Mycoscience 2006, 47, 317–326. [Google Scholar] [CrossRef]
- Drilling, K.; Dettner, K. Electrophysiological responses of four fungivorous coleoptera to volatiles of Trametes versicolor: Implications for host selection. Chemoecology 2009, 19, 109–115. [Google Scholar] [CrossRef]
- Farh, M.E.-A.; Jeon, J. Roles of Fungal Volatiles from Perspective of Distinct Lifestyles in Filamentous Fungi. Plant Pathol. J. 2020, 36, 193–203. [Google Scholar] [CrossRef]
- Ramírez, A.M.C.; Amézquita, A.; Jaramillo, J.E.C.C.; Matiz-Cerón, L.F.; Andrade-Martínez, J.S.; Triana, S.; Mantilla, M.J.; Restrepo, S.; Barrios, A.F.G.; De Cock, H. Analysis of Malassezia Lipidome Disclosed Differences Among the Species and Reveals Presence of Unusual Yeast Lipids. Front. Cell. Infect. Microbiol. 2020, 10, 338. [Google Scholar] [CrossRef]
- De Vrieze, M.; Pandey, P.; Bucheli, T.D.; Varadarajan, A.R.; Ahrens, C.H.; Weisskopf, L.; Bailly, A. Volatile Organic Compounds from Native Potato-associated Pseudomonas as Potential Anti-oomycete Agents. Front. Microbiol. 2015, 6, 1295. [Google Scholar] [CrossRef] [Green Version]
- Elmassry, M.M.; Farag, M.A.; Preissner, R.; Gohlke, B.-O.; Piechulla, B.; Lemfack, M.C. Sixty-One Volatiles Have Phylogenetic Signals Across Bacterial Domain and Fungal Kingdom. Front. Microbiol. 2020, 11, 557253. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, D.; Gershenzon, J.; Hammerbacher, A. Volatile Organic Compounds Emitted by Fungal Associates of Conifer Bark Beetles and their Potential in Bark Beetle Control. J. Chem. Ecol. 2016, 42, 952–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, C.; Bean, H. Dependence of the Staphylococcal Volatilome Composition on Microbial Nutrition. Metabolites 2020, 10, 347. [Google Scholar] [CrossRef] [PubMed]
- Hung, R.; Lee, S.; Bennett, J.W. Fungal volatile organic compounds and their role in ecosystems. Appl. Microbiol. Biotechnol. 2015, 99, 3395–3405. [Google Scholar] [CrossRef] [PubMed]
- Sawahata, T.; Shimano, S.; Suzuki, M. Tricholoma matsutake 1-Octen-3-ol and methyl cinnamate repel mycophagous Proisotoma minuta (Collembola: Insecta). Mycorrhiza 2008, 18, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Al-Fatimi, M.; Wurster, M.; Lindequist, U. Chemical Composition, Antimicrobial and Antioxidant Activities of the Volatile Oil of Ganoderma pfeifferi Bres. Medicines 2016, 3, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dandurishvili, N.; Toklikishvili, N.; Ovadis, M.; Eliashvili, P.; Giorgobiani, N.; Keshelava, R.; Tediashvili, M.; Vainstein, A.; Khmel, I.; Szegedi, E.; et al. Broad-range antagonistic rhizobacteria Pseudomonas fluorescens and Serratia plymuthica suppress Agrobacterium crown gall tumours on tomato plants. J. Appl. Microbiol. 2010, 110, 341–352. [Google Scholar] [CrossRef]
- Scott, J.; Sueiro-Olivares, M.; Ahmed, W.; Heddergott, C.; Zhao, C.; Thomas, R.; Bromley, M.; Latgé, J.-P.; Krappmann, S.; Fowler, S.; et al. Pseudomonas aeruginosa-Derived Volatile Sulfur Compounds Promote Distal Aspergillus fumigatus Growth and a Synergistic Pathogen-Pathogen Interaction that Increases Pathogenicity in Co-infection. Front. Microbiol. 2019, 10, 2311. [Google Scholar] [CrossRef]
- Peñuelas, J.; Asensio, D.; Tholl, D.; Wenke, K.; Rosenkranz, M.; Piechulla, B.; Schnitzler, J. Biogenic volatile emissions from the soil. Plant Cell Environ. 2014, 37, 1866–1891. [Google Scholar] [CrossRef]
- Lee, S.; Yap, M.; Behringer, G.; Hung, R.; Bennett, J.W. Volatile organic compounds emitted by Trichoderma species mediate plant growth. Fungal Biol. Biotechnol. 2016, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Schulz, S.; Dickschat, J.S. Bacterial volatiles: The smell of small organisms. Nat. Prod. Rep. 2007, 24, 814–842. [Google Scholar] [CrossRef] [PubMed]
- Dolenc-Voljč, M. Diseases Caused by Malassezia Species in Human Beings. Microbiol. Ski. Soft Tissue Bone Jt. Infect. 2017, 2, 77–91. [Google Scholar] [CrossRef]
- Ashrafi, M.; Novak-Frazer, L.; Bates, M.; Baguneid, M.; Alonso-Rasgado, T.; Xia, G.; Rautemaa-Richardson, R.; Bayat, A. Validation of biofilm formation on human skin wound models and demonstration of clinically translatable bacteria-specific volatile signatures. Sci. Rep. 2018, 8, 9431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Encinas-Basurto, D.; Valenzuela-Quintanar, M.I.; Sánchez-Estrada, A.; Tiznado-Hernández, M.E.; Rodríguez-Félix, A.; Troncoso-Rojas, R. Alterations in volatile metabolites profile of fresh tomatoes in response to Alternaria alternata (Fr.) Keissl. 1912 infection. Chil. J. Agric. Res. 2017, 77, 194–201. [Google Scholar] [CrossRef] [Green Version]
- Cale, J.A.; Collignon, R.M.; Klutsch, J.G.; Kanekar, S.S.; Hussain, A.; Erbilgin, N. Fungal Volatiles Can Act as Carbon Sources and Semiochemicals to Mediate Interspecific Interactions Among Bark Beetle-Associated Fungal Symbionts. PLoS ONE 2016, 11, e0162197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korpi, A.; Järnberg, J.; Pasanen, A.L. Microbial volatile organic compounds. Crit. Rev. Toxicol. 2009, 39, 139–193. [Google Scholar] [CrossRef]
- Junior, W.J.L.; Binati, R.L.; Felis, G.E.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Volatile organic compounds from Starmerella bacillaris to control gray mold on apples and modulate cider aroma profile. Food Microbiol. 2020, 89, 103446. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Ruiz, J.; Esteban-Fernández, A.; Navascués, E.; Marquina, D.; Santos, A.; Moreno-Arribas, M.V. Microbial contribution to wine aroma and its intended use for wine quality improvement. Molecules 2017, 22, 189. [Google Scholar] [CrossRef] [Green Version]
- Boekhout, T.; Guého-Kellermann, E.; Mayser, P.; Velegraki, A. (Eds.) Malassezia and the Skin: Science and Clinical Practice; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Reese, K.L.; Rasley, A.; Avila, J.R.; Jones, A.D.; Frank, M. Metabolic Profiling of Volatile Organic Compounds (VOCs) Emitted by the Pathogens Francisella tularensis and Bacillus anthracis in Liquid Culture. Sci. Rep. 2020, 10, 9333. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- van den Berg, R.A.; Hoefsloot, H.C.J.; Westerhuis, J.A.; Smilde, A.K.; Van Der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodacre, R.; Broadhurst, D.; Smilde, A.K.; Kristal, B.S.; Baker, J.D.; Beger, R.; Bessant, C.; Connor, S.; Capuani, G.; Craig, A.; et al. Proposed minimum reporting standards for data analysis in metabolomics. Metabolomics 2007, 3, 231–241. [Google Scholar] [CrossRef]
- Paliy, O.; Shankar, V. Application of multivariate statistical techniques in microbial ecology. Mol. Ecol. 2016, 25, 1032–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, Y.; Hu, Y.-J. Extension of PERMANOVA to Testing the Mediation Effect of the Microbiome. Genes 2022, 13, 940. [Google Scholar] [CrossRef] [PubMed]
- Gromski, P.S.; Muhamadali, H.; Ellis, D.I.; Xu, Y.; Correa, E.; Turner, M.L.; Goodacre, R. A tutorial review: Metabolomics and partial least squares-discriminant analysis—A marriage of convenience or a shotgun wedding. Anal. Chim. Acta 2015, 879, 10–23. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
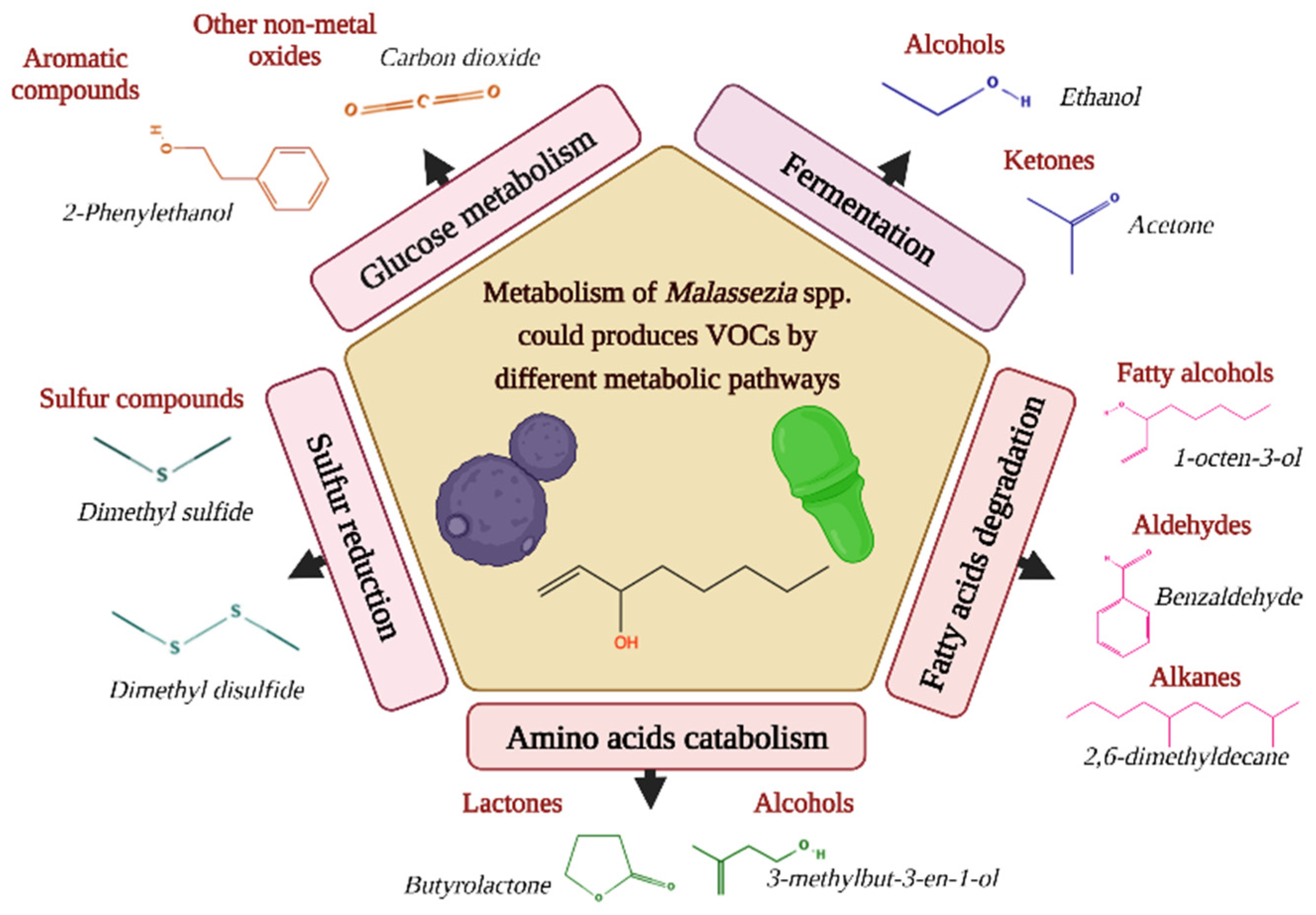{kind=link}
| M. globosa | M. restricta | M. sympodialis | |
|---|---|---|---|
| Exponential phase | 30 h | 30 h | 10 h |
| Stationary phase | 90 h | 55 h | 35 h |
| No. | Compound | CAS | RI | RI Exp. | Ret. Time | ClassyFire Class/Subclass | M. globosa | M. restricta | M. sympodialis | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Growth Phase | ||||||||||||||||||||||||
| Exp. | Stat. | Exp. | Stat. | Exp. | Stat. | |||||||||||||||||||
| % Area | SD | N | % Area | SD | N | % Area | SD | N | % Area | SD | N | % Area | SD | N | % Area | SD | N | |||||||
| 1 | Carbon dioxide | 124-38-9 | 1.33 | Other nonmetal oxides | 13.89 | 5.7 | 5 | 11.17 | 4.2 | 5 | 11.28 | 1.3 | 5 | 22.78 | 5.6 | 4 | 7.06 | 2.1 | 5 | 23.67 | 2 | 5 | ||
| 2 | Unknown 1 | 1.80 | 2.51 | 2.6 | 3 | 0.77 | 1 | 1.57 | 1 | 4.52 | 4.3 | 3 | ||||||||||||
| 3 | Propan-1-ol | 71-23-8 | 568 | 1.82 | Alcohols and polyols | 3.81 | 2.8 | 4 | 3.09 | 3.1 | 3 | |||||||||||||
| 4 | 2-Methylpropanal | 78-84-2 | 532 | 1.82 | Carbonyl compounds | 1.29 | 1 | 2.84 | 3.5 | 3 | ||||||||||||||
| 5 | Butane-2,3-dione | 431-03-8 | 591 | 1.96 | Carbonyl compounds | 4.70 | 2.9 | 4 | 2.57 | 2.5 | 3 | |||||||||||||
| 6 | Hex-1-ene | 592-41-6 | 601 | 1.99 | Unsaturated aliphatic hydrocarbons | 0.88 | 1 | 1.96 | 1 | 5 | 0.92 | 1 | 0.53 | 1 | 1.60 | 2.2 | 2 | 0.50 | 1 | |||||
| 7 | 2-Methylpropan-1-ol | 78-83-1 | 614 | 626 | 2.26 | Alcohols and polyols | 4.35 | 3.7 | 4 | 8.73 | 5.5 | 4 | 7.80 | 3.9 | 5 | |||||||||
| 8 | Butan-1-ol | 71-36-3 | 653 | 657 | 2.56 | Alcohols and polyols | 4.12 | 4.8 | 3 | 3.73 | 0.9 | 5 | 9.26 | 3.7 | 5 | 8.01 | 1.3 | 4 | 7.37 | 2.6 | 5 | |||
| 9 | Pentan-2-one | 107-87-9 | 695 | 684 | 2.80 | Organooxygen compounds | 3.24 | 2.7 | 5 | 1.45 | 0.6 | 5 | 3.12 | 2.5 | 5 | 5.45 | 1 | 4 | 1.09 | 1 | 3 | 1.89 | 0.4 | 5 |
| 10 | Methylcyclohexane | 108-87-2 | 755 | 720 | 3.28 | Cycloalkanes | 1.33 | 1.9 | 2 | 0.66 | 1 | 2 | 0.75 | 1 | 2.47 | 2.3 | 3 | 0.59 | 1 | |||||
| 11 | 2,5-Dimethylhexane | 592-13-2 | 726 | 3.38 | Alkanes | 0.29 | 1 | 0.12 | 1 | 0.46 | 1 | |||||||||||||
| 12 | 3-Methylbut-3-en-1-ol | 763-32-6 | 731 | 727 | 3.40 | Alcohols and polyols | 0.23 | 1 | 1.03 | 0.8 | 3 | |||||||||||||
| 13 | 3-Methylbutan-1-ol | 123-51-3 | 735 | 730 | 3.45 | Alcohols and polyols | 14.32 | 4.4 | 5 | 21.80 | 6.3 | 5 | 9.52 | 1.9 | 5 | 21.07 | 9.2 | 4 | 9.25 | 2.6 | 5 | 11.74 | 4.1 | 5 |
| 14 | 2-Methylbutan-1-ol | 137-32-6 | 767 | 734 | 3.50 | Alcohols and polyols | 3.84 | 3.5 | 3 | 4.69 | 0.7 | 5 | 6.38 | 1.9 | 4 | 3.52 | 2 | 4 | 3.00 | 2.2 | 4 | |||
| 15 | 3-Methylpentan-2-one | 565-61-7 | 759 | 748 | 3.74 | Carbonyl compounds | 2.74 | 0.7 | 5 | 0.98 | 1 | |||||||||||||
| 16 | Pentan-1-ol | 71-41-0 | 751 | 763 | 3.98 | Alcohols and polyols | 3.53 | 2.2 | 4 | 3.05 | 4.4 | 2 | 1.05 | 1 | 7.77 | 4.4 | 4 | |||||||
| 17 | Unknown 2 | 768 | 4.06 | 0.46 | 1 | 0.54 | 1 | |||||||||||||||||
| 18 | (E)-oct-3-ene | 14919-01-8 | 797 | 4.54 | Branched unsaturated hydrocarbons | 0.31 | 0.4 | 2 | 0.11 | 1 | ||||||||||||||
| 19 | (Z)-oct-3-ene | 14850-22-7 | 805 | 4.72 | Unsaturated aliphatic hydrocarbons | 3.47 | 2.4 | 4 | 0.88 | 0.8 | 4 | 2.48 | 1.5 | 4 | 0.26 | 0.4 | 2 | 5.17 | 2.9 | 4 | 1.95 | 1.3 | 4 | |
| 20 | Unknown 3 | 813 | 4.90 | 3.93 | 2.7 | 4 | 1.32 | 1.4 | 3 | 3.64 | 1.6 | 5 | 0.83 | 0.8 | 3 | 8.28 | 1.1 | 5 | 2.34 | 1.9 | 4 | |||
| 21 | 2,4-Dimethylheptane | 2213-23-2 | 818 | 819 | 5.08 | Alkanes | 15.00 | 8.7 | 4 | 1.44 | 1 | 4.07 | 5.6 | 2 | 12.42 | 3.9 | 5 | 2.62 | 2.4 | 3 | ||||
| 22 | (3E)-octa-1,3-diene | 1002-33-1 | 827 | 822 | 5.14 | Olefins | 6.78 | 3.8 | 4 | 2.16 | 2.1 | 3 | ||||||||||||
| 23 | Propylcyclopentane | 2040-96-2 | 828 | 5.30 | Cycloalkanes | 1.28 | 1.1 | 3 | 1.02 | 1.2 | 3 | 2.05 | 1.5 | 5 | 0.47 | 0.4 | 3 | 4.38 | 1.8 | 5 | 1.49 | 1.6 | 4 | |
| 24 | 2,4-Dimethylhept-1-ene | 19549-87-2 | 842 | 837 | 5.53 | Branched unsaturated hydrocarbons | 3.63 | 3.3 | 5 | 0.45 | 0.4 | 3 | 0.67 | 0.6 | 5 | 0.12 | 1 | 1.73 | 1.1 | 5 | 0.60 | 0.5 | 4 | |
| 25 | 3-Methylhexan-2-one | 2550-21-2 | 841 | 5.62 | Carbonyl compounds | 1.14 | 0.4 | 5 | 0.39 | 1 | ||||||||||||||
| 26 | Ethylbenzene | 100-41-4 | 860 | 857 | 6.02 | Benzene and substituted derivatives | 0.23 | 1 | 0.39 | 0.4 | 3 | 0.52 | 0.8 | 2 | 0.57 | 0.4 | 3 | 1.43 | 0.6 | 5 | 0.62 | 0.4 | 5 | |
| 27 | Hexan-1-ol | 111-27-3 | 861 | 865 | 6.22 | Fatty alcohols | 19.96 | 5.9 | 5 | 4.96 | 1 | 5 | 22.16 | 6.2 | 5 | 13.52 | 2.3 | 4 | 16.16 | 1.5 | 5 | 11.51 | 1.5 | 5 |
| 28 | 3-Methylbutyl acetate | 123-92-2 | 884 | 873 | 6.44 | Carboxylic acid derivatives | 0.25 | 0.3 | 2 | 0.20 | 0 | 5 | 0.18 | 1 | 0.43 | 0.5 | 3 | |||||||
| 29 | 3-Methylhexan-2-ol | 2313-65-7 | 877 | 6.52 | Alcohols and polyols | 0.15 | 0.1 | 4 | 0.20 | 1 | 0.15 | 1 | ||||||||||||
| 30 | 2,5-Dimethylpyrazine | 123-32-0 | 911 | 909 | 7.42 | Pyrazines | 0.07 | 1 | 1.26 | 0.9 | 4 | 0.38 | 0.5 | 2 | 0.15 | 0.2 | 2 | |||||||
| 31 | 3-Propan-2-ylcyclohexene | 3983-08-2 | 944 | 8.66 | Unsaturated aliphatic hydrocarbons | 0.38 | 0.3 | 3 | 0.04 | 1 | ||||||||||||||
| 32 | 2,3-Dimethylcyclopent-2-en-1-one | 1121-05-7 | 947 | 8.75 | Olefins | 0.18 | 0.2 | 3 | 0.03 | 1 | ||||||||||||||
| 33 | Unknown 4 (alkane) | 958 | 9.13 | Alkanes | 2.71 | 0.6 | 5 | 0.36 | 0.1 | 5 | 2.59 | 0.8 | 5 | 1.16 | 0.5 | 4 | 2.73 | 0.3 | 5 | 1.33 | 0.3 | 5 | ||
| 34 | Hept-2-en-1-ol | 33467-76-4 | 965 | 9.40 | Fatty alcohols | 0.04 | 0.1 | 2 | ||||||||||||||||
| 35 | Octan-3-one | 106-68-3 | 987 | 984 | 10.06 | Carbonyl compounds | 3.36 | 2.3 | 4 | 0.03 | 1 | |||||||||||||
| 36 | Octan-3-ol | 589-98-0 | 994 | 994 | 10.40 | Fatty alcohols | 2.99 | 0.9 | 5 | |||||||||||||||
| 37 | 2,6-Dimethylnonane | 17302-28-2 | 1030 | 987 | 10.97 | Alkanes | 0.50 | 1 | 0.05 | 1 | 0.75 | 0.8 | 3 | 0.22 | 1 | 1.07 | 0.7 | 4 | 0.47 | 0.5 | 3 | |||
| 38 | Unknown 5 (alkane) | 996 | 11.03 | Alkanes | 0.46 | 0.6 | 2 | 0.26 | 0.1 | 5 | 1.77 | 0.6 | 5 | 0.82 | 0.3 | 4 | 1.94 | 0.4 | 5 | 0.95 | 0.2 | 5 | ||
| 39 | 2-Ethylhexan-1-ol | 104-76-7 | 1039 | 1028 | 11.78 | Fatty alcohols | 18.18 | 11 | 4 | |||||||||||||||
| 40 | Unknown 6 (alcohol) | 1081 | 13.96 | 0.48 | 0.7 | 2 | 1.12 | 0.2 | 5 | 0.37 | 0.6 | 2 | ||||||||||||
| 41 | 2,6-Dimethyldecane | 13150-81-7 | 1119 | 1103 | 14.87 | Alkanes | 2.46 | 2.4 | 3 | 0.47 | 0.2 | 5 | 3.08 | 1.0 | 5 | 1.36 | 0.4 | 4 | 3.25 | 0.6 | 5 | 1.29 | 0.3 | 5 |
| 42 | 2-Phenylethanol | 60-12-8 | 1115 | 1111 | 15.21 | Benzene and substituted derivatives | 4.93 | 1.3 | 5 | 0.14 | 1 | 0.10 | 1 | |||||||||||
| 43 | Heptyl acetate | 112-06-1 | 1091 | 1112 | 15.29 | Carboxylic acid derivatives | 0.68 | 0.3 | 5 | 0.62 | 0.3 | 4 | 0.19 | 0.3 | 2 | |||||||||
| 44 | Non-3-en-1-ol | 51494-28-1 | 1126 | 1147 | 16.84 | Fatty alcohols | 0.05 | 0.1 | 3 | 0.03 | 1 | 1.53 | 0.5 | 5 | ||||||||||
| 45 | Unknown 7 | 1153 | 17.08 | 0.39 | 0.2 | 5 | ||||||||||||||||||
| 46 | Unknown 8 | 1157 | 17.26 | 0.43 | 0.3 | 4 | 0.10 | 0 | 5 | 0.68 | 0.3 | 5 | 0.18 | 0.2 | 2 | 0.58 | 0.2 | 5 | 0.20 | 0.2 | 3 | |||
| 47 | Isomer of dimethyldecane | 1164 | 17.57 | Saturated hydrocarbons | 0.36 | 0.2 | 5 | 0.07 | 0.1 | 5 | 0.46 | 0.4 | 4 | 0.19 | 0.2 | 3 | 0.70 | 0.3 | 5 | 0.36 | 0.1 | 5 | ||
| 48 | Unknown 9 | 1165 | 17.64 | 0.21 | 0.3 | 2 | 0.09 | 0.1 | 2 | 0.10 | 1 | 0.07 | 0.1 | 2 | ||||||||||
| 49 | Unknown 10 (alcohol) | 1176 | 18.12 | 0.06 | 1 | 0.04 | 0.1 | 2 | ||||||||||||||||
| 50 | Unknown 11 | 1181 | 18.35 | 0.11 | 0.1 | 4 | 0.10 | 0.1 | 2 | 0.17 | 0.1 | 3 | 0.19 | 0.2 | 3 | 0.20 | 0 | 5 | ||||||
| 51 | 2,8-Dimethylundecane | 17301-25-6 | 1225 | 20.25 | Alkanes | 0.29 | 0.3 | 3 | 0.05 | 0.1 | 3 | 0.25 | 0.3 | 3 | 0.10 | 0.1 | 2 | 0.37 | 0.1 | 5 | 0.15 | 0.1 | 4 | |
| 52 | Unknown 12 (alcohol) | 1350 | 25.74 | 0.04 | 1 | 0.04 | 1 | |||||||||||||||||
| 53 | Unknown 13 (alkane) | 1371 | 26.60 | 0.03 | 1 | 0.05 | 1 | 0.02 | 1 | |||||||||||||||
| 54 | Unknown 14 | 1471 | 30.75 | 0.01 | 1 | 0.05 | 1 | |||||||||||||||||
| Variable | R2 | p Value |
|---|---|---|
| Species | 0.20 | <0.001 |
| Phase | 0.20 | <0.001 |
| Species/phase | 0.56 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rios-Navarro, A.; Gonzalez, M.; Carazzone, C.; Celis Ramírez, A.M. Why Do These Yeasts Smell So Good? Volatile Organic Compounds (VOCs) Produced by Malassezia Species in the Exponential and Stationary Growth Phases. Molecules 2023, 28, 2620. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28062620
Rios-Navarro A, Gonzalez M, Carazzone C, Celis Ramírez AM. Why Do These Yeasts Smell So Good? Volatile Organic Compounds (VOCs) Produced by Malassezia Species in the Exponential and Stationary Growth Phases. Molecules. 2023; 28(6):2620. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28062620
Chicago/Turabian StyleRios-Navarro, Andrea, Mabel Gonzalez, Chiara Carazzone, and Adriana Marcela Celis Ramírez. 2023. "Why Do These Yeasts Smell So Good? Volatile Organic Compounds (VOCs) Produced by Malassezia Species in the Exponential and Stationary Growth Phases" Molecules 28, no. 6: 2620. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28062620