
Microbial Production of Human Milk Oligosaccharides

Department of Chemistry, University of California, Davis, Davis, CA 95616, USA
*
Author to whom correspondence should be addressed.
Molecules 2023, 28(3), 1491; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28031491
Submission received: 30 December 2022
/
Revised: 25 January 2023
/
Accepted: 31 January 2023
/
Published: 3 February 2023
(This article belongs to the Special Issue Feature Papers in Chemical Biology—Edition of 2022-2023)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Human milk oligosaccharides (HMOs) are complex nonnutritive sugars present in human milk. These sugars possess prebiotic, immunomodulatory, and antagonistic properties towards pathogens and therefore are important for the health and well-being of newborn babies. Lower prevalence of breastfeeding around the globe, rising popularity of nutraceuticals, and low availability of HMOs have inspired efforts to develop economically feasible and efficient industrial-scale production platforms for HMOs. Recent progress in synthetic biology and metabolic engineering tools has enabled microbial systems to be a production system of HMOs. In this regard, the model organism Escherichia coli has emerged as the preferred production platform. Herein, we summarize the remarkable progress in the microbial production of HMOs and discuss the challenges and future opportunities in unraveling the scope of production of complex HMOs. We focus on the microbial production of five HMOs that have been approved for their commercialization.
1. Introduction
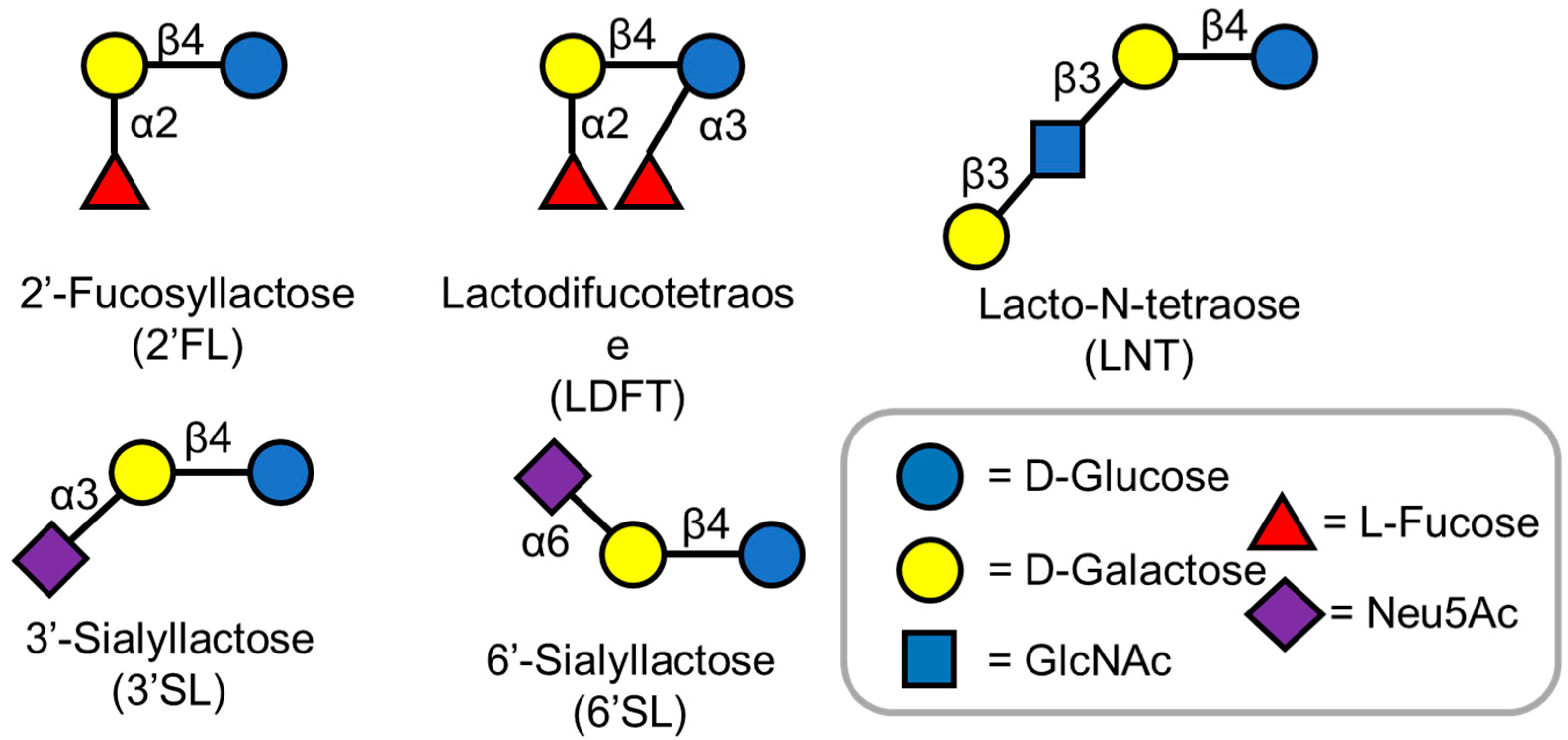
Carbohydrates are the most abundant biomolecules on earth and also are the primary energy source for diverse organisms [1]. Human milk, the primary source of nutrition for infants, contains about 7% carbohydrates [2]. Lactose is the primary sugar component in human milk. In addition to lactose, various non-lactose carbohydrates, called human milk oligosaccharides (HMOs) have been identified [3]. Currently, over 200 different HMO structures are known, and with development of cutting-edge analytical methods, more HMO structures are being elucidated [4]. HMOs are made of five basic monosaccharides: D-glucose, D-galactose, N-acetylglucosamine (GlcNAc), L-fucose, and N-acetylneuraminic acid (Figure 1) [5]. Almost all HMO contain a lactose motif at the reducing end, which can be elongated by either (β1,3) or (β1,4) glycosidic bonds [6]. The addition of L-fucose and N-acetyl neuraminic acid to these linear or branched chain oligosaccharides further gives rise to the diverse oligosaccharide structures observed in human milk [5,6]. These complex oligosaccharides contribute significantly to the health of infants by lowering their susceptibility towards diseases [3].
Therefore, human milk is recognized as the gold standard for human infant nutrition by the World Health Organization and the United Nations Children’s Fund [6]. It consists of all the nutritional and physiological components required for the first 6 months of an infant’s life [7]. HMOs possess prebiotic, immunomodulatory, and antagonistic properties towards pathogens and can also serve as a key support for the neurocognitive development of infants [8,9,10]. These and other potential benefits of HMOs make them attractive research targets for preventing or treating diseases in both children and adults [8]. The bioactive properties of HMOs have prompted efforts to elucidate the mechanisms of the individual HMO’s actions [8,9,10], but the sources of HMOs for such studies are limited. Due to limited access to these HMOs, there is an emerging need to synthesize these complex molecules [10].
2. HMO Production Methods
The global HMO market was valued at USD 199 million in 2022 [11]. Lower prevalence of breast-feeding around the world [12] and rising popularity of the concept of nutraceuticals has contributed to the growing market size of HMO as a food additive [13], leading to increased demand for a cheap, large-scale production platform for synthesizing HMOs.
Several successful strategies have been developed to chemically synthesize HMOs. One of the earliest chemical syntheses was developed for Lacto-N-tetraose (LNT) in 1979 [14]. Advancement in solid phase synthesis, involving the attachment of sugar molecules to solid support, enabled the synthesis of the complex hexose sugar lacto-N-hexaose (LNH) [15]. Recently, gram-scale production of LNT was achieved from cheap starting materials such as lactose, D-glucosamine, and D-galactose, which paved the way for understanding and elucidating the importance of this sugar in human milk [16]. Over the years, more than 15 different HMOs have been chemically synthesized [3]; however, the major drawback in the chemical synthesis of HMOs is the need for many protecting group manipulations, which increase with the chain length of the target oligosaccharide [17]. Poor atom economy increases the cost of reagents, and a large number of intermediate purification steps leads to low product yields [18]. Therefore, most chemical syntheses of HMOs are not cost-efficient.
Chemoenzymatic synthesis has emerged as an alternative to traditional chemical synthesis methods for the production of natural products [19]. This approach combines the flexibility of chemical synthesis methods with the high regio- and stereoselectivity of enzyme-catalyzed reactions, eliminating the need for protection and de-protection steps [20]. Recently, 31 HMOs were chemoenzymatically synthesized [21].
Building upon these findings, a multigram-scale production system method was established for LNT synthesis based on the one-pot multienzyme (OPME) strategy [22]. OPME involves performing multistep enzymatic reactions in a single reaction system thereby eliminating the need to isolate reaction intermediates [23]. The number of diverse HMOs made available through chemical and chemoenzymatic synthesis is unparalleled [24]. Nevertheless, the high cost of purified enzymes, as well as the necessary addition of similarly expensive cofactors such as ATP, makes chemoenzymatic synthesis a less economically feasible strategy for the industrial-scale production of HMOs [25].
Microorganisms can be used as whole-cell catalysts for the production of industrially relevant molecules [26,27]. Microbial cell factories utilize cheap carbon feedstocks and also possess the high specificity of biocatalysts, which makes them great candidates for industrial-scale production [28]. In addition, microbial cell factories function at relatively lower temperatures and do not require costly purified enzymes and cofactors [28]. Recent progress in synthetic biology and metabolic engineering tools have enabled microbial production platforms to be an alternative and sustainable source of chemicals [29]. About 42 HMO structures have been produced using microbial cell factories [13]. Escherichia coli has emerged as the preferred microbial host for HMO synthesis owing to its fast growth, genetic tractability, and ease of scaling up culture volumes [30]. E. coli does not possess any native glycosyltransferase enzymes to produce HMOs. However, the presence of an efficient pathway for sugar nucleotide production, a precursor for HMO biosynthesis [31,32], makes this organism a viable platform for HMO synthesis. In this review, we summarize the latest progress in the microbial production of HMOs and discuss the challenges and future opportunities in unraveling the scope of production of complex HMOs. Herein, we focus on the microbial production of five HMOs that are approved for their commercialization and several companies are currently developing their industrial-scale production methods [30].
3. HMO Production by Microbial Cell Factories
3.1. 2′-Fucosyllactose
2′-Fucosyllactose (2′-FL) is the most abundant HMO present in human milk (Figure 1) [33]. The abundance of 2′-FL has attracted a lot of attention to its potential activity [34]. 2′-FL acts as prebiotic, promotes early development of healthy Bifidobacteria-dominated gut microbiota, and provides protection against specific diarrheal diseases in infants [35]. The regulatory approval regarding the commercialization of 2′-FL produced by microbial cell factories was granted in 2016 [13]. Recently, 2′-FL has been produced on a multigram scale via chemoenzymatic and chemical synthesis strategies [36,37]. However, there are various challenges to commercializing these methods, leading to the development of inexpensive and efficient systems to produce 2′-FL.
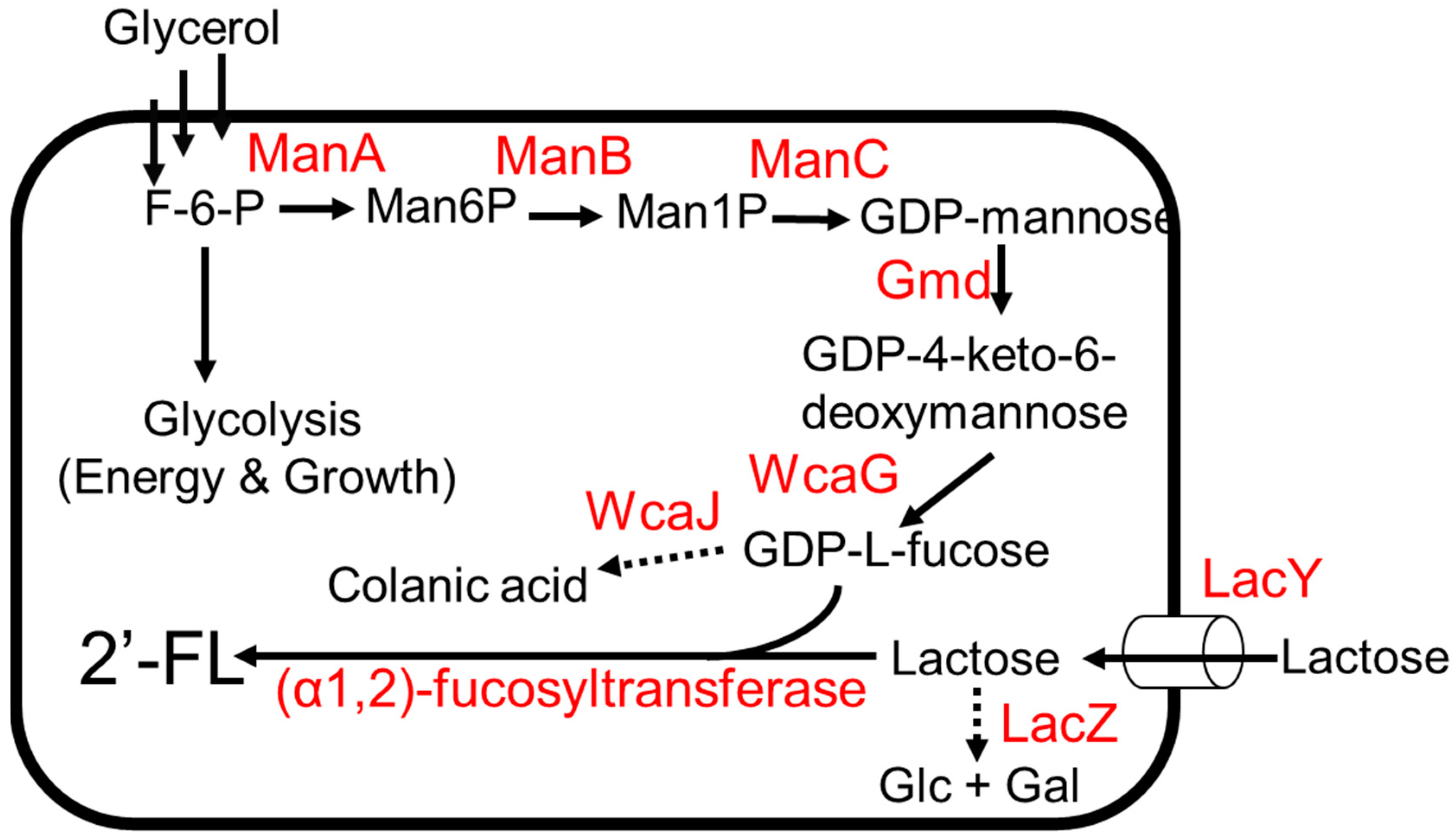
The biosynthetic pathway of 2′-FL involves the addition of GDP-L-fucose onto the galactose moiety of lactose by an (α1,2)-fucosyltransferase. Two metabolic pathways have been reported to generate GDP-L-fucose, namely the de novo pathway and the salvage pathway [38,39]. The de novo pathway is ubiquitous in both prokaryotes and eukaryotes [38]. The conversion from fructose-6-phosphate to GDP-L-fucose is carried out in five enzymatic reactions (Figure 2) [38]. Mannose-6-phosphate isomerase, ManA, converts fructose-6-phosphate to mannose-6-phosphate followed by the conversion of mannose-6-phosphate to mannose-1-phosphate by phosphomannomutase, ManB; α-D-mannose-1-phosphate guanyltransferase, ManC, converts mannose-1-phosphate to GDP-D-mannose (Figure 2). GDP-D-mannose is then converted to GDP-4-keto-6-deoxymannose by GDP-D-mannose-4,6-dehyratase, Gmd, and finally, GDP-L-fucose is produced from GDP-4-keto-6-deoxymannose by GDP-L-fucose synthetase, WcaG [38]. The salvage pathway, generally found in eukaryotes, has also been recently discovered in the gut bacterium Bacteroides fragilis [39]. This pathway involves a bifunctional fucokinase/fucose-1-phosphate guanylyltransferase (Fkp), which converts exogenous fucose to GDP-L-fucose in a two-step enzymatic reaction (Figure 3) [39]. Fkp converts L-fucose to L-fucose-1-phosphate followed by conversion of L-fucose-1-phosphate to GDP-L-fucose. For the last step in the 2′-FL biosynthetic pathway, several (α1,2)-fucosyltransferases have been identified [40].
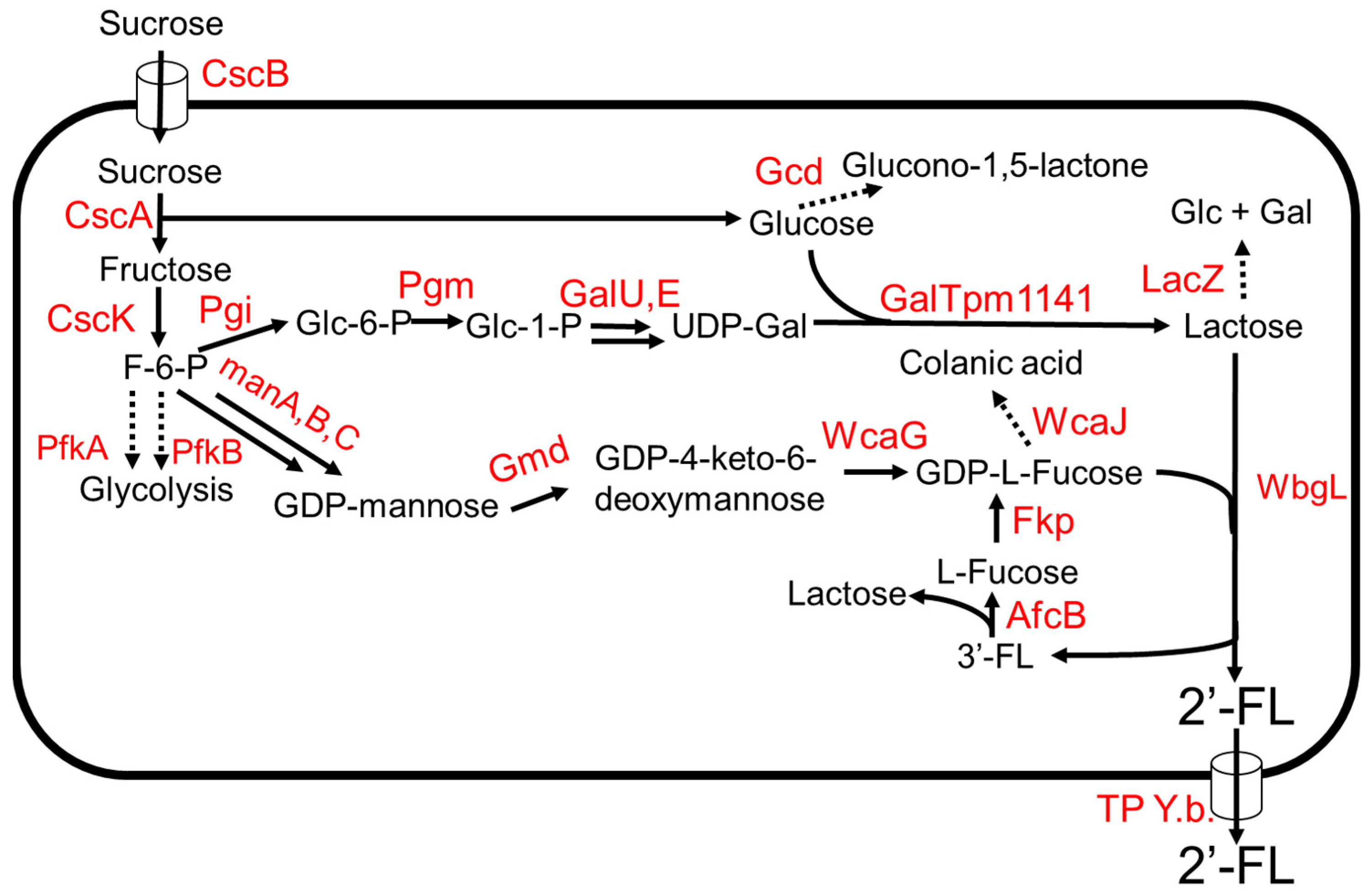
A 2′-FL production system from sucrose as the sole carbon source was developed in E. coli (Figure 3) [41]. The key intermediates, lactose, and GDP-L-fucose, were produced from sucrose (Figure 3). (β1,4)-galactosyltransferase, GalTpm1141, from Pasteurella multocida showed high specific activity towards the production of lactose from D-glucose and UDP-D-galactose [41]. Chromosomal integration of genes involved in the de novo pathway of GDP-L-fucose and cscABKR from E. coli W enabled the engineered strain to grow on sucrose and increase production of substrates, lactose, and GDP-L-fucose, in the biosynthetic pathway. The deletions of lacZ, pfkA, gnd, and wcaJ increased the production of 2′-FL. The expression of the TP Y.b. gene encoding a sugar efflux transporter from Yersinia bercovieri ATCC43970 allowed the strain to export 2′-FL out of the cell. The engineered strain expressing wbgL encoding for α1–2-fucosyltransferase from E. coli O126 [42] produced 3 g/L of 2′-FL using sucrose as the sole carbon source in a fed-batch bioreactor. Due to the promiscuity of WbgL, side products 3FL and LDFT were formed. To reduce 3FL production, afcB encoding an α1–3-fucosidase from Bifidobacterium bifidum, and the salvage pathway gene, fkp, were introduced into the production strain. AfcB hydrolyzes 3FL to produce lactose and L-fucose (Figure 3). Fkp converts the free L-fucose to GDP-L-fucose via L-fucose-1-phosphate by the salvage pathway. The strain produced ~60 g/L of 2′-FL using sucrose as the sole carbon source in a fed-batch 3 L fermenter [41].
The biosynthetic pathway of 2′-FL production via the de novo pathway for GDP-L-fucose was developed in E. coli (Figure 2) [43]. The overexpression of genes involved in de novo pathway genes using a high-copy-number plasmid was used to enhance GDP-L-fucose production. To improve lactose import, lacY was additionally expressed. To reduce lactose degradation, lacZ was deleted. The deletion of wcaJ blocks metabolic flux from GDP-L-fucose to colonic acid (Figure 2), thereby increasing GDP-L-fucose accumulation. Chromosomal integration of wbgL encoding for (α1,2)-fucosyltransferase enabled the strain to produce 2′-FL. The final production strain expressing wbgL produced 79 g/L of 2′-FL using glycerol and lactose as carbon sources in a 3 L fermenter [43].
The futC gene encoding for (α1,2)-fucosyltransferase from Helicobacter pylori [40] was used for 2′-FL production in E. coli due to its high specificity towards lactose [44]. The expression levels of genes involved in lactose uptake and de novo L-fucose assimilation were modulated to improve the accumulation of lactose, GDP-L-fucose, and the intermediates for 2′-FL production. The biosynthetic pathway for 2′-FL utilized one NADPH and GTP molecule. To reduce cell burden, the balance of NADPH and GTP was achieved by the overexpression of zwf, pntAB, and gsk, leading to a significant increase in 2′-FL production. The final strain produced 22 g/L of 2′-FL from lactose in a 3 L fermenter [44].
Saccharomyces cerevisiae was engineered to produce 2′-FL from xylose and lactose [45]. The use of yeast as a host organism avoids the risk of bacteriophage infection during large-scale production, possible endotoxin contamination, and unfavorable consumer perception. A lactose transporter gene LAC12 from Kluyveromyces lactis and wbgL from E. coli O126 were introduced into S. cerevisiae. To enable the de novo production of GDP-L-fucose, the gmd and wcaG genes from E. coli were introduced to S. cerevisiae. The engineered strain produced 26 g/L of 2′-FL from xylose and lactose in a fed-batch 1L fermentor [45]. The putative (α1,2)-fucosyltransferase FutBc from Bacillus cereus was also used to produce 2′-FL. Expression of futBC enhanced 2′-FL production by 1.8-fold compared to futC from H. pylori. The futBC genes along with LAC12 was introduced into S. cerevisiae. The strain produced 27 g/L of 2′-FL from lactose and sucrose in a fed-batch 5 L bioreactor [46].
3.2. LDFT (Lactodifucotetraose)
LDFT is the most abundant fucosylated HMO and over the course of the first year of lactation, LDFT concentration in human milk is ~0.43 g/L [47]. LDFT can be metabolized by immunomodulatory bacteria, promoting healthy gut microbiome in infants [48]. The lack of availability of LDFT is a major roadblock to further understanding its health benefits. The unique challenge posed by the chemistry of LDFT structure, two different glycosidic bonds between fucose and lactose, makes it a difficult molecule to synthesize chemically. LDFT produced by microorganisms was given regulatory approval as Generally Recognized As Safe (GRAS) in 2018 [30]. The presence of a promiscuous enzyme, (α1,3/4)-fucosyltransferase (3/4FT) from H. pylori UA948 was successfully isolated and expressed in E. coli [49].
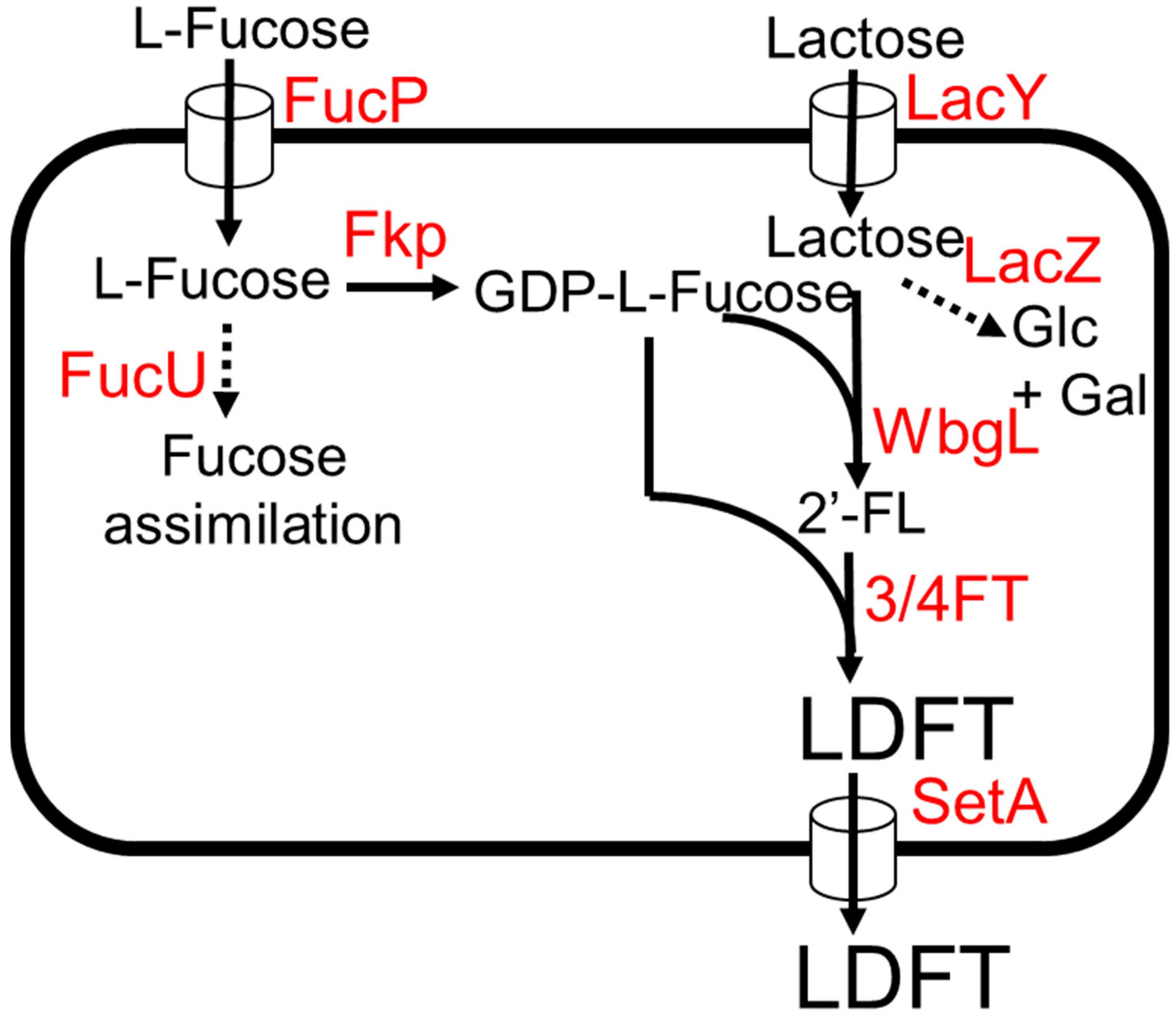
E. coli was engineered to produce LDFT from lactose and L-fucose [50]. The biosynthetic pathway of LDFT production involves the addition of GDP-L-fucose onto both glucose and galactose moiety of lactose by an (α1,3)-fucosyltransferase and (α1,2)-fucosyltransferase, respectively (Figure 4) [50]. It has been shown that α1–3/4-fucosyltransferase (3/4FT) from H. pylori UA948 can use both non-fucosylated and α1–2-fucosylated galactosyl oligosaccharides as substrates [22,51], while for (α1,2)-fucosyltransferases, WbgL is selective towards lactose and other non-fucosylated galactosyl oligosaccharide acceptor substrates [22,42]. For the production of GDP-L-fucose, fkp encoding for bifunctional fucokinase/fucose-1-phosphate from Bacteroides fragilis was expressed (Figure 4). The lacZ and fucU genes were deleted to remove competing pathways. To enhance the uptake rate of L-fucose and lactose into the cell, the sugar transporter genes lacY and fucP were additionally expressed. The final strain produced 5.1 g/L of LDFT from 3 g/L lactose and 3 g/L L-fucose, achieving 91% of the theoretical maximum yield in test tube conditions. The production of monofucoside side products (2′-FL and 3-FL) was very low compared to LDFT, suggesting the potential viability of this production system for large-scale LDFT production [50].
3.3. 3′ and 6′-Sialyllactose
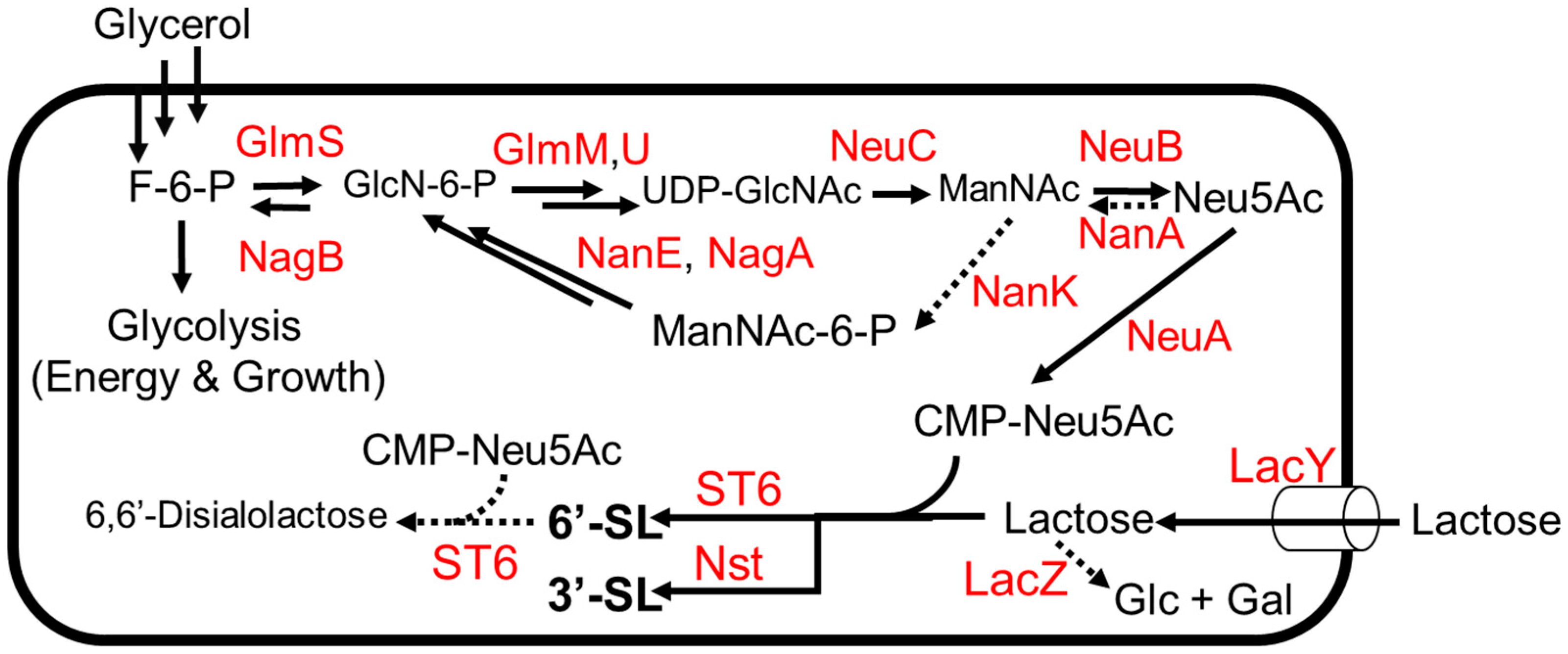
Sialylated HMOs are of interest due to their roles as dietary sources of sialic acid, which plays a role in preventing intestinal infections and promoting healthy brain function in humans [52]. As with other HMOs, sialylated HMOs have been investigated as biosynthetic targets due to the prohibitive cost of chemical synthesis, which suffers from low yields due to the many protection and de-protection steps needed to achieve a stereo- and regiospecific product. The sialic acid N-acetylneuraminic acid (Neu5Ac) is rare in nature and difficult to chemically synthesize for similar reasons, meaning that an efficient biosynthesis of any sialyllactose must also include the biosynthesis of the Neu5Ac moiety itself (Figure 5). An exogenous pathway was used to biosynthesize CMP-Neu5Ac in combination with a glycosyltransferase to yield 3′-sialolactose [53]. In this study, three exogenous genes (neuABC) from Campylobacter jejuni ATCC 4348 were expressed in E. coli K12, thus completing a pathway from glycolysis to CMP-Neu5Ac (Figure 6). Genes for competing pathways (nanA, nanK) were deleted to direct carbon flux towards CMP-Neu5Ac. Finally, an (α2,3)-sialyltransferase gene from Neisseria meningitidis was introduced to convert CMP-Neu5Ac to 3′-sialyllactose in the presence of lactose, which serves as the donor substrate for the glycosyltransferase enzyme (Figure 5). The engineered strain was grown to high density (OD600~100) on mineral media with a continuous supply of glycerol and lactose, producing 15.5 g/L of 3′-sialyllactose (3′-SL) after 70 h in a 2 L fermentor.
Efforts to biosynthesize 6′-sialyllactose (6′-SL) have required more optimization than efforts to biosynthesize 3′-SL due to the relative promiscuity of (α2,6)-sialyltransferase enzymes (Figure 5). The sialyltransferase gene in the 3′-SL-producing strain described above [53] was substituted with an (α2,6)-sialyltransferase from Photobacterium sp. JT-ISH-224 [54]. The (α2,6)-sialyltransferase was chosen because it was the most specific (α2,6)-sialyltransferase known at the time [54]. When this strain was grown on glycerol and lactose, it was found that lactose was exclusively conjugated with 2-keto-3-deoxyoctonate (KDO). To combat the apparent promiscuity of the sialyltransferase enzyme, a variety of strategies were used. First, the supply of substrate for the sialyltransferase was increased by placing the neuABC operon under a stronger promoter on a plasmid with a higher copy number. With this change in place, KDO-lactose was no longer formed, and the strain yielded the intended product 6′-sialyllactose and the side product 6,6′-disalyllactose (Figure 5). Hypothesizing that this disialylated product was formed by a side activity of the sialyltransferase, the strain was fed a constant supply of lactose to outcompete 6′-SL as a substrate. The strain produced 16 g L−1 of 6′-siallylactose after 47 h with a negligible amount of 6–6′-disialyllactose in a 2 L fermentor.
3.4. LNT (Lacto-N-Tetraose)
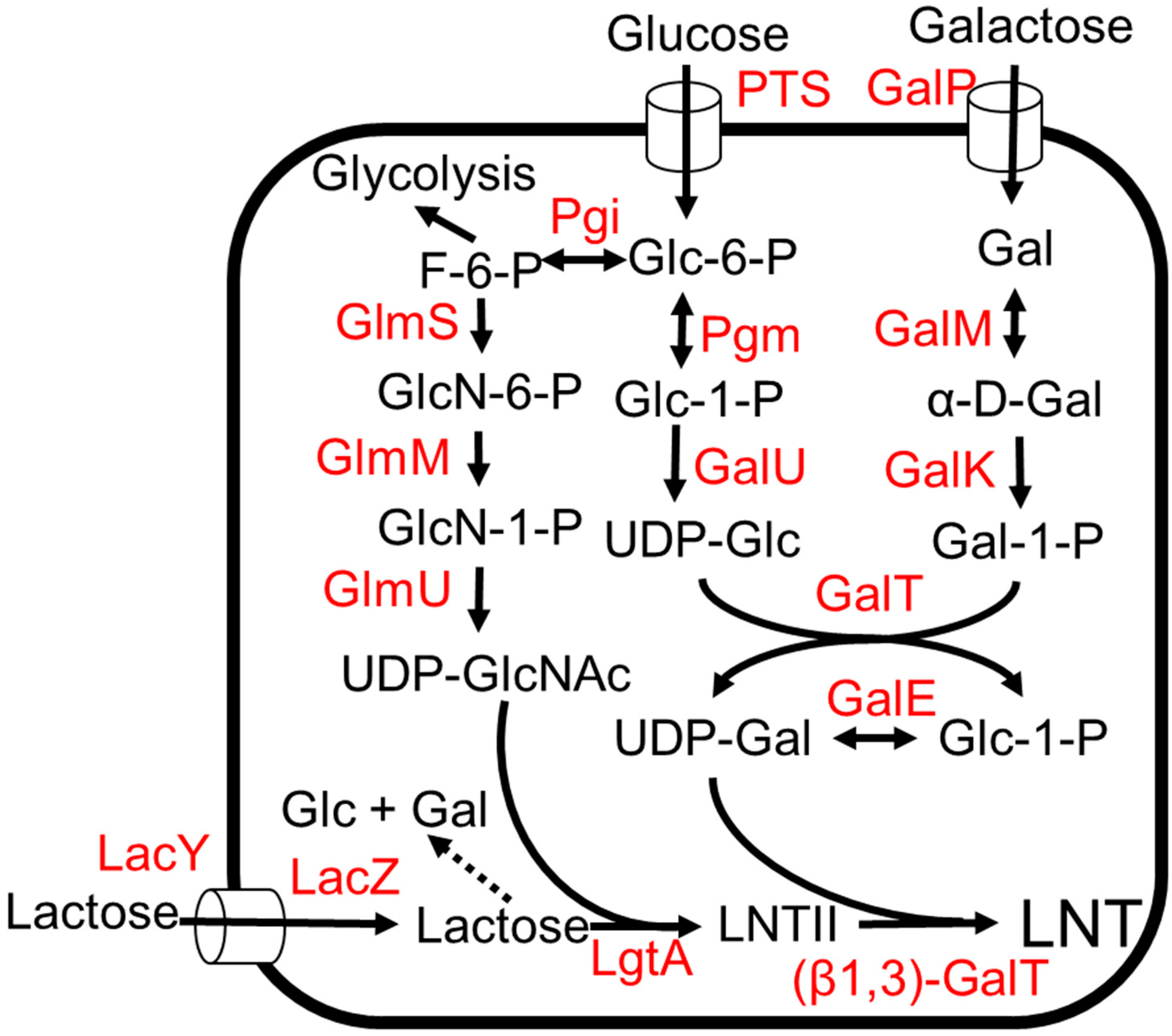
LNT represents a core structure for most HMO structures as most HMOs are derived from different attachments of L-fucose and the sialic acid group (Figure 1) [3]. It is also the most abundant solid component in HMOs, comprising 6% (w/w) of total HMOs [55]. Efficient one-pot multienzyme (OPME) strategies have been developed for the production of LNT [22,56]. The biosynthetic pathway of LNT involves the production of UDP-N-GlcNAc and the production of UDP-galactose (Figure 6). Both these intermediates are added to a lactose unit to form LNT (Figure 6) [55,57,58,59]. LNT produced by microorganisms was given regulatory approval as GRAS for commercialization in 2016 [13].
Structural elucidation of LNT-like units in the lipopolysaccharides of Neisseria meningtidis led to the discovery of (β1,3)-N-acetylglucosaminyltransferase enzyme, LgtA, which can add a galactose unit to lactose to produce LNT II, the precursor of LNT (Figure 6) [60,61]. The O antigen structure of E. coli O55:H7 consists of a repeating polysaccharide containing Gal b1,3GlcNAc. The (β1,3)-galactosyltransferase, WbgO, which is involved in forming the repeating polysaccharide, was identified and shown to convert LNT II to LNT [22,62]. E. coli K12 was engineered to produce LNT (Figure 6) [57]. To avoid the use of antibiotics and increase gene stability, chromosomal integration of the heterologous glycosyltransferase gene instead of a plasmid expression system was used. The lgtA gene encoding for (β1,3) N-acetylglucosaminyltransferase, LgtA, from Neisseria meningtidis was integrated into the lacZYA locus, thereby eliminating the lactose-utilization pathway. The wbgO gene encoding b1–3-galactosyltransferase, WbgO, from E. coli O55:H7 was integrated into the xylAB locus in E. coli. To increase lactose uptake, the lacY gene encoding for lactose transporter LacY was additionally expressed under a strong inducible Ptac promoter [63]. In a shaken flask, this strain produced 0.2 g/L of LNT from glucose and lactose [57]. However, the strain produced 9 times more LNT II than LNT. This suggests that the availability of UDP-galactose and the activity of WbgO would be the bottleneck for the LNT production [57]. With galactose instead of glucose, the strain produced 0.8 g/L LNT in a fed batch bioreactor [58]. The presence of glucose inhibits lactose utilization by catabolite repression and inducer exclusion, so galactose is a better feedstock for the production of LNT [64].
To increase the UDP-galactose pool, the genes (galE-galT-galK) of the Leloir pathway [65] were overexpressed (Figure 6). The deletion of ugd encoding for UDP-glucose-6-dehyrdogenase has been previously shown to increase UDP-galactose accumulation in E. coli [65]. Thus, ugd was deleted in the production strain, and the final strain produced 32 g/L of LNT from glycerol and lactose in a fed-batch bioreactor [55].
Several sugar efflux transporters have been identified in E. coli [66]. These transporters relieve the osmotic stress due to the accumulation of sugars by exporting sugars out of the cells. The presence of LNT II in the production media suggests that E. coli has a transporter for LNT II [59]. Fourteen native sugar transporters were screened to identify an LNT transporter. This study showed that SetA is the major LNT II transporter in E. coli. The deletion of setA increased LNT titers from 2.19 g/L to 2.96 g/L and exogenous LNT II concentration decreased from 0.52 g/L to 0.23 g/L [59].
4. Challenges
When considering the application of whole-cell biocatalysts in the production of food additives, food safety must be thoroughly considered. Food products based on Gram-negative bacteria such as Escherichia coli increase the likelihood of lipopolysaccharide contamination [67]. Lipopolysaccharides are a structural component of bacterial cells and are also known as endotoxins [68]. Bacterial lipopolysaccharides have been associated with a number of diseases, including liver damage, neurological degeneration, gut inflammation, and diabetes [67]. The purification of HMOs produced by E. coli to remove these endotoxins is a crucial safeguard and will remain a major hurdle for the commercialization of these microbial production platforms [68].
Sugar transporters can be useful tools in enhancing HMO production, but they can also hamper biosynthesis. The presence of several native sugar transporters in E. coli, which can export HMO precursors out of the cell, is not optimal for the production of more complex HMOs that have many intermediates [66]. The identification and characterization of these sugar transporters are very important for HMO production not only to minimize substrate loss but to selectively export target HMOs out of the cell. In addition, more complex HMOs may not have any known transporters, and the biosynthesis of these targets could be limited by the buildup of the product in the cytosol. The study of new and existing sugar transporters is necessary to progress the field of HMO biosynthesis.
There is a lack of protein-engineering strategies to increase the activity, specificity, and expression of glycosyltransferases in microbial hosts. Underperformance in these areas leads to low titers and the formation of side products [53,57]. For example, in HMO biosynthetic pathways involving the expression of multiple glycosyltransferases, the use of highly specific enzymes can help in controlling the order of glycosylation and avoiding the formation of side products.
5. Conclusions and Perspectives
Progress in the field of analytical methods continues to reveal new molecules contributing to the vast diversity of HMOs. With the increasing beneficial effects of HMOs being reported, there is a growing demand for the large-scale production of HMOs. Here, we summarize the latest research in the microbial production of HMOs. Microbial cell factories function at relatively low temperatures, do not require costly purified enzymes and cofactors, utilize cheap carbon feedstock, and also possess the high specificity of biocatalysts, which makes them great candidates for industrial-scale production. The lack of knowledge on the sugar transporters for HMOs is a major bottleneck in the production of long-chain HMOs. Furthermore, the unintentional export of intermediates in the biosynthetic pathway will lead to low titer and yield. The fundamental basis for metabolically engineering HMO-producing strains is the activity of glycosyltransferase enzymes, and therefore, there is a need for protein engineering to increase the specificity and solubility of these heterologous genes in microbial production hosts. Nevertheless, microbial production platforms possess great potential to produce a wide range of HMOs on a large scale.
Author Contributions
D.S.K.P., S.R.P. and S.A. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the National Institutes of Health (GM145842 to S.A.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
We would like to thank Jake Gonzales for critical reading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jéquier, E. Carbohydrates as a Source of Energy. Am. J. Clin. Nutr. 1994, 59, 682S–685S. [Google Scholar] [CrossRef] [PubMed]
- Boquien, C.-Y. Human Milk: An Ideal Food for Nutrition of Preterm Newborn. Front. Pediatr. 2018, 6, 295. [Google Scholar] [CrossRef] [PubMed]
- Chen, X. Human Milk Oligosaccharides (HMOS): Structure, Function, and Enzyme-Catalyzed Synthesis. Adv. Carbohydr. Chem. Biochem. 2015, 72, 113–190. [Google Scholar] [CrossRef] [PubMed]
- Auer, F.; Jarvas, G.; Guttman, A. Recent Advances in the Analysis of Human Milk Oligosaccharides by Liquid Phase Separation Methods. J. Chromatogr. B 2021, 1162, 122497. [Google Scholar] [CrossRef] [PubMed]
- Soyyılmaz, B.; Mikš, M.H.; Röhrig, C.H.; Matwiejuk, M.; Meszaros-Matwiejuk, A.; Vigsnæs, L.K. The Mean of Milk: A Review of Human Milk Oligosaccharide Concentrations throughout Lactation. Nutrients 2021, 13, 2737. [Google Scholar] [CrossRef]
- Garwolińska, D.; Namieśnik, J.; Kot-Wasik, A.; Hewelt-Belka, W. Chemistry of Human Breast Milk—A Comprehensive Review of the Composition and Role of Milk Metabolites in Child Development. J. Agric. Food Chem. 2018, 66, 11881–11896. [Google Scholar] [CrossRef]
- Bode, L.; McGuire, M.; Rodriguez, J.M.; Geddes, D.T.; Hassiotou, F.; Hartmann, P.E.; McGuire, M.K. It’s Alive: Microbes and Cells in Human Milk and Their Potential Benefits to Mother and Infant. Adv. Nutr. 2014, 5, 571–573. [Google Scholar] [CrossRef]
- Wiciński, M.; Sawicka, E.; Gębalski, J.; Kubiak, K.; Malinowski, B. Human Milk Oligosaccharides: Health Benefits, Potential Applications in Infant Formulas, and Pharmacology. Nutrients 2020, 12, 266. [Google Scholar] [CrossRef]
- Hill, D.R.; Chow, J.M.; Buck, R.H. Multifunctional Benefits of Prevalent HMOs: Implications for Infant Health. Nutrients 2021, 13, 3364. [Google Scholar] [CrossRef]
- Christian, P.; Smith, E.R.; Lee, S.E.; Vargas, A.J.; Bremer, A.A.; Raiten, D.J. The Need to Study Human Milk as a Biological System. Am. J. Clin. Nutr. 2021, 113, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Markets and Markets Website, Human Milk Oligosaccharides (HMO) Market. Available online: https://www.marketsandmarkets.com/Market-Reports/human-milk-oligosaccharides-hmo-market-220134357.html (accessed on 20 December 2022).
- Neves, P.A.; Barros, A.J.; Baker, P.; Piwoz, E.; Santos, T.M.; Gatica-Domínguez, G.; Vaz, J.S.; Rollins, N.; Victora, C.G. Consumption of Breast Milk, Formula and Other Non-Human Milk by Children Aged under 2 Years: Analysis of Eighty-Six Low- and Middle-Income Countries. Public Health Nutr. 2022, 25, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.; Lane, J.A.; van Sinderen, D.; Hickey, R.M. From Lab Bench to Formulated Ingredient: Characterization, Production, and Commercialization of Human Milk Oligosaccharides. J. Funct. Foods 2020, 72, 104052. [Google Scholar] [CrossRef]
- Takamura, T.; Chiba, T.; Ishihara, H.; Tejima, S. Chemical Modification of Lactose. XIII. Synthesis of Lacto-N-Tetraose. Chem. Pharm. Bull. 1979, 27, 1497–1499. [Google Scholar] [CrossRef]
- Knerr, L.; Schmidt, R.R. Solid-Phase Synthesis of a Branched Hexasaccharide Related to Lacto-N-Hexaose. Eur. J. Org. Chem. 2000, 2000, 2803–2808. [Google Scholar] [CrossRef]
- Craft, K.M.; Townsend, S.D. Synthesis of Lacto-N-Tetraose. Carbohydr. Res. 2017, 440–441, 43–50. [Google Scholar] [CrossRef]
- Das, R.; Mukhopadhyay, B. Chemical O-Glycosylations: An Overview. ChemistryOpen 2016, 5, 401–433. [Google Scholar] [CrossRef]
- McKay, M.J.; Nguyen, H.M. Recent Advances in Transition Metal-Catalyzed Glycosylation. ACS Catal. 2012, 2, 1563–1595. [Google Scholar] [CrossRef]
- Chakrabarty, S.; Romero, E.O.; Pyser, J.B.; Yazarians, J.A.; Narayan, A.R.H. Chemoenzymatic Total Synthesis of Natural Products. Acc. Chem. Res. 2021, 54, 1374–1384. [Google Scholar] [CrossRef]
- Muthana, S.; Cao, H.; Chen, X. Recent Progress in Chemical and Chemoenzymatic Synthesis of Carbohydrates. Curr. Opin. Chem. Biol. 2009, 13, 573–581. [Google Scholar] [CrossRef]
- Xiao, Z.; Guo, Y.; Liu, Y.; Li, L.; Zhang, Q.; Wen, L.; Wang, X.; Kondengaden, S.M.; Wu, Z.; Zhou, J.; et al. Chemoenzymatic Synthesis of a Library of Human Milk Oligosaccharides. J. Org. Chem. 2016, 81, 5851–5865. [Google Scholar] [CrossRef] [Green Version]
- McArthur, J.B.; Yu, H.; Chen, X. A Bacterial Β1–3-Galactosyltransferase Enables Multigram-Scale Synthesis of Human Milk Lacto- N -Tetraose (LNT) and Its Fucosides. ACS Catal. 2019, 9, 10721–10726. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Xu, H.; Fang, J.; Zhang, X. Enzymatic and Chemoenzymatic Synthesis of Human Milk Oligosaccharides and Derivatives. Carbohydr. Polym. 2022, 291, 119564. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.L.; Townsend, S.D. Synthesis as an Expanding Resource in Human Milk Science. J. Am. Chem. Soc. 2021, 143, 11277–11290. [Google Scholar] [CrossRef]
- Zeuner, B.; Teze, D.; Muschiol, J.; Meyer, A.S. Synthesis of Human Milk Oligosaccharides: Protein Engineering Strategies for Improved Enzymatic Transglycosylation. Molecules 2019, 24, 2033. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.L.; Desai, S.H.; Atsumi, S. Microbial Production of Scent and Flavor Compounds. Curr. Opin. Biotechnol. 2016, 37, 8–15. [Google Scholar] [CrossRef]
- Chubukov, V.; Mukhopadhyay, A.; Petzold, C.J.; Keasling, J.D.; Martín, H.G. Synthetic and Systems Biology for Microbial Production of Commodity Chemicals. Npj Syst. Biol. Appl. 2016, 2, 16009. [Google Scholar] [CrossRef]
- Cho, J.S.; Kim, G.B.; Eun, H.; Moon, C.W.; Lee, S.Y. Designing Microbial Cell Factories for the Production of Chemicals. JACS Au 2022, 2, 1781–1799. [Google Scholar] [CrossRef]
- Tashiro, Y.; Rodriguez, G.M.; Atsumi, S. 2-Keto Acids Based Biosynthesis Pathways for Renewable Fuels and Chemicals. J. Ind. Microbiol. Biotechnol. 2015, 42, 361–373. [Google Scholar] [CrossRef]
- Bych, K.; Mikš, M.H.; Johanson, T.; Hederos, M.J.; Vigsnæs, L.K.; Becker, P. Production of HMOs Using Microbial Hosts—From Cell Engineering to Large Scale Production. Curr. Opin. Biotechnol. 2019, 56, 130–137. [Google Scholar] [CrossRef]
- Woo, J.E.; Seong, H.J.; Lee, S.Y.; Jang, Y.-S. Metabolic Engineering of Escherichia coli for the Production of Hyaluronic Acid From Glucose and Galactose. Front. Bioeng. Biotechnol. 2019, 7, 351. [Google Scholar] [CrossRef] [Green Version]
- Byun, S.-G.; Kim, M.-D.; Lee, W.-H.; Lee, K.-J.; Han, N.S.; Seo, J.-H. Production of GDP-L-Fucose, L-Fucose Donor for Fucosyloligosaccharide Synthesis, in Recombinant Escherichia coli. Appl. Microbiol. Biotechnol. 2007, 74, 768–775. [Google Scholar] [CrossRef]
- Hegar, B.; Wibowo, Y.; Basrowi, R.W.; Ranuh, R.G.; Sudarmo, S.M.; Munasir, Z.; Atthiyah, A.F.; Widodo, A.D.; Supriatmo; Kadim, M.; et al. The Role of Two Human Milk Oligosaccharides, 2′-Fucosyllactose and Lacto-N-Neotetraose, in Infant Nutrition. Pediatr. Gastroenterol. Hepatol. Nutr. 2019, 22, 330–340. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Duysburgh, C.; Vazquez, E.; Chow, J.; Buck, R.; Marzorati, M. 2′-Fucosyllactose Alters the Composition and Activity of Gut Microbiota from Formula-Fed Infants Receiving Complementary Feeding in a Validated Intestinal Model. J. Funct. Foods 2019, 61, 103484. [Google Scholar] [CrossRef]
- Vandenplas, Y.; Berger, B.; Carnielli, V.P.; Ksiazyk, J.; Lagström, H.; Sanchez Luna, M.; Migacheva, N.; Mosselmans, J.-M.; Picaud, J.-C.; Possner, M.; et al. Human Milk Oligosaccharides: 2′-Fucosyllactose (2′-FL) and Lacto-N-Neotetraose (LNnT) in Infant Formula. Nutrients 2018, 10, 1161. [Google Scholar] [CrossRef] [PubMed]
- Agoston, K.; Hederos, M.J.; Bajza, I.; Dekany, G. Kilogram Scale Chemical Synthesis of 2′-Fucosyllactose. Carbohydr. Res. 2019, 476, 71–77. [Google Scholar] [CrossRef]
- Li, C.; Wu, M.; Gao, X.; Zhu, Z.; Li, Y.; Lu, F.; Qin, H.-M. Efficient Biosynthesis of 2′-Fucosyllactose Using an In Vitro Multienzyme Cascade. J. Agric. Food Chem. 2020, 68, 10763–10771. [Google Scholar] [CrossRef]
- Wan, L.; Zhu, Y.; Li, W.; Zhang, W.; Mu, W. Combinatorial Modular Pathway Engineering for Guanosine 5′-Diphosphate-l-Fucose Production in Recombinant Escherichia coli. J. Agric. Food Chem. 2020, 68, 5668–5675. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Han, D.; Pan, Y.; Wang, S.; Fang, J.; Wang, P.; Liu, X. Enhancing GDP-Fucose Production in Recombinant Escherichia coli by Metabolic Pathway Engineering. Enzyme Microb. Technol. 2015, 69, 38–45. [Google Scholar] [CrossRef]
- Seydametova, E.; Yu, J.; Shin, J.; Park, Y.; Kim, C.; Kim, H.; Yu, S.H.; Park, Y.; Kweon, D.-H. Search for Bacterial A1,2-Fucosyltransferases for Whole-Cell Biosynthesis of 2′-Fucosyllactose in Recombinant Escherichia coli. Microbiol. Res. 2019, 222, 35–42. [Google Scholar] [CrossRef]
- Parschat, K.; Schreiber, S.; Wartenberg, D.; Engels, B.; Jennewein, S. High-Titer De Novo Biosynthesis of the Predominant Human Milk Oligosaccharide 2′-Fucosyllactose from Sucrose in Escherichia coli. ACS Synth. Biol. 2020, 9, 2784–2796. [Google Scholar] [CrossRef]
- Engels, L.; Elling, L. WbgL: A Novel Bacterial A1,2-Fucosyltransferase for the Synthesis of 2′-Fucosyllactose. Glycobiology 2014, 24, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, Y.; Wan, L.; Chen, R.; Zhang, W.; Mu, W. High-Level De Novo Biosynthesis of 2′-Fucosyllactose by Metabolically Engineered Escherichia coli. J. Agric. Food Chem. 2022, 70, 9017–9025. [Google Scholar] [CrossRef]
- Li, M.; Li, C.; Hu, M.; Zhang, T. Metabolic Engineering Strategies of de Novo Pathway for Enhancing 2′-Fucosyllactose Synthesis in Escherichia coli. Microb. Biotechnol. 2022, 15, 1561–1573. [Google Scholar] [CrossRef]
- Lee, J.W.; Kwak, S.; Liu, J.-J.; Yu, S.; Yun, E.J.; Kim, D.H.; Liu, C.; Kim, K.H.; Jin, Y.-S. Enhanced 2′-Fucosyllactose Production by Engineered Saccharomyces cerevisiae Using Xylose as a Co-Substrate. Metab. Eng. 2020, 62, 322–329. [Google Scholar] [CrossRef]
- Xu, M.; Meng, X.; Zhang, W.; Shen, Y.; Liu, W. Improved Production of 2′-Fucosyllactose in Engineered Saccharomyces cerevisiae Expressing a Putative α-1, 2-Fucosyltransferase from Bacillus Cereus. Microb. Cell Factories 2021, 20, 165. [Google Scholar] [CrossRef]
- Chaturvedi, P.; Warren, C.D.; Altaye, M.; Morrow, A.L.; Ruiz-Palacios, G.; Pickering, L.K.; Newburg, D.S. Fucosylated Human Milk Oligosaccharides Vary between Individuals and over the Course of Lactation. Glycobiology 2001, 11, 365–372. [Google Scholar] [CrossRef]
- Yu, Z.-T.; Chen, C.; Newburg, D.S. Utilization of Major Fucosylated and Sialylated Human Milk Oligosaccharides by Isolated Human Gut Microbes. Glycobiology 2013, 23, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.A.; Wang, G.; Palcic, M.M.; Taylor, D.E. Cloning and Characterization of the α(1,3/4) Fucosyltransferase of Helicobacter Pylori. J. Biol. Chem. 2000, 275, 4988–4994. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, L.; Bai, Y.; Yu, H.; McArthur, J.B.; Chen, X.; Atsumi, S. Microbial Production of Human Milk Oligosaccharide Lactodifucotetraose. Metab. Eng. 2021, 66, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Li, Y.; Wu, Z.; Li, L.; Zeng, J.; Zhao, C.; Wu, Y.; Tasnima, N.; Wang, J.; Liu, H.; et al. Pylori A1–3/4-Fucosyltransferase (Hp3/4FT)-Catalyzed One-Pot Multienzyme (OPME) Synthesis of Lewis Antigens and Human Milk Fucosides. Chem. Commun. 2017, 53, 11012–11015. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Brand-Miller, J. The Role and Potential of Sialic Acid in Human Nutrition. Eur. J. Clin. Nutr. 2003, 57, 1351–1369. [Google Scholar] [CrossRef]
- Fierfort, N.; Samain, E. Genetic Engineering of Escherichia coli for the Economical Production of Sialylated Oligosaccharides. J. Biotechnol. 2008, 134, 261–265. [Google Scholar] [CrossRef]
- Drouillard, S.; Mine, T.; Kajiwara, H.; Yamamoto, T.; Samain, E. Efficient Synthesis of 6′-Sialyllactose, 6,6′-Disialyllactose, and 6′-KDO-Lactose by Metabolically Engineered E. coli Expressing a Multifunctional Sialyltransferase from the Photobacterium Sp. JT-ISH-224. Carbohydr. Res. 2010, 345, 1394–1399. [Google Scholar] [CrossRef]
- Hu, M.; Li, M.; Miao, M.; Zhang, T. Engineering Escherichia coli for the High-Titer Biosynthesis of Lacto-N-Tetraose. J. Agric. Food Chem. 2022, 70, 8704–8712. [Google Scholar] [CrossRef]
- Yao, W.; Yan, J.; Chen, X.; Wang, F.; Cao, H. Chemoenzymatic Synthesis of Lacto-N-Tetrasaccharide and Sialyl Lacto-N-Tetrasaccharides. Carbohydr. Res. 2015, 401, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Baumgärtner, F.; Conrad, J.; Sprenger, G.A.; Albermann, C. Synthesis of the Human Milk Oligosaccharide Lacto-N-Tetraose in Metabolically Engineered, Plasmid-Free E. Coli. Chembiochem Eur. J. Chem. Biol. 2014, 15, 1896–1900. [Google Scholar] [CrossRef] [PubMed]
- Baumgärtner, F.; Sprenger, G.A.; Albermann, C. Galactose-Limited Fed-Batch Cultivation of Escherichia coli for the Production of Lacto-N-Tetraose. Enzyme Microb. Technol. 2015, 75–76, 37–43. [Google Scholar] [CrossRef]
- Sugita, T.; Koketsu, K. Transporter Engineering Enables the Efficient Production of Lacto-N-Triose II and Lacto-N-Tetraose in Escherichia coli. J. Agric. Food Chem. 2022, 70, 5106–5114. [Google Scholar] [CrossRef] [PubMed]
- Blixt, O.; van Die, I.; Norberg, T.; van den Eijnden, D.H. High-Level Expression of the Neisseria Meningitidis LgtA Gene in Escherichia coli and Characterization of the Encoded N-Acetylglucosaminyltransferase as a Useful Catalyst in the Synthesis of GlcNAcβ1→3Gal and GalNAcβ1→3Gal Linkages. Glycobiology 1999, 9, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Jennings, M.P.; Hood, D.W.; Peak, I.R.; Virji, M.; Moxon, E.R. Molecular Analysis of a Locus for the Biosynthesis and Phase-Variable Expression of the Lacto-N-Neotetraose Terminal Lipopolysaccharide Structure in Neisseria Meningitidis. Mol. Microbiol. 1995, 18, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xia, C.; Li, L.; Guan, W.; Pettit, N.; Zhang, H.; Chen, M.; Wang, P.G. Characterization and Synthetic Application of a Novel Β1,3-Galactosyltransferase from Escherichia coli O55:H7. Bioorg. Med. Chem. 2009, 17, 4910–4915. [Google Scholar] [CrossRef] [PubMed]
- de Boer, H.A.; Comstock, L.J.; Vasser, M. The Tac Promoter: A Functional Hybrid Derived from the Trp and Lac Promoters. Proc. Natl. Acad. Sci. USA 1983, 80, 21–25. [Google Scholar] [CrossRef]
- Santillán, M.; Mackey, M.C. Quantitative Approaches to the Study of Bistability in the Lac Operon of Escherichia coli. J. R. Soc. Interface 2008, 5 (Suppl. S1), S29–S39. [Google Scholar] [CrossRef]
- Lee, S.J.; Trostel, A.; Adhya, S. Metabolite Changes Signal Genetic Regulatory Mechanisms for Robust Cell Behavior. MBio 2014, 5, e00972-13. [Google Scholar] [CrossRef] [PubMed]
- Koita, K.; Rao, C.V. Identification and analysis of the putative pentose sugar efflux transporters in Escherichia coli. Plos ONE 2012, 7, e43700. [Google Scholar] [CrossRef]
- Wassenaar, T.M.; Zimmermann, K. Lipopolysaccharides in Food, Food Supplements, and Probiotics: Should We Be Worried? Eur. J. Microbiol. Immunol. 2018, 8, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Ongkudon, C.M.; Chew, J.H.; Liu, B.; Danquah, M.K. Chromatographic Removal of Endotoxins: A Bioprocess Engineer’s Perspective. Int. Sch. Res. Not. 2012, 2012, e649746. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Structures of Human milk oligosaccharides described in this review. HMOs are made of five basic monosaccharides: D-glucose, D-galactose, L-fucose, N-acetyl-D-glucosamine (GlcNAc), and N-acetylneuraminic acid (Neu5A).
Figure 1.
Structures of Human milk oligosaccharides described in this review. HMOs are made of five basic monosaccharides: D-glucose, D-galactose, L-fucose, N-acetyl-D-glucosamine (GlcNAc), and N-acetylneuraminic acid (Neu5A).

Figure 2.
The 2′FL production from glycerol and lactose. Dotted arrows indicate competing pathways. Abbreviations: ManA, mannose-6-phosphate isomerase; ManB, phosphomannomutase; ManC, α-D-mannose-1-phosphate guanyltransferase; Gmd, GDP-D-mannose-4,6-dehyratase; WcaG, GDP-L-fucose synthetase; LacZ, β-galactosidase; WcaJ, UDP-glucose:undecaprenyl-phosphate glucose-1-phosphate transferase; LacY, lactose permease.
Figure 2.
The 2′FL production from glycerol and lactose. Dotted arrows indicate competing pathways. Abbreviations: ManA, mannose-6-phosphate isomerase; ManB, phosphomannomutase; ManC, α-D-mannose-1-phosphate guanyltransferase; Gmd, GDP-D-mannose-4,6-dehyratase; WcaG, GDP-L-fucose synthetase; LacZ, β-galactosidase; WcaJ, UDP-glucose:undecaprenyl-phosphate glucose-1-phosphate transferase; LacY, lactose permease.

Figure 3.
The 2′FL production from sucrose. Dotted arrows indicate competing pathways. Abbreviations: ManA, mannose-6-phosphate isomerase; ManB, phosphomannomutase; ManC, α-D-mannose-1-phosphate guanyltransferase; Gmd, GDP-D-mannose-4,6-dehyratase; WcaG, GDP-L-fucose synthetase; Pgi, glucose-6-phosphate isomerase; Pgm, phosphoglucomutase; GalU, UTP—glucose-1-phosphate uridylyltransferase; GalE, UDP-glucose 4-epimerase; WcaJ, UDP-glucose:undecaprenyl-phosphate glucose-1-phosphate transferase; Gcd, quinoprotein glucose dehydrogenase; LacZ, β-galactosidase; Fkp, bifunctional fucokinase/fucose-1-phosphate guanylyltransferase; AfcB, α1–3-fucosidase; GalTpm1141, b1–4-galactosyltransferase; WbgL, (α1,2)-fucosyltransferase; TP Y.b., sugar efflux transporter; CscB, sucrose permease; CscA, sucrose hydrolase; CscK, fructokinase; PfkA, 6-phosphofructokinase 1; PfkB, 6-phosphofructokinase 2.
Figure 3.
The 2′FL production from sucrose. Dotted arrows indicate competing pathways. Abbreviations: ManA, mannose-6-phosphate isomerase; ManB, phosphomannomutase; ManC, α-D-mannose-1-phosphate guanyltransferase; Gmd, GDP-D-mannose-4,6-dehyratase; WcaG, GDP-L-fucose synthetase; Pgi, glucose-6-phosphate isomerase; Pgm, phosphoglucomutase; GalU, UTP—glucose-1-phosphate uridylyltransferase; GalE, UDP-glucose 4-epimerase; WcaJ, UDP-glucose:undecaprenyl-phosphate glucose-1-phosphate transferase; Gcd, quinoprotein glucose dehydrogenase; LacZ, β-galactosidase; Fkp, bifunctional fucokinase/fucose-1-phosphate guanylyltransferase; AfcB, α1–3-fucosidase; GalTpm1141, b1–4-galactosyltransferase; WbgL, (α1,2)-fucosyltransferase; TP Y.b., sugar efflux transporter; CscB, sucrose permease; CscA, sucrose hydrolase; CscK, fructokinase; PfkA, 6-phosphofructokinase 1; PfkB, 6-phosphofructokinase 2.

Figure 4.
The LDFT production from L-fucose and lactose. Dotted arrows indicate competing pathways. Abbreviations: FucP, L-fucose:H+ symporter; Fkp, bifunctional fucokinase/fucose-1-phosphate guanylyltransferase; FucU, L-fucose mutarotase; LacY, lactose permease; LacZ, β-galactosidase; WbgL, (α1,2)-fucosyltransferase; ¾FT, (α1,3/4)-fucosyltransferase; SetA, sugar efflux transporter A.
Figure 4.
The LDFT production from L-fucose and lactose. Dotted arrows indicate competing pathways. Abbreviations: FucP, L-fucose:H+ symporter; Fkp, bifunctional fucokinase/fucose-1-phosphate guanylyltransferase; FucU, L-fucose mutarotase; LacY, lactose permease; LacZ, β-galactosidase; WbgL, (α1,2)-fucosyltransferase; ¾FT, (α1,3/4)-fucosyltransferase; SetA, sugar efflux transporter A.

Figure 5.
The 6′-SL and 3′-SL production from glycerol and lactose. Dotted arrows indicate competing pathways. Abbreviations: GlmS, L-glutamine—D-fructose-6-phosphate aminotransferase; GlmM, phosphoglucosamine mutase; GlmU, fused N-acetylglucosamine-1-phosphate uridyltransferase and glucosamine-1-phosphate acetyltransferase; LacZ, β-galactosidase; LacY, lactose permease; NagB, glucosamine-6-phosphate deaminase; NanE, N-acetylmannosamine-6-phosphate 2-epimerase; NagA, N-acetylglucosamine-6-phosphate deacetylase; NanK, N-acetylmannosamine kinase; NeuC, GlcNAc-6-phosphate 2 epimerase; NeuB, sialic acid synthase; NeuA, CMP-Neu5Ac synthetase; NanA, N-acetylneuraminate lyase; ST6, (α2,6)-Sialyltransferase; Nst, (α2,3) NeuAc transferase.
Figure 5.
The 6′-SL and 3′-SL production from glycerol and lactose. Dotted arrows indicate competing pathways. Abbreviations: GlmS, L-glutamine—D-fructose-6-phosphate aminotransferase; GlmM, phosphoglucosamine mutase; GlmU, fused N-acetylglucosamine-1-phosphate uridyltransferase and glucosamine-1-phosphate acetyltransferase; LacZ, β-galactosidase; LacY, lactose permease; NagB, glucosamine-6-phosphate deaminase; NanE, N-acetylmannosamine-6-phosphate 2-epimerase; NagA, N-acetylglucosamine-6-phosphate deacetylase; NanK, N-acetylmannosamine kinase; NeuC, GlcNAc-6-phosphate 2 epimerase; NeuB, sialic acid synthase; NeuA, CMP-Neu5Ac synthetase; NanA, N-acetylneuraminate lyase; ST6, (α2,6)-Sialyltransferase; Nst, (α2,3) NeuAc transferase.

Figure 6.
The LNT production from lactose and glucose or galactose. Dotted arrows indicate competing pathways. Abbreviations: PTS, phosphotransferase system; GlmS, L-glutamine—D-fructose-6-phosphate aminotransferase; GlmM, phosphoglucosamine mutase; GlmU, fused N-acetylglucosamine-1-phosphate uridyltransferase and glucosamine-1-phosphate acetyltransferase; Pgi, glucose-6-phosphate isomerase; Pgm, phosphoglucomutase; GalU, UTP—glucose-1-phosphate uridylyltransferase; GalT, galactose-1-phosphate uridylyltransferase; GalE, UDP-glucose 4-epimerase; GalK, galactokinase; GalM, galactose-1-epimerase; GalP, galactose:H+ symporter; LacY, lactose permease; LacZ, β-galactosidase; LgtA, (β1,3)-N-acetylglucosaminyltransferase; β1,3-GalT, (β1,3)-galactosyltransferase.
Figure 6.
The LNT production from lactose and glucose or galactose. Dotted arrows indicate competing pathways. Abbreviations: PTS, phosphotransferase system; GlmS, L-glutamine—D-fructose-6-phosphate aminotransferase; GlmM, phosphoglucosamine mutase; GlmU, fused N-acetylglucosamine-1-phosphate uridyltransferase and glucosamine-1-phosphate acetyltransferase; Pgi, glucose-6-phosphate isomerase; Pgm, phosphoglucomutase; GalU, UTP—glucose-1-phosphate uridylyltransferase; GalT, galactose-1-phosphate uridylyltransferase; GalE, UDP-glucose 4-epimerase; GalK, galactokinase; GalM, galactose-1-epimerase; GalP, galactose:H+ symporter; LacY, lactose permease; LacZ, β-galactosidase; LgtA, (β1,3)-N-acetylglucosaminyltransferase; β1,3-GalT, (β1,3)-galactosyltransferase.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Palur, D.S.K.; Pressley, S.R.; Atsumi, S. Microbial Production of Human Milk Oligosaccharides. Molecules 2023, 28, 1491. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28031491
AMA Style
Palur DSK, Pressley SR, Atsumi S. Microbial Production of Human Milk Oligosaccharides. Molecules. 2023; 28(3):1491. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28031491
Chicago/Turabian StylePalur, Dileep Sai Kumar, Shannon R. Pressley, and Shota Atsumi. 2023. "Microbial Production of Human Milk Oligosaccharides" Molecules 28, no. 3: 1491. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28031491