Lentivirus-Mediated ERK2 siRNA Reduces Joint Capsule Fibrosis in a Rat Model of Post-Traumatic Joint Contracture

Abstract
:1. Introduction
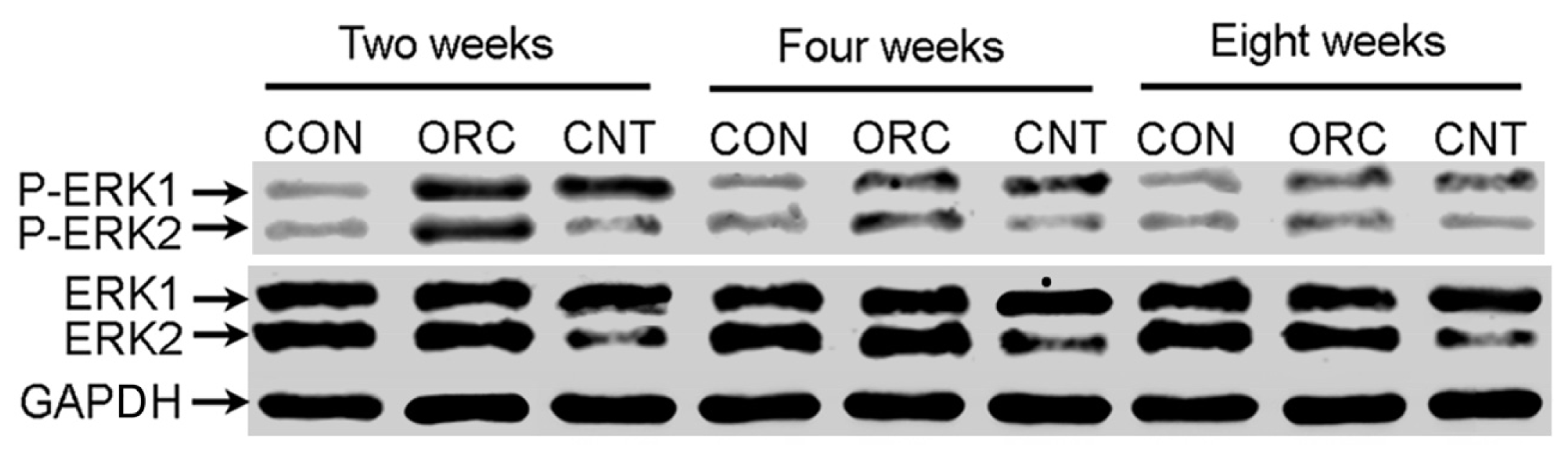
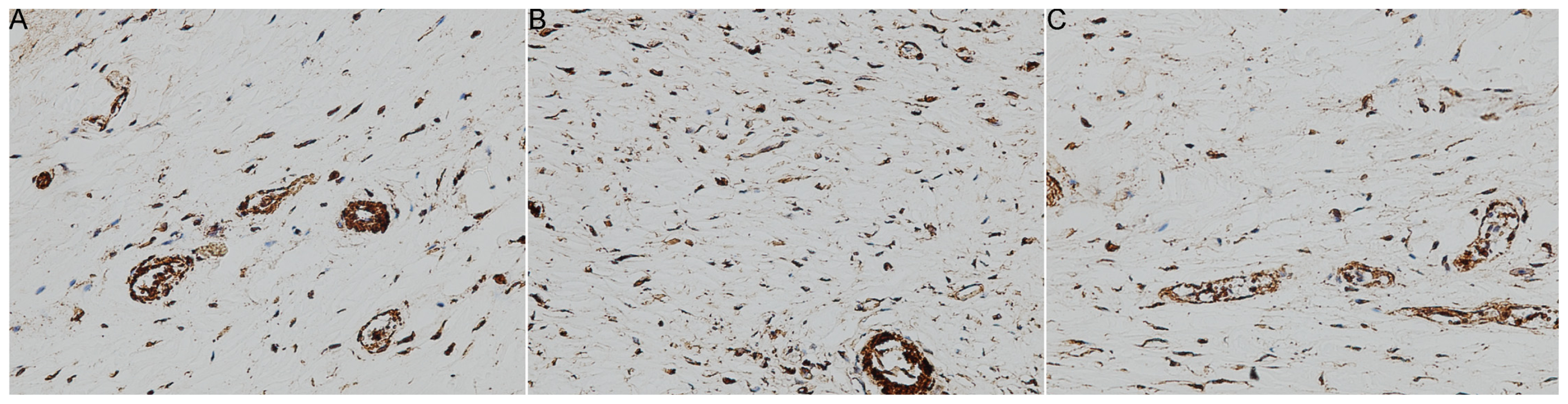
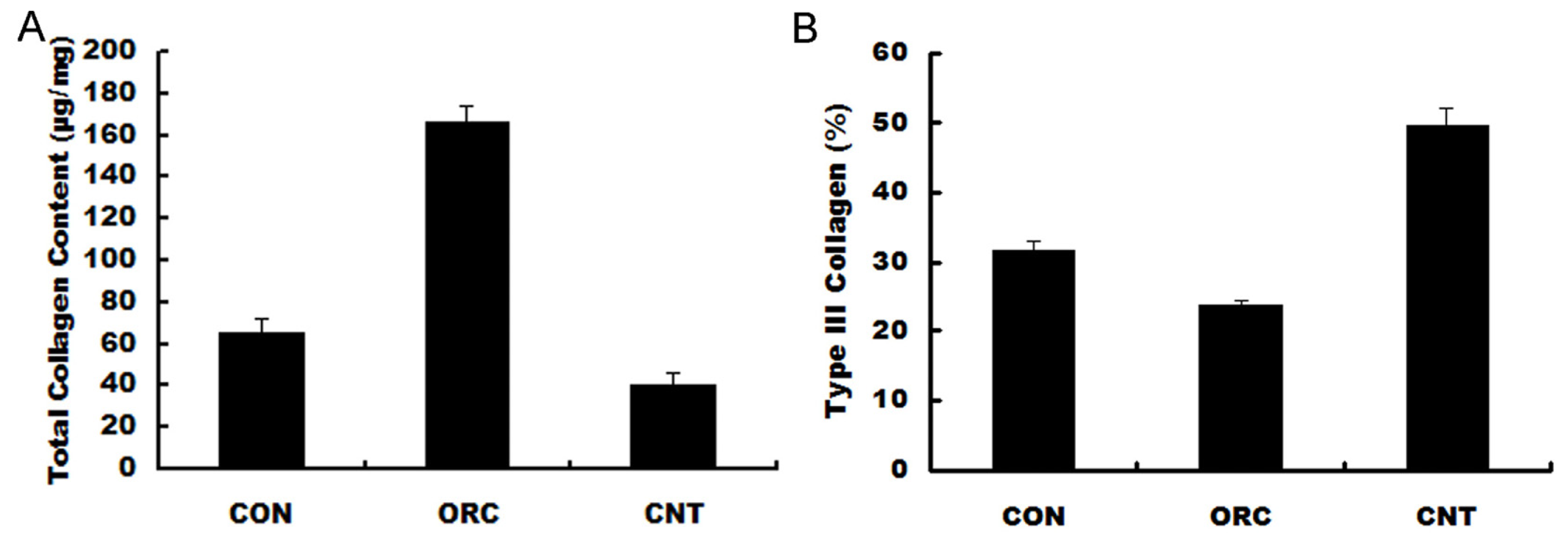
2. Results and Discussion
3. Experiment Section
3.2. Group Allocation
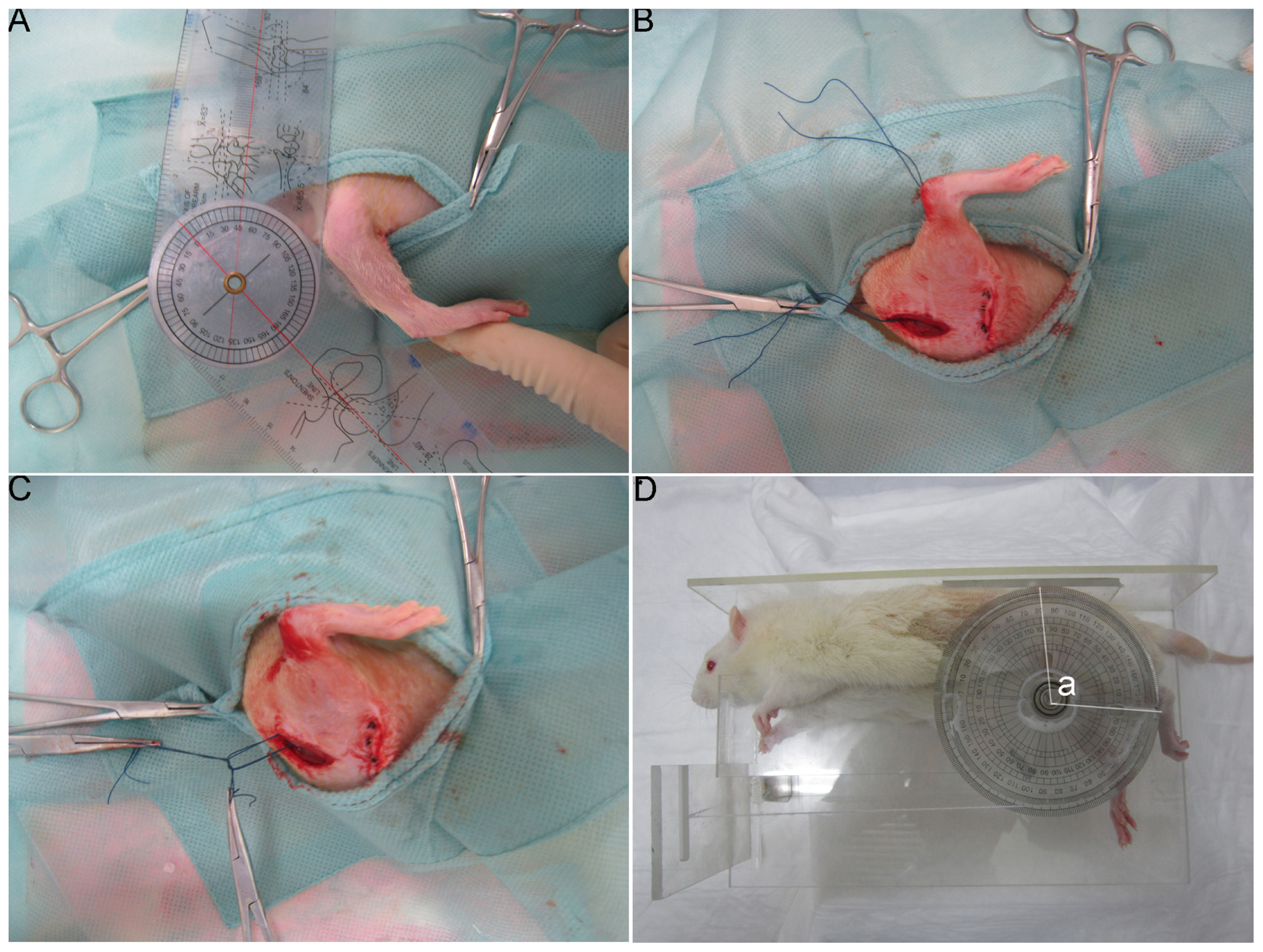
3.3. Joint Interventions
3.4. In Vivo Bioluminescence Assay
3.5. Western Blot
3.6. Immunohistochemistry
3.7. Measurement of the Flexion Contracture Angle
3.8. Biochemical Analysis
3.9. Statistics
4. Conclusions






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number | Myofibroblasts (hpf) | ap | bp |
|---|---|---|---|---|
| CON | 8 | 81 ± 15 | NA | <0.01 |
| ORC | 8 | 253 ± 47 | <0.01 | NA |
| CNT | 7 | 63 ± 14 | >0.05 | <0.01 |
| Group | Number | Contracture angles (°) | ap | bp |
|---|---|---|---|---|
| CON | 8 | 28.1 ± 3.3 | NA | <0.01 |
| ORC | 7 | 124.0 ± 12.3 | <0.01 | NA |
| CNT | 8 | 41.2 ± 5.6 | >0.05 | <0.01 |
Acknowledgments
Conflicts of Interest
References
- Doornberg, J.N.; van Duijn, P.J.; Linzel, D.; Ring, D.C.; Zurakowski, D.; Marti, R.K.; Kloen, P. Surgical treatment of intra-articular fractures of the distal part of the humerus. Functional outcome after twelve to thirty years. J. Bone Joint Surg. Am 2007, 89, 1524–1532. [Google Scholar]
- Forthman, C.; Henket, M.; Ring, D.C. Elbow dislocation with intra-articular fracture: The results of operative treatment without repair of the medial collateral ligament. J. Hand Surg. Am 2007, 32, 1200–1209. [Google Scholar]
- Lindenhovius, A.L.; Jupiter, J.B. The posttraumatic stiff elbow: A review of the literature. J. Hand Surg. Am 2007, 32, 1605–1623. [Google Scholar]
- Cohen, M.S.; Hastings, I.H. Post-traumatic contracture of the elbow. Operative release using a lateral collateral ligament sparing approach. J. Bone Joint Surg. Br 1998, 80B, 805–812. [Google Scholar]
- Gelinas, J.J.; Faber, K.J.; Patterson, S.D.; King, G.J. The effectiveness of turnbuckle splinting for elbow contractures. J. Bone Joint Surg. Br 2000, 82, 74–78. [Google Scholar]
- Hayashi, M.; Sekiya, H.; Takatoku, K.; Kariya, Y.; Hoshino, Y. Experimental model of knee contracture in extension: Its prevention using a sheet made from hyaluronic acid and carboxymethylcellulose. Knee Surg. Sports Traumatol. Arthrosc 2004, 12, 545–551. [Google Scholar]
- Usuba, M.; Miyanaga, Y.; Miyakawa, S.; Maeshima, T.; Shirasaki, Y. Effect of heat in increasing the range of knee motion after the development of a joint contracture: An experiment with an animal model. Arch. Phys. Med. Rehabil 2006, 87, 247–253. [Google Scholar]
- Monument, M.J.; Hart, D.A.; Befus, A.D.; Salo, P.T.; Zhang, M.; Hildebrand, K.A. The mast cell stabilizer ketotifen fumarate lessens contracture severity and myofibroblast hyperplasia: A study of a rabbit model of posttraumatic joint contractures. J. Bone Joint Surg. Am 2010, 92, 1468–1477. [Google Scholar]
- Elbashir, S.M.; Harborth, J.; Lendeckel, W.; Yalcin, A.; Weber, K.; Tuschl, T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 2001, 411, 494–498. [Google Scholar]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar]
- Hannon, G.J. RNA interference. Nature 2002, 418, 244–251. [Google Scholar]
- Wang, Q.; Peng, Z.; Xiao, S.; Geng, S.; Yuan, J.; Li, Z. RNAi-mediated inhibition of COL1A1 and COL3A1 in human skin fibroblasts. Exp. Dermatol 2007, 16, 611–617. [Google Scholar]
- Nakamura, H.; Siddiqui, S.S.; Shen, X.; Malik, A.B.; Pulido, J.S.; Kumar, N.M.; Yue, B.Y. Interference targeting transforming growth factor-beta type II receptor suppresses ocular inflammation and fibrosis. Mol. Vis 2004, 10, 703–711. [Google Scholar]
- Wang, Z.; Gao, Z.; Shi, Y.; Sun, Y.; Lin, Z.; Jiang, H.; Hou, T.; Wang, Q.; Yuan, X.; Zhu, X.; et al. Inhibition of Smad3 expression decreases collagen synthesis in keloid disease fibroblasts. J. Plast. Reconstr. Aesthet. Surg 2007, 60, 1193–1199. [Google Scholar]
- Li, F.; Ruan, H.; Fan, C.; Zeng, B.; Wang, C.; Wang, X. Efficient inhibition of the formation of joint adhesions by ERK2 small interfering RNAs. Biochem. Biophys. Res. Commun 2010, 391, 795–799. [Google Scholar]
- Bunker, T.D.; Reilly, J.; Baird, K.S.; Hamblen, D.L. Expression of growth factors, cytokines and matrix metalloproteinases in frozen shoulder. J. Bone Joint Surg. Br 2000, 82, 768–773. [Google Scholar]
- Kirk, T.Z.; Mark, M.E.; Chua, C.C.; Chua, B.H.; Mayes, M.D. Myofibroblasts from scleroderma skin synthesize elevated levels of collagen and tissue inhibitor of metalloproteinase (TIMP-1) with two forms of TIMP-1. J. Biol. Chem 1995, 270, 3423–3428. [Google Scholar]
- Harunari, N.; Zhu, K.Q.; Armendariz, R.T.; Deubner, H.; Muangman, P.; Carrougher, G.J.; Isik, F.F.; Gibran, N.S.; Engrav, L.H. Histology of the thick scar on the female, red Duroc pig: Final similarities to human hypertrophic scar. Burns 2006, 32, 669–677. [Google Scholar]
- Fine, A.; Poliks, C.F.; Smith, B.D.; Goldstein, R.H. The accumulation of type I collagen mRNAs in human embryonic lung fibroblasts stimulated by transforming growth factor-beta. Connect. Tissue Res 1990, 24, 237–247. [Google Scholar]
- Hildebrand, K.A.; Zhang, M.; Hart, D.A. Myofibroblast upregulators are elevated in joint capsules in posttraumatic contractures. Clin. Orthop. Relat. Res 2007, 456, 85–91. [Google Scholar]
- Benzakour, O.; Merzak, A.; Dooghe, Y.; Pironin, M.; Lawrence, D.; Vigier, P. Transforming growth factor beta stimulates mitogenically mouse NIH3T3 fibroblasts and those cells transformed by the EJ-H ras oncogene. Growth Factors 1992, 6, 265–275. [Google Scholar]
- Li, F.; Fan, C.; Cheng, T.; Jiang, C.; Zeng, B. Efficient inhibition of fibroblast proliferation and collagen expression by ERK2 siRNAs. Biochem. Biophys. Res. Commun 2009, 382, 259–263. [Google Scholar]
- Lois, C.; Hong, E.J.; Pease, S.; Brown, E.J.; Baltimore, D. Germline transmission and tissue-specific expression of transgenes delivered by lentiviral vectors. Science 2002, 295, 868–872. [Google Scholar]
- Pfeifer, A.; Ikawa, M.; Dayn, Y.; Verma, I.M. Transgenesis by lentiviral vectors: Lack of gene silencing in mammalian embryonic stem cells and preimplantation embryos. Proc. Natl. Acad. Sci. USA 2002, 99, 2140–2145. [Google Scholar]
- Lai, Z.; Brady, R.O. Gene transfer into the central nervous system in vivo using a recombinant lentivirus vector. J. Neurosci. Res 2002, 67, 363–371. [Google Scholar]
- Yu, X.; Zhan, X.; D’Costa, J.; Tanavde, V.M.; Ye, Z.; Peng, T.; Malehorn, M.T.; Yang, X.; Civin, C.I.; Cheng, L. Lentiviral vectors with two independent internal promoters transfer high-level expression of multiple transgenes to human hematopoietic stem-progenitor cells. Mol. Ther 2003, 7, 827–838. [Google Scholar]
- Gallay, S.H.; Richards, R.R.; O’Driscoll, S.W. Intraarticular capacity and compliance of stiff and normal elbows. Arthroscopy 1993, 9, 9–13. [Google Scholar]
- Cohen, M.S.; Schimmel, D.R.; Masuda, K.; Hastings, H., 2nd; Muehleman, C. Structural and biochemical evaluation of the elbow capsule after trauma. J. Shoulder Elbow Surg 2007, 16, 484–490. [Google Scholar]
- Germscheid, N.M.; Hildebrand, K.A. Regional variation is present in elbow capsules after injury. Clin. Orthop. Relat. Res 2006, 450, 219–224. [Google Scholar]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechanoregulation of connective tissue remodeling. Nat. Rev. Mol. Cell Biol 2002, 3, 349–363. [Google Scholar]
- Hildebrand, K.A.; Zhang, M.; Hart, D.A. High rate of joint capsule matrix turnover in chronic human elbow contractures. Clin. Orthop. Relat. Res 2005, 439, 228–234. [Google Scholar]
- Forrest, L. Current concepts in soft connective tissue wound healing. Br. J. Surg 1983, 70, 133–140. [Google Scholar]
- Clore, J.N.; Cohen, I.K.; Diegelmann, R.F. Quantitation of collagen types I and III during wound healing in rat skin. Proc. Soc. Exp. Biol. Med 1979, 161, 337–340. [Google Scholar]
- Parry, D.A.; Barnes, G.R.; Craig, A.S. A comparison of the size distribution of collagen fibrils in connective tissues as a function of age and possible relation between fibril size distribution and mechanical properties. Proc. R. Soc. Lond 1978, 203, 305–321. [Google Scholar]
- Hildebrand, K.A.; Sutherland, C.; Zhang, M. Rabbit knee model of post-traumatic joint contractures: The long-term natural history of motion loss and myofibroblasts. J. Orthop. Res 2004, 22, 313–320. [Google Scholar]
- Hildebrand, K.A.; Zhang, M.; Hart, D.A. Joint capsule matrix turnover in a rabbit model of chronic joint contractures: Correlation with human contractures. J. Orthop. Res 2006, 24, 1036–1043. [Google Scholar]
- Monument, M.J.; Hart, D.A.; Befus, A.D.; Salo, P.T.; Zhang, M.; Hildebrand, K.A. The mast cell stabilizer ketotifen reduces joint capsule fibrosis in a rabbit model of post-traumatic joint contractures. Inflamm. Res 2012, 61, 285–292. [Google Scholar]
- Nesterenko, S.; Morrey, M.E.; Abdel, M.P.; An, K.N.; Steinmann, S.P.; Morrey, B.F.; Sanchez-Sotelo, J. New rabbit knee model of posttraumatic joint contracture: Indirect capsular damage induces a severe contracture. J. Orthop. Res 2009, 27, 1028–1032. [Google Scholar]
- Edwards, C.A.; O’Brien, W.D., Jr. Modified assay for determination of hydroxyproline in a tissue hydrolyzate. Clin. Chim. Acta 1980, 104, 161–167. [Google Scholar]
- Woessner, J.F., Jr. The determination of hydroxyproline in tissue and protein samples containing small proportions of this imino acid. Arch. Biochem. Biophys 1961, 93, 440–447. [Google Scholar]
- Chan, D.; Cole, W.G. Quantitation of type I and III collagens using electrophoresis of alpha chains and cyanogen bromide peptides. Anal. Biochem 1984, 139, 322–328. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, F.; Liu, S.; Fan, C. Lentivirus-Mediated ERK2 siRNA Reduces Joint Capsule Fibrosis in a Rat Model of Post-Traumatic Joint Contracture. Int. J. Mol. Sci. 2013, 14, 20833-20844. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms141020833
Li F, Liu S, Fan C. Lentivirus-Mediated ERK2 siRNA Reduces Joint Capsule Fibrosis in a Rat Model of Post-Traumatic Joint Contracture. International Journal of Molecular Sciences. 2013; 14(10):20833-20844. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms141020833
Chicago/Turabian StyleLi, Fengfeng, Shen Liu, and Cunyi Fan. 2013. "Lentivirus-Mediated ERK2 siRNA Reduces Joint Capsule Fibrosis in a Rat Model of Post-Traumatic Joint Contracture" International Journal of Molecular Sciences 14, no. 10: 20833-20844. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms141020833