Gypenosides Protected the Neural Stem Cells in the Subventricular Zone of Neonatal Rats that Were Prenatally Exposed to Ethanol

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Ethanol Treatment Increased the Lethality and Abnormality of the Neonatal Rats

2.2. Blood Alcohol Concentration
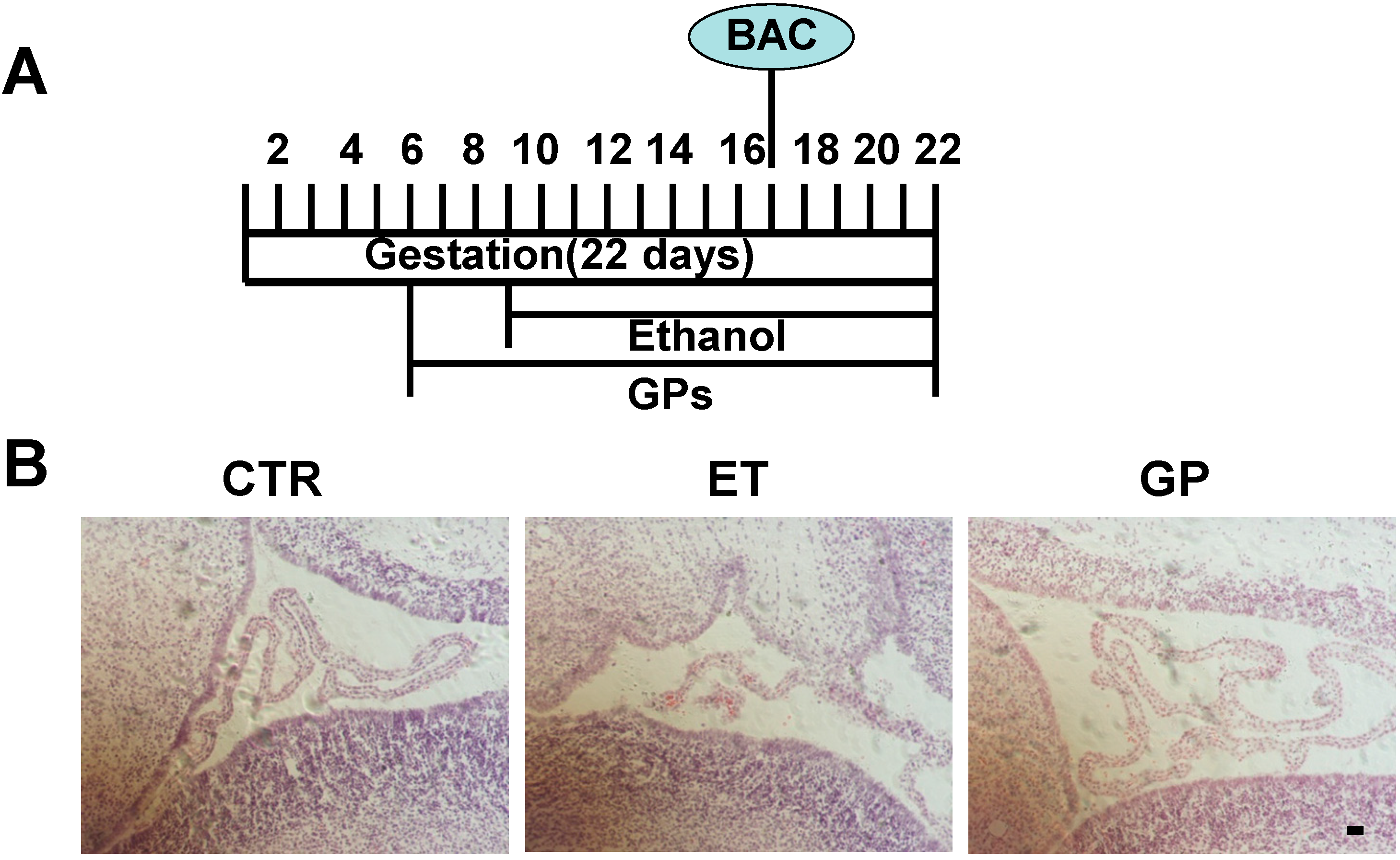
2.3. GPs Protected the Cell Morphology of SVZ in the Neonatal Brains Exposed to Ethanol as Embryos
2.4. GPs Increased the Cell Proliferation of Neural Stem Cells in the SVZ, Reversing the Inhibition by Ethanol

2.5. GPs Promoted the Cell Differentiation of Neural Stem Cells in the SVZ that Were Inhibited by Ethanol


2.6. Discussion
3. Experimental Section
3.1. Experimental Materials
3.2. Study Design for Animal Models
3.3. H&E Staining
3.4. Immunohistochemistry
3.5. Microscopy Analysis
3.6. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jones, K.L.; Smith, D.W.; Ulleland, C.N.; Streissguth, P. Pattern of malformation in offspring of chronic alcoholic mothers. Lancet 1973, 1, 1267–1271. [Google Scholar] [PubMed]
- Moore, E.S.; Ward, R.E.; Jamison, P.L.; Morris, C.A.; Bader, P.I.; Hall, B.D. The subtle facial signs of prenatal exposure to alcohol: An anthropometric approach. J. Pediatr. 2001, 139, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.W. Development of the Central Nervous System: Effects of Alcohol and Opiates; Wiley-Liss: New York, NY, USA, 1992. [Google Scholar]
- Luo, J.; Miller, M.W. Growth factor-mediated neural proliferation: target of ethanol toxicity. Brain Res. Rev. 1998, 27, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, T.J.; Richards, L.J.; Bartlett, P.F. The regulation of neural precursor cells within the mammalian brain. Mol. Cell. Neurosci. 1995, 6, 2–15. [Google Scholar] [PubMed]
- Miranda, R.C. MicroRNAs and fetal brain development: Implications for ethanol teratology during the second trimester period of neurogenesis. Front. Genet. 2012, 3, 77. [Google Scholar] [PubMed]
- Hao, H.N.; Parker, G.C.; Zhao, J.; Barami, K.; Lyman, W.D. Human neural stem cells are more sensitive than astrocytes to ethanol exposure. Alcohol. Clin. Exp. Res. 2003, 27, 1310–1317. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Li, B.S.; Maric, D.; Zhao, W.Q.; Lin, H.J.; Zhang, L.; Pant, H.C.; Barker, J.L. Ethanol blocks both basic fibroblast growth factor- and carbachol-mediated neuroepithelial cell expansion with differential effects on carbachol-activated signaling pathways. Neuroscience 2003, 118, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Tochitani, S.; Sakata-Haga, H.; Fukui, Y. Embryonic exposure to ethanol disturbs regulation of mitotic spindle orientation via GABA(A) receptors in neural progenitors in ventricular zone of developing neocortex. Neurosci. Lett. 2010, 472, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Vangipuram, S.D.; Lyman, W.D. Ethanol alters cell fate of fetal human brain-derived stem and progenitor cells. Alcohol. Clin. Exp. Res. 2010, 34, 1574–1583. [Google Scholar] [CrossRef] [PubMed]
- Vangipuram, S.D.; Lyman, W.D. Ethanol affects differentiation-related pathways and suppresses Wnt signaling protein expression in human neural stem cells. Alcohol. Clin. Exp. Res. 2012, 36, 788–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.C.; Balaraman, Y.; Teng, M.; Liu, Y.; Singh, R.P.; Nephew, K.P. Alcohol alters DNA methylation patterns and inhibits neural stem cell differentiation. Alcohol. Clin. Exp. Res. 2011, 35, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Redila, V.A.; Olson, A.K.; Swann, S.E.; Mohades, G.; Webber, A.J.; Weinberg, J.; Christie, B.R. Hippocampal cell proliferation is reduced following prenatal ethanol exposure but can be rescued with voluntary exercise. Hippocampus 2006, 16, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.Y.; Allan, A.M.; Cunningham, L.A. Moderate fetal alcohol exposure impairs the neurogenic response to an enriched environment in adult mice. Alcohol. Clin. Exp. Res. 2005, 29, 2053–2062. [Google Scholar] [CrossRef] [PubMed]
- Klintsova, A.Y.; Helfer, J.L.; Calizo, L.H.; Dong, W.K.; Goodlett, C.R.; Greenough, W.T. Persistent impairment of hippocampal neurogenesis in young adult rats following early postnatal alcohol exposure. Alcohol. Clin. Exp. Res. 2007, 31, 2073–2082. [Google Scholar] [CrossRef] [PubMed]
- Ieraci, A.; Herrera, D.G. Single alcohol exposure in early life damages hippocampal stem/progenitor cells and reduces adult neurogenesis. Neurobiol. Dis. 2007, 26, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, D.F.; Hartman, R.E.; Boyle, M.P.; Vogt, S.K.; Brooks, A.R.; Tenkova, T.; Young, C.; Olney, J.W.; Muglia, L.J. Apoptotic neurodegeneration induced by ethanol in neonatal mice is associated with profound learning/memory deficits in juveniles followed by progressive functional recovery in adults. Neurobiol. Dis. 2004, 17, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Crews, F.T.; Mdzinarishvili, A.; Kim, D.; He, J.; Nixon, K. Neurogenesis in adolescent brain is potently inhibited by ethanol. Neuroscience 2006, 137, 437–445. [Google Scholar] [PubMed]
- Shang, L.; Liu, J.; Zhu, Q.; Zhao, L.; Feng, Y.; Wang, X.; Cao, W.; Xin, H. Gypenosides protect primary cultures of rat cortical cells against oxidative neurotoxicity. Brain Res. 2006, 1102, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.L.; Deng, J.P.; Wang, B.H.; Zhao, Z.W.; Li, J.; Gao, L.; Liu, B.L.; Xong, J.R.; Guo, X.D.; Yan, Z.Q.; et al. Gypenosides improve cognitive impairment induced by chronic cerebral hypoperfusion in rats by suppressing oxidative stress and astrocytic activation. Behav. Pharmacol. 2011, 22, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Niu, L.; Guo, X.D.; Gao, L.; Li, W.X.; Jia, D.; Wang, X.L.; Ma, L.T.; Gao, G.D. Gypenosides protects dopaminergic neurons in primary culture against MPP(+)-induced oxidative injury. Brain Res. Bull. 2010, 83, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Park, M.S.; Kim, S.H.; Hwang, B.Y.; Lee, C.K.; Lee, M.K. Neuroprotective effects of herbal ethanol extracts from Gynostemma pentaphyllum in the 6-hydroxydopamine-lesioned rat model of Parkinson’s disease. Molecules 2010, 15, 2814–2824. [Google Scholar] [CrossRef]
- Wang, P.; Niu, L.; Gao, L.; Li, W.X.; Jia, D.; Wang, X.L.; Gao, G.D. Neuroprotective effect of gypenosides against oxidative injury in the substantia nigra of a mouse model of Parkinson’s disease. J. Int. Med. Res. 2010, 38, 1084–1092. [Google Scholar] [CrossRef] [PubMed]
- Lois, C.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Chain migration of neuronal precursors. Science 1996, 271, 978–981. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Reeves, A.J.; Graziano, M.S.; Gross, C.G. Neurogenesis in the neocortex of adult primates. Science 1999, 286, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Kubbutat, M.H.; Key, G.; Duchrow, M.; Schluter, C.; Flad, H.D.; Gerdes, J. Epitope analysis of antibodies recognising the cell proliferation associated nuclear antigen previously defined by the antibody Ki-67 (Ki-67 protein). J. Clin. Pathol. 1994, 47, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.P.; Couillard-Despres, S.; Cooper-Kuhn, C.M.; Winkler, J.; Aigner, L.; Kuhn, H.G. Transient expression of doublecortin during adult neurogenesis. J. Comp. Neurol. 2003, 467, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bignami, A.; Dahl, D. Astrocyte-specific protein and neuroglial differentiation. An immunofluorescence study with antibodies to the glial fibrillary acidic protein. J. Comp. Neurol. 1974, 153, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Hansson, A.C.; Nixon, K.; Rimondini, R.; Damadzic, R.; Sommer, W.H.; Eskay, R.; Crews, F.T.; Heilig, M. Long-term suppression of forebrain neurogenesis and loss of neuronal progenitor cells following prolonged alcohol dependence in rats. Int. J. Neuropsychopharmacol. 2010, 13, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.W. Effects of prenatal exposure to ethanol on neocortical development: II. Cell proliferation in the ventricular and subventricular zones of the rat. J. Comp. Neurol. 1989, 287, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.W.; Nowakowski, R.S. Effect of prenatal exposure to ethanol on the cell cycle kinetics and growth fraction in the proliferative zones of fetal rat cerebral cortex. Alcohol. Clin. Exp. Res. 1991, 15, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Doetsch, F.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Cellular composition and three-dimensional organization of the subventricular germinal zone in the adult mammalian brain. J. Neurosci. 1997, 17, 5046–5061. [Google Scholar] [PubMed]
- Romanko, M.J.; Rola, R.; Fike, J.R.; Szele, F.G.; Dizon, M.L.; Felling, R.J.; Brazel, C.Y.; Levison, S.W. Roles of the mammalian subventricular zone in cell replacement after brain injury. Prog. Neurobiol. 2004, 74, 77–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Sun, T.; Kong, L.; Shang, Z.H.; Yang, K.Q.; Zhang, Q.Y.; Jing, F.M.; Dong, L.; Xu, X.F.; Liu, J.X.; et al. Gypenosides pre-treatment protects the brain against cerebral ischemia and increases neural stem cells/progenitors in the subventricular zone. Int. J. Dev. Neurosci. 2014, 33, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhao, Z.; Gao, L.; Deng, J.; Wang, B.; Xu, D.; Liu, B.; Qu, Y.; Yu, J.; Li, J.; et al. Gypenoside attenuates white matter lesions induced by chronic cerebral hypoperfusion in rats. Pharmacol. Biochem. Behav. 2011, 99, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Kakita, A.; Goldman, J.E. Patterns and dynamics of SVZ cell migration in the postnatal forebrain: Monitoring living progenitors in slice preparations. Neuron 1999, 23, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Luskin, M.B. Restricted proliferation and migration of postnatally generated neurons derived from the forebrain subventricular zone. Neuron 1993, 11, 173–189. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Verdugo, J.M.; Doetsch, F.; Wichterle, H.; Lim, D.A.; Alvarez-Buylla, A. Architecture and cell types of the adult subventricular zone: In search of the stem cells. J. Neurobiol. 1998, 36, 234–248. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, L.; Yang, K.-Q.; Fu, W.-Y.; Shang, Z.-H.; Zhang, Q.-Y.; Jing, F.-M.; Li, L.-L.; Xin, H.; Wang, X.-J. Gypenosides Protected the Neural Stem Cells in the Subventricular Zone of Neonatal Rats that Were Prenatally Exposed to Ethanol. Int. J. Mol. Sci. 2014, 15, 21967-21979. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151221967
Dong L, Yang K-Q, Fu W-Y, Shang Z-H, Zhang Q-Y, Jing F-M, Li L-L, Xin H, Wang X-J. Gypenosides Protected the Neural Stem Cells in the Subventricular Zone of Neonatal Rats that Were Prenatally Exposed to Ethanol. International Journal of Molecular Sciences. 2014; 15(12):21967-21979. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151221967
Chicago/Turabian StyleDong, Lun, Kun-Qi Yang, Wen-Yan Fu, Zhen-Hua Shang, Qing-Yu Zhang, Fang-Miao Jing, Lin-Lin Li, Hua Xin, and Xiao-Jing Wang. 2014. "Gypenosides Protected the Neural Stem Cells in the Subventricular Zone of Neonatal Rats that Were Prenatally Exposed to Ethanol" International Journal of Molecular Sciences 15, no. 12: 21967-21979. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151221967