Effect of Wnt3a on Keratinocytes Utilizing in Vitro and Bioinformatics Analysis


 ,
,
Abstract
:1. Introduction
2. Results
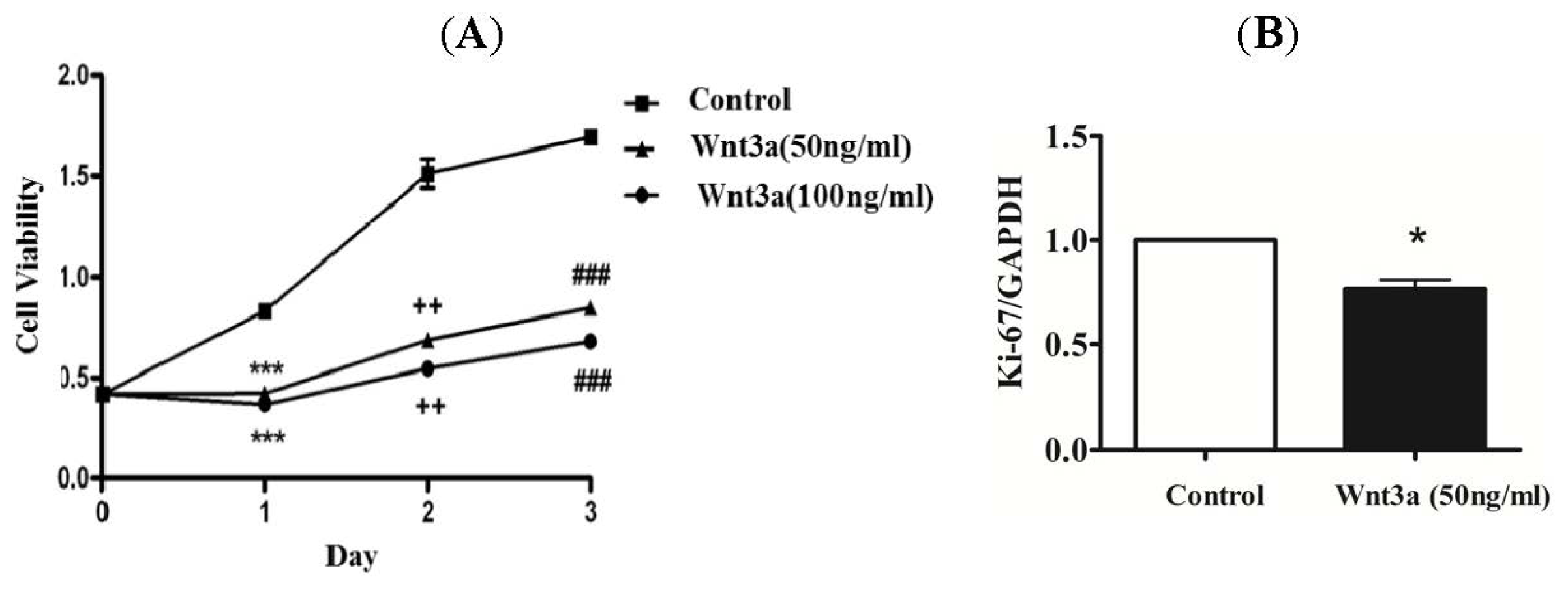
2.1. Effect of Wnt3a on the Proliferation of Cultured Keratinocytes
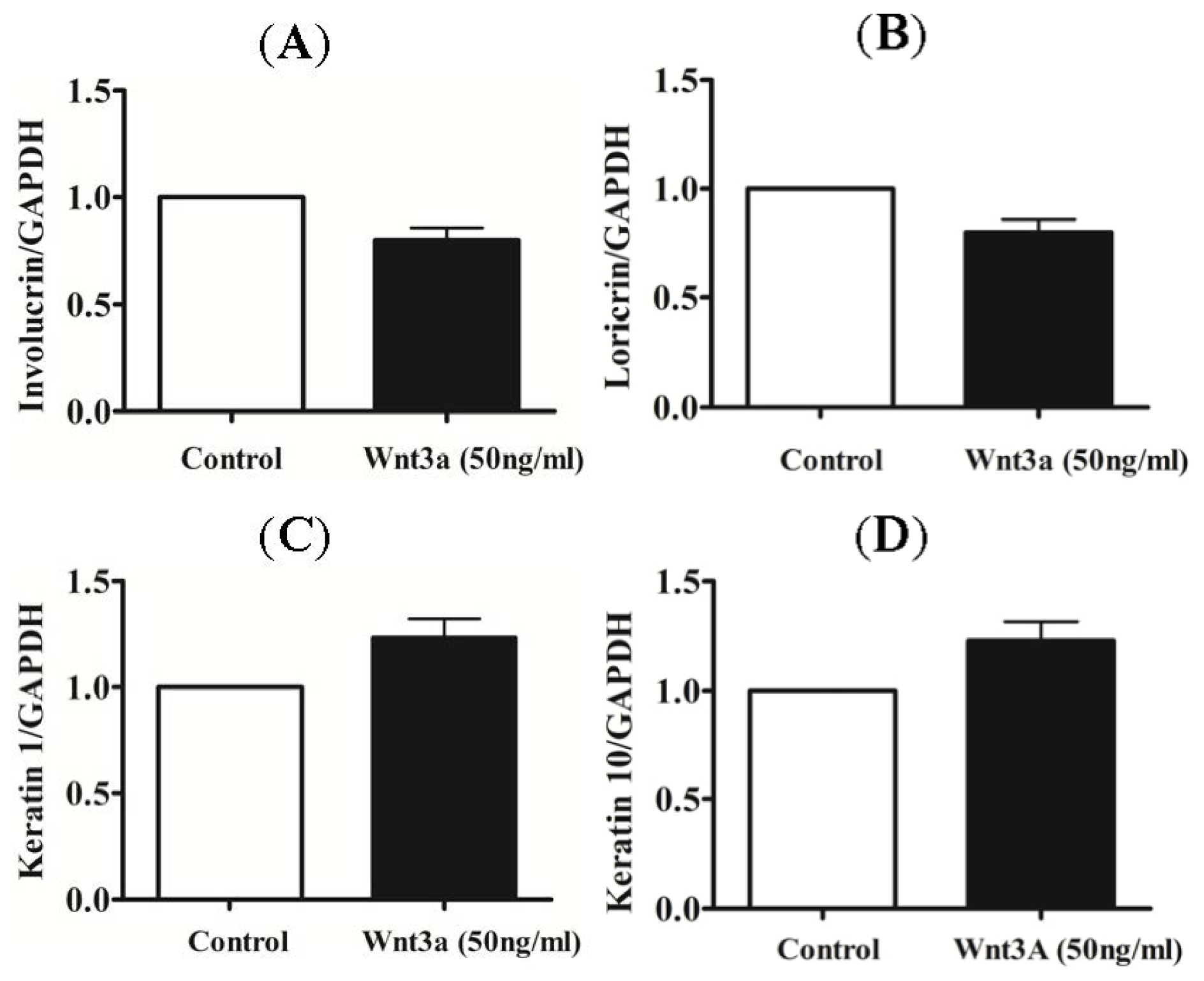
2.2. Effect of Exogenous Wnt3a on the Differentiation of Cultured Keratinocytes
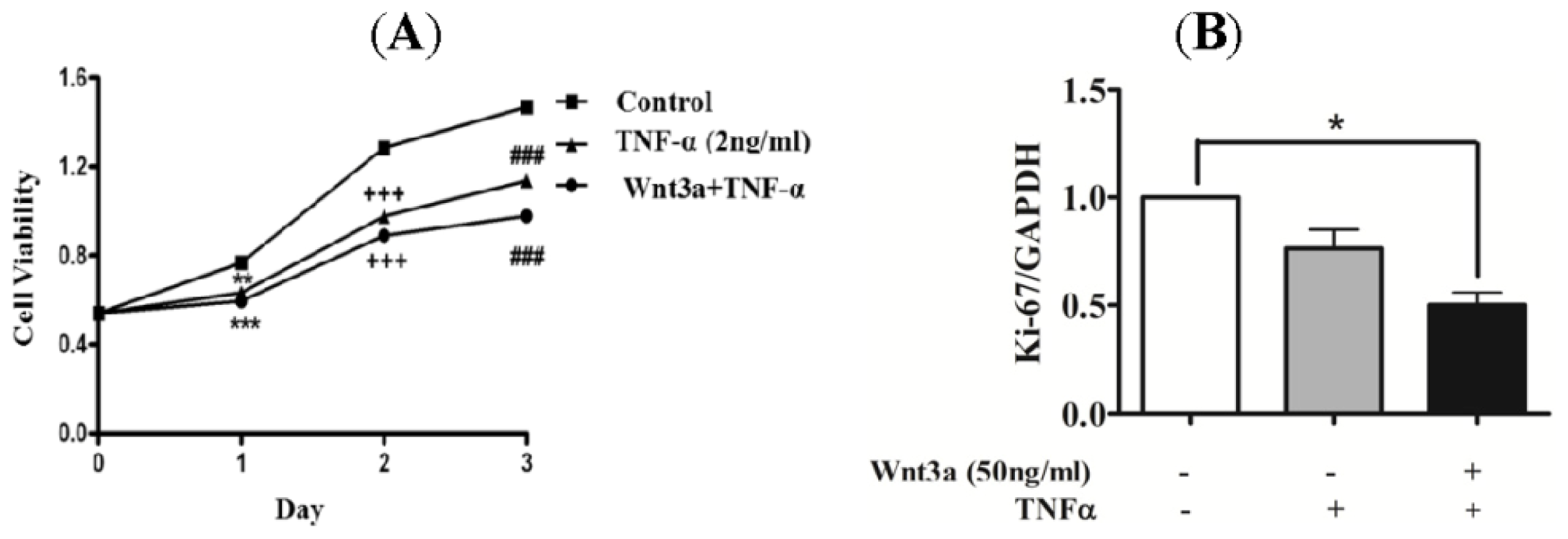
2.3. Effect of Wnt3a on Proliferation of TNFprocess of keratinocytes.a
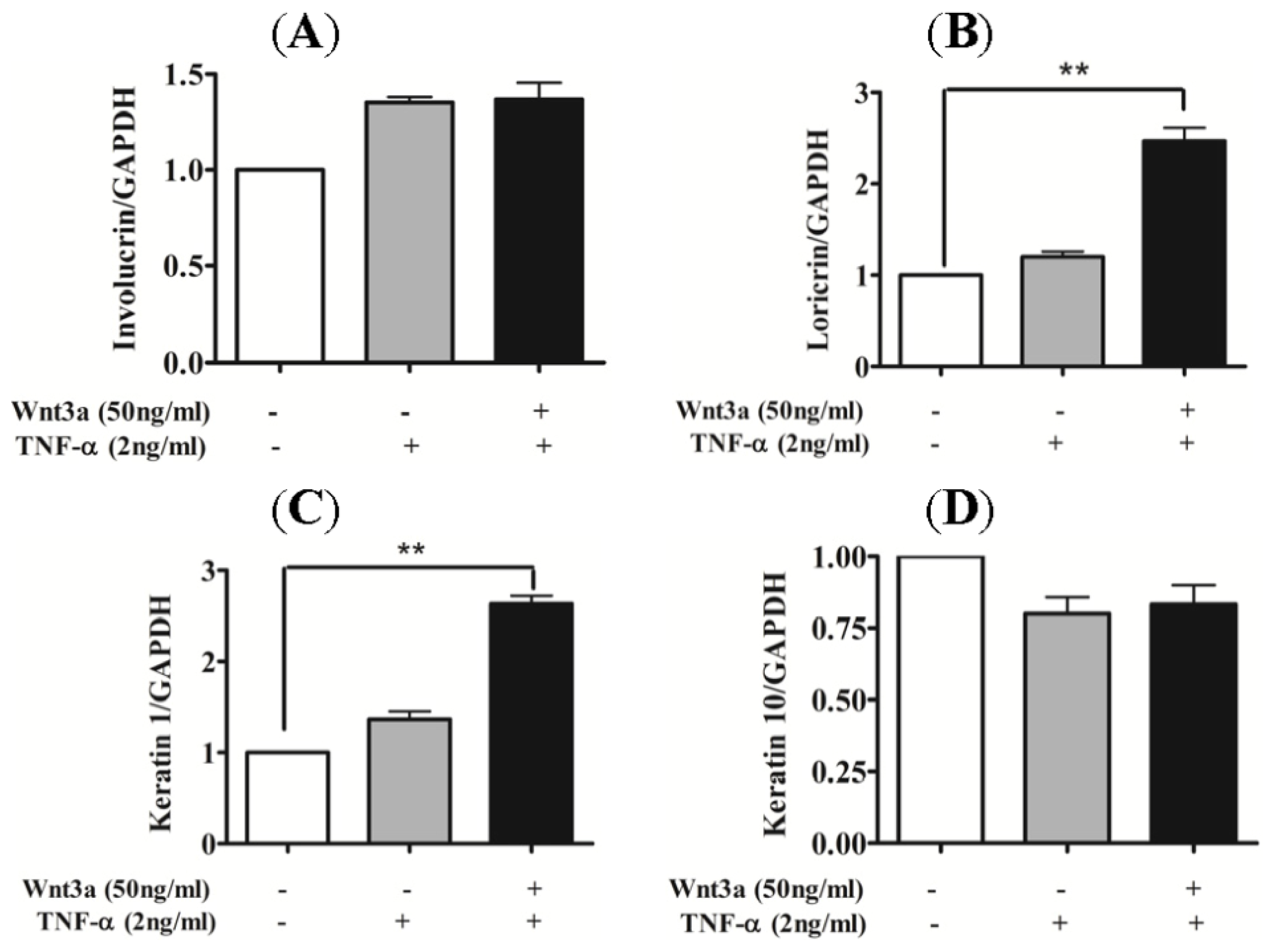
2.4. Effect of Wnt3a on Differentiation of TNFα-Stimulated Keratinocytes
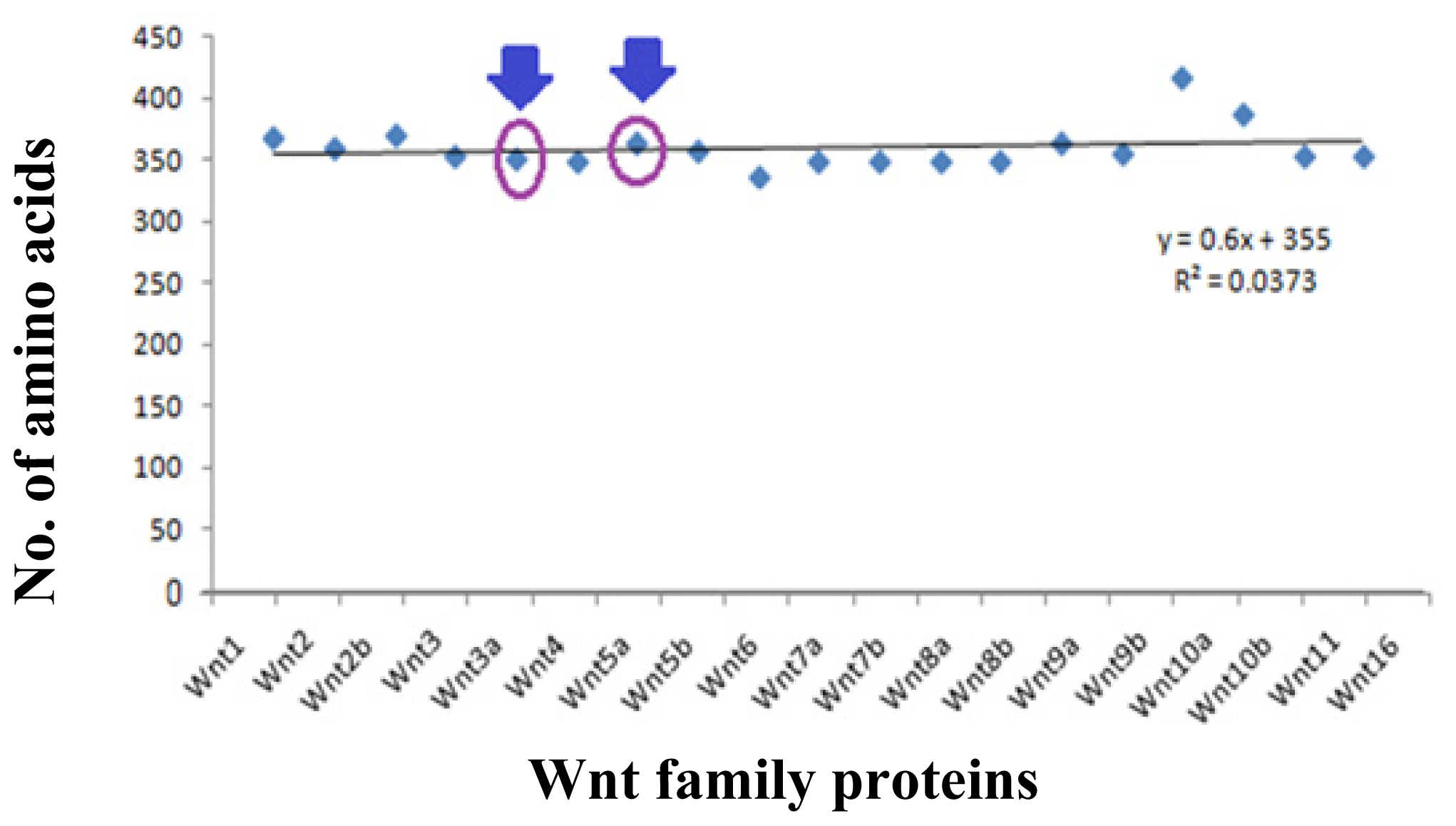
2.5. Data Collection of Wnt Family Proteins for Computational Analysis
2.6. Multiple Sequences Alignment (MSA) of Wnt Proteins
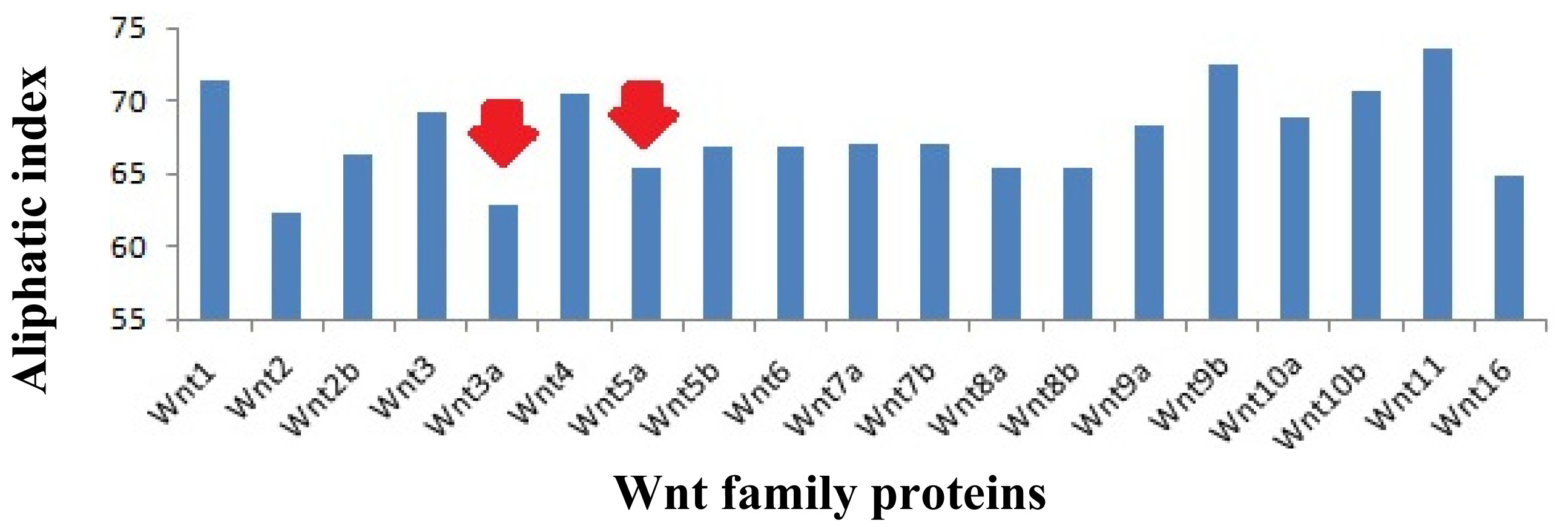
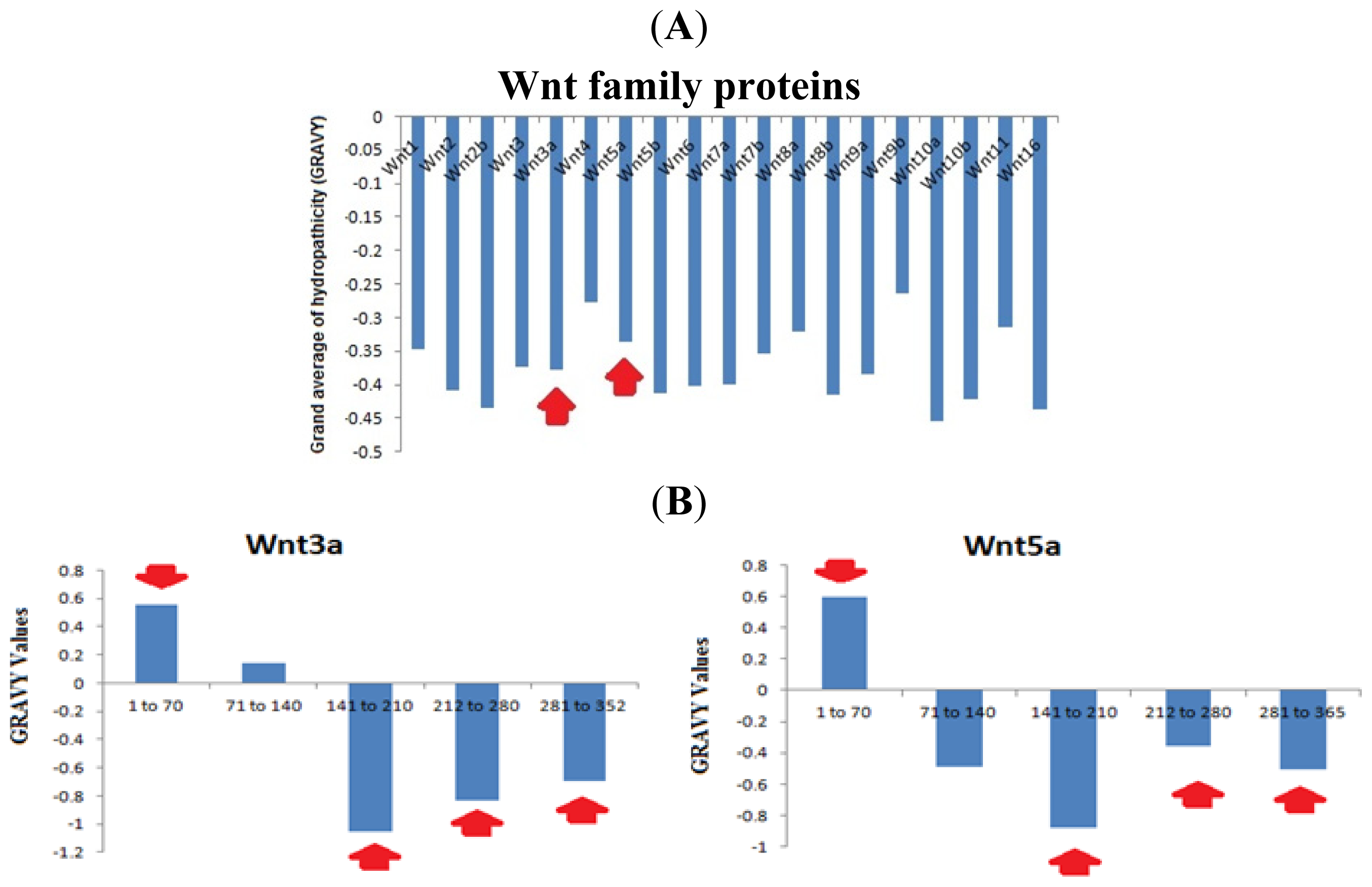
2.7. Analysis of the Aliphatic Index and GRAVY of the Wnt Family Proteins
2.8. Molecular Evolution Analysis of the Wnt Family Proteins
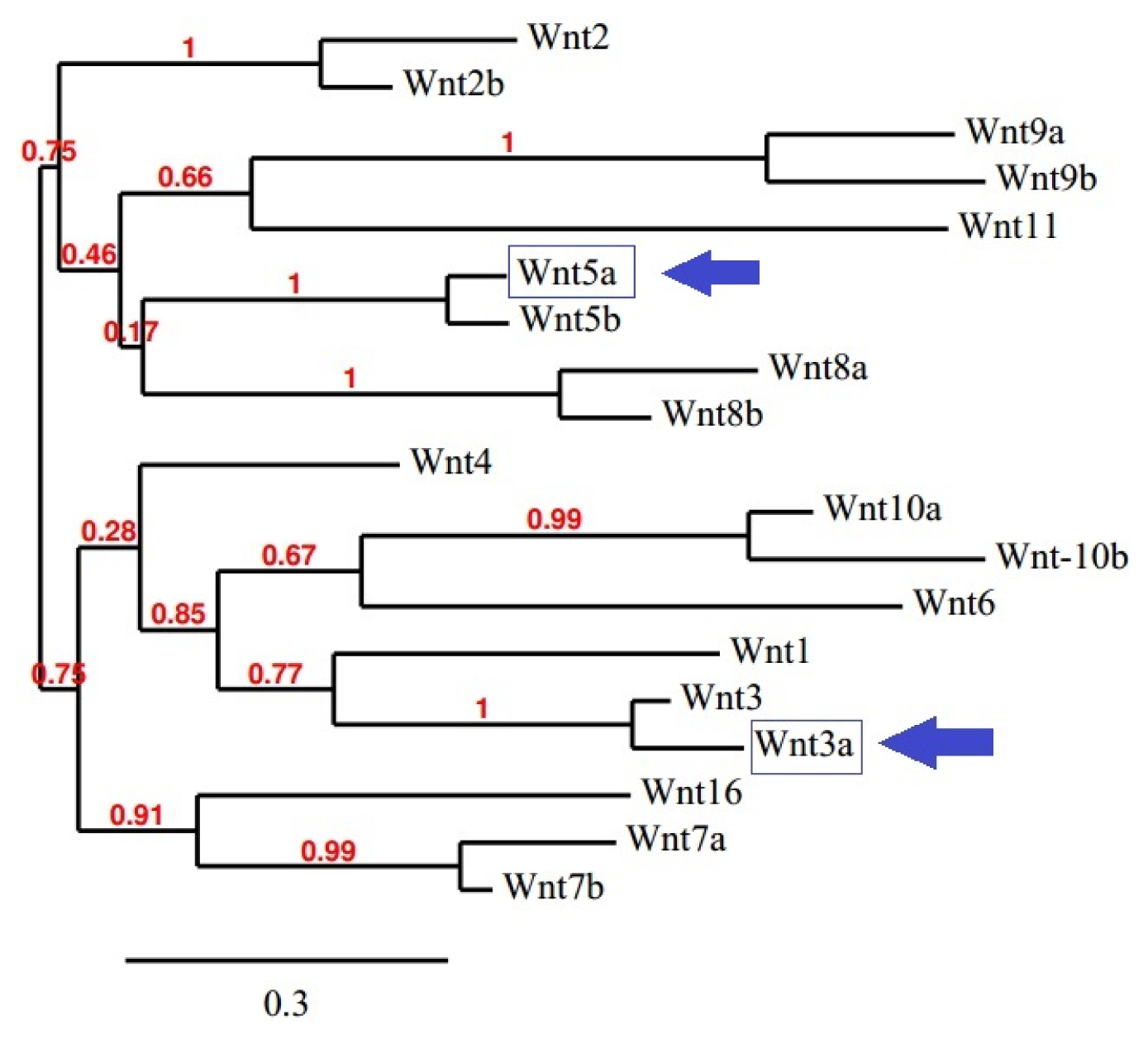
2.9. Creation of the Phylogenetic Tree for the Wnt Family Seed Alignment
2.10. Conservation Assessment of Wnt Family Proteins in Different Species
2.11. Prediction of Precise Version of Sequence Similarity and Generation of HMM Logos of Wnt Family Proteins
2.12. Understanding the Protein–Protein Interaction Network of Wnt Family Proteins
3. Discussion
4. Methods
4.1. Tissue Specimens
4.2. Cell Culture
4.3. Cell Viability Assay (MTT Assay)
4.4. Real Time PCR
4.5. Data Collection of Wnt Family Proteins for Computational Analysis
4.6. Multiple Sequences Alignment (MSA) of Wnt Proteins
4.7. Analysis of Aliphatic Index and GRAVY of Wnt Family Proteins
4.8. Molecular Evolution Analysis of Wnt Family Proteins
4.9. Creation of the Phylogenetic Tree for Wnt Family Seed Alignment
4.10. Conservation Assessment of Wnt Family Proteins in Different Species
4.11. Prediction of Precise Version of Sequence Similarity through Sequence Logo and Generation of HMM Logos of Wnt Family Proteins
4.12. Understanding the Protein–Protein Interaction Network of Wnt Family Proteins
4.13. Statistical Analysis
5. Conclusions
Supplementary Information
ijms-15-05472-s001.pdfAcknowledgments
Conflicts of Interest
- Author ContributionsDong-Keun Song, Sang-Soo Lee and Myung-Sun Hong participated in designing the study. Ashish Ranjan Sharma, Chiranjib Chakraborty and Ju-Suk Nam conducted the study. Data was collected by Kee-Jeong Bae and Young Her and analyzed by Sang-Soo Lee, Ju-Suk Nam and Garima Sharma. Manuscript was written by Ashish Ranjan Sharma, George Priya Doss and Chiranjib Chakraborty.
References
- Kikuchi, A.; Yamamoto, H.; Sato, A. Selective activation mechanisms of Wnt signaling pathways. Trends Cell Biol 2009, 19, 119–129. [Google Scholar]
- Sen, M.; Ghosh, G. Transcriptional outcome of Wnt-Frizzled signal transduction in inflammation: Evolving concepts. J. Immunol 2008, 181, 4441–4445. [Google Scholar]
- Amerongen, R.V.; Mikels, A.; Nusse, R. Alternative Wnt signaling is initiated by distinct receptors. Sci. Signal 2008, 1, re9. [Google Scholar]
- Wehrli, M.; Dougan, S.T.; Caldwell, K.; O’Keefe, L.; Schwartz, S.; Vaizel-Ohayon, D.; Schejter, E.; Tomlinson, A.; DiNardo, S. Arrow encodes an LDL-receptor-related protein essential for wingless signalling. Nature 2000, 407, 527–530. [Google Scholar]
- Verkaar, F.; Zaman, G.J.A. Model for signaling specificity of Wnt/Frizzled combinations through co-receptor recruitment. FEBS Lett 2010, 584, 3850–3854. [Google Scholar]
- Liu, G.; Bafico, A.; Aaronson, S.A. The mechanism of endogenous receptor activation functionally distinguishes prototype canonical and noncanonical Wnts. Mol. Cell Biol 2005, 25, 3475–3482. [Google Scholar]
- Tao, Q.; Yokota, C.; Puck, H.; Kofron, M.; Birsoy, B.; Yan, D.; Asashima, M.; Wylie, C.C.; Lin, X.; Heasman, J. Maternal Wnt11 activates the canonical Wnt signaling pathway required for axis formation in Xenopus embryos. Cell 2005, 120, 857–871. [Google Scholar]
- Kawano, Y.; Kypta, R. Secreted antagonists of the Wnt signalling pathway. J. Cell Sci 2003, 116, 2627–2634. [Google Scholar]
- Logan, C.Y.; Nusse, R. The Wnt signaling pathway in development and disease. Annu. Rev. Cell Dev. Biol 2004, 20, 781–810. [Google Scholar]
- Hsieh, J.C.; Kodjabachian, L.; Rebbert, M.L.; Rattner, A.; Smallwood, P.M.; Samos, C.H.; Nusse, R.; Dawid, I.B.; Nathans, J.A. New secreted protein that binds to Wnt proteins and inhibits their activities. Nature 1999, 398, 431–436. [Google Scholar]
- Perera, G.K.; Di Meglio, P.; Nestle, F.O. Psoriasis. Annu. Rev. Pathol 2012, 7, 385–422. [Google Scholar]
- Schon, M.P.; Boehncke, W.H. Psoriasis. N. Engl. J. Med 2005, 52, 1899–1912. [Google Scholar]
- Lebwohl, M. Psoriasis. Lancet 2003, 361, 1197–1204. [Google Scholar]
- Weinstein, G.D.; McCullough, J.L.; Ross, P.A. Cell kinetic basis for pathophysiology of psoriasis. J. Investig. Dermatol 1985, 85, 579–583. [Google Scholar]
- Gudjonsson, J.E.; Johnston, A.; Stoll, S.W.; Riblett, M.B.; Xing, X.; Kochkodan, J.J.; Ding, J.; Nair, R.P.; Aphale, A.; Voorhees, J.J.; et al. Evidence for altered Wnt signaling in psoriatic skin. J. Investig. Dermatol 2010, 130, 1849–1859. [Google Scholar]
- Teh, M.T.; Blaydon, D.; Ghali, L.R.; Briggs, V.; Edmunds, S.; Pantazi, E.; Barnes, M.R.; Leigh, I.M.; Kelsell, D.P.; Philpott, M.P. Role for Wnt16B in human epidermal keratinocyte proliferation and differentiation. J. Cell Sci 2007, 120, 330–339. [Google Scholar]
- Scholzen, T.; Gerdes, J. The Ki-67 protein: From the known and the unknown. J. Cell Physiol 2000, 182, 311–322. [Google Scholar]
- Gordon, K.B.; Langley, R.G.; Leonardi, C.; Toth, D.; Menter, M.A.; Kang, S.; Heffernan, M.; Miller, B.; Hamlin, R.; Lim, L. Clinical response to adalimumab treatment in patients with moderate to severe psoriasis: Double-blind, randomized controlled trial and open-label extension study. J. Am. Acad. Dermatol 2006, 55, 598–606. [Google Scholar]
- Reich, K.; Nestle, F.O.; Papp, K.; Ortonne, J.P.; Evans, R.; Guzzo, C.; Li, S.; Dooley, L.T.; Griffiths, C.E. EXPRESS study investigators. Infliximab induction and maintenance therapy for moderate-to-severe psoriasis: A phase III, multicentre, double-blind trial. Lancet 2005, 366, 1367–1374. [Google Scholar]
- Ikai, A. Thermostability and aliphatic index of globular proteins. J. Biochem 1980, 88, 1895–1898. [Google Scholar]
- Cassano, N.; Vestita, M.; Apruzzi, D.; Vena, G.A. Alcohol, psoriasis, liver disease, and anti-psoriasis drugs. Int. J. Dermatol 2011, 50, 1323–1331. [Google Scholar]
- Chamcheu, J.C.; Siddiqui, I.A.; Syed, D.N.; Adhami, V.M.; Liovic, M.; Mukhtar, H. Keratin gene mutations in disorders of human skin and its appendages. Arch. Biochem. Biophys 2011, 508, 123–137. [Google Scholar]
- Guttman-Yassky, E.; Nograles, K.E.; Krueger, J.G. Contrasting pathogenesis of atopic dermatitis and psoriasis—Part I: Clinical and pathologic concepts. J. Allergy Clin. Immunol 2011, 127, 1110–1118. [Google Scholar]
- Chua, A.W.; Ma, D.; Gan, S.U.; Fu, Z.; Han, H.C.; Song, C.; Sabapathy, K.; Phan, T.T. The role of R-spondin2 in keratinocyte proliferation and epidermal thickening in keloid scarring. J. Investig. Dermatol 2011, 131, 644–654. [Google Scholar]
- Romanowska, M.; Evans, A.; Kellock, D.; Bray, S.E.; McLean, K.; Donandt, S.; Foerster, J. Wnt5a exhibits layer-specific expression in adult skin, is upregulated in psoriasis, and synergizes with type 1 interferon. PLoS One 2009, 4, e5354. [Google Scholar]
- Yamaguchi, Y.; Passeron, T.; Hoashi, T.; Watabe, H.; Rouzaud, F.; Yasumoto, K.; Hara, T.; Tohyama, C.; Katayama, I.; Miki, T.; et al. Dickkopf 1 (DKK1) regulates skin pigmentation and thickness by affecting Wnt/beta-catenin signaling in keratinocytes. FASEB J 2008, 22, 1009–1020. [Google Scholar]
- Reischl, J.; Schwenke, S.; Beekman, J.M.; Mrowietz, U.; Sturzebecher, S.; Heubach, J.F. Increased expression of Wnt5a in psoriatic plaques. J. Investig. Dermatol 2007, 127, 163–169. [Google Scholar]
- Chiricozzi, A.; Guttman-Yassky, E.; Suarez-Farinas, M.; Nograles, K.E.; Tian, S.; Cardinale, I.; Chimenti, S.; Krueger, J.G. Integrative responses to IL-17 and TNF-α in human keratinocytes account for key inflammatory pathogenic circuits in psoriasis. J. Investig. Dermatol 2011, 131, 677–687. [Google Scholar]
- Fujiwara, S.; Nagai, H.; Oniki, S.; Yoshimoto, T.; Nishigori, C. Interleukin (IL)-17 vs. IL-27: opposite effects on tumor necrosis factor-α-mediated chemokine production in human keratinocytes. Exp. Dermatol 2012, 21, 70–72. [Google Scholar]
- Kumari, S.; Bonnet, M.C.; Ulvmar, M.H.; Wolk, K.; Karagianni, N.; Witte, E.; Uthoff-Hachenberg, C.; Renauld, J.C.; Kollias, G.; Toftgard, R.; et al. Tumor necrosis factor receptor signaling in keratinocytes triggers interleukin-24-dependent psoriasis-like skin inflammation in mice. Immunity 2013, 39, 899–911. [Google Scholar]
- Nusse, R.; Varmus, H. Three decades of Wnts: A personal perspective on how a scientific field developed. EMBO J 2012, 31, 2670–2684. [Google Scholar]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/β-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar]
- Cadigan, K.M.; Nusse, R. Wnt signaling: A common theme in animal development. Genes Dev 1997, 11, 3286–3305. [Google Scholar]
- Kumar, M.; Ahmad, S.; Ahmad, E.; Saifi, M.A.; Khan, R.H. In silico prediction and analysis of Caenorhabditis EF-hand containing proteins. PLoS One 2012, 7, e36770. [Google Scholar]
- Miller, J. The Wnts. Genome Biol 2002, 3, 1–15. [Google Scholar]
- Ain, Q.U.; Seemab, U.; Rashid, S.; Nawaz, M.S.; Kamal, M.A. Prediction of structure of human Wnt-CRD (FZD) complex for computational drug repurposing. PLoS One 2013, 8, e54630. [Google Scholar]
- Finch, P.W.; He, X.; Kelley, M.J.; Uren, A.; Schaudies, R.P.; Popescu, N.C.; Rudikoff, S.; Aaronson, S.A.; Varmus, H.E.; Rubin, J.S. Purification and molecular cloning of a secreted, Frizzled-related antagonist of Wnt action. Proc. Natl. Acad. Sci. USA 1997, 94, 6770–6775. [Google Scholar]
- Hansbrough, J.F. U.S. Human Keratinocytes Supported on a Hydrophilic Membrane and Methods of Using Same to Effect Wound Closure. U.S. Patent 5,693,332, 2 December 1997. [Google Scholar]
- Yamato, M.; Utsumi, M.; Kushida, A.; Konno, C.; Kikuchi, A.; Okano, T. Thermo-responsive culture dishes allow the intact harvest of multilayered keratinocyte sheets without dispase by reducing temperature. Tissue Eng 2001, 7, 473–480. [Google Scholar]
- Hanakam, F.; Gerisch, G.; Lotz, S.; Alt, T.; Seelig, A. Binding of hisactophilin I and II to lipid membranes is controlled by a pH-dependent myristoyl-histidine switch. Biochemistry 1996, 35, 11036–11044. [Google Scholar]
- Gonnet, F.; Lemaitre, G.; Waksman, G.; Tortajada, J. MALDI/MS peptide mass fingerprinting for proteome analysis: Identification of hydrophobic proteins attached to eucaryote keratinocyte cytoplasmic membrane using different matrices in concert. Proteome Sci 2003, 1, 2. [Google Scholar]
- Van Oss, C.J. Long-range and short-range mechanisms of hydrophobic attraction and hydrophilic repulsion in specific and aspecific interactions. J. Mol. Recognit 2003, 16, 177–190. [Google Scholar]
- Kundu, S.K.; Gupta, S.; Stellbrink, J.; Willner, L.; Richter, D. Relating structure and flow of soft colloids. Eur. Phys. J. Spec. Top 2013, 222, 2757–2772. [Google Scholar]
- Sayers, E.W.; Barrett, T.; Benson, D.A.; Bolton, E.; Bryant, S.H.; Canese, K.; Chetvernin, V.; Church, D.M.; DiCuccio, M.; Federhen, S.; et al. Database resources of the national center for biotechnology information. Nucleic Acids Res 2011, 39, D38–D51. [Google Scholar]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol 2011, 7, 539. [Google Scholar]
- Eddy, S.R. Profile hidden Markov models. Bioinformatics 1998, 14, 755–763. [Google Scholar]
- Eddy, S.R. What is a hidden Markov model? Nat. Biotechnol 2004, 22, 1315–1316. [Google Scholar]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res 2003, 31, 3784–3788. [Google Scholar]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res 2008, 36, W465–W469. [Google Scholar]
- Punta, M.; Coggill, P.C.; Eberhardt, R.Y.; Mistry, J.; Tate, J.; Boursnell, C.; Pang, N.; Forslund, K.; Ceric, G.; Clements, J.; et al. The Pfam protein families database. Nucleic Acids Res 2012, 40, D290–D301. [Google Scholar]
- Jensen, L.J.; Kuhn, M.; Stark, M.; Chaffron, S.; Creevey, C.; Muller, J.; Doerks, T.; Julien, P.; Roth, A.; Simonovic, M.; et al. STRING 8—A global view on proteins and their functional interactions in 630 organisms. Nucleic Acids Res 2009, 37, D412–D416. [Google Scholar]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG database: An updated version includes eukaryotes. BMC Bioinform 2003, 4, 41. [Google Scholar]
- Mering, C.V.; Jensen, L.J.; Snel, B.; Hooper, S.D.; Krupp, M.; Foglierini, M.; Jouffre, N.; Huynen, M.A.; Bork, P. STRING: known and predicted protein–protein associations, integrated and transferred across organisms. Nucleic Acids Res 2005, 33, D433–D437. [Google Scholar]
- Smith, T.F.; Waterman, M.S. Identification of common molecular subsequences. J. Mol. Biol 1981, 47, 195–197. [Google Scholar]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res 2004, 14, 1188–1190. [Google Scholar]
- Schneider, T.D.; Stephens, R.M. Sequence logos a new way to display consensus sequences. Nucleic Acids Res 1990, 18, 6097–6100. [Google Scholar]
- Schuster-Bocklerm, B.; Schultz, J.; Rahmann, S. HMM Logos for visualization of protein families. BMC Bioinform 2004, 5, 7. [Google Scholar]





 ; R,K =
; R,K =
 ; N,Q,S,T =
; N,Q,S,T =
 ; C =
(Threshhold + 60%); C =
; C =
(Threshhold + 60%); C =
 (Threshhold + 100%); E,D =
(Threshhold + 100%); E,D =
 ; G =
; G =
 ; H,Y =
; H,Y =
 ; P =
; P =
 .
; R,K =
; N,Q,S,T =
; C =
(Threshhold + 60%); C =
(Threshhold + 100%); E,D =
; G =
; H,Y =
; P =
.
.
; R,K =
; N,Q,S,T =
; C =
(Threshhold + 60%); C =
(Threshhold + 100%); E,D =
; G =
; H,Y =
; P =
.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Forward primer (5′–3′) | Reverse primer (3′–5′) | GenBank No. |
|---|---|---|---|
| Loricrin | TAGAGCTCTCATGATGCTACCCGA | GGGTTGGGAGGTAGTTGTACAGAA | NM_000427.2 |
| Involucrin | AATGAAACAGCCAACTCCACTGCC | AGGCAGTCCCTTTACAGCAGTCAT | NM_005517.2 |
| Keratin1 | GGCAGTTCCAGCGTGAAGTTTGTT | TTCTCCGGTAAGGCTGGGACAAAT | NM_006121.3 |
| Keratin10 | TTGGTGGAGGTAGCTTTCGTGGAA | AGAAGGCCACCATCTCCTCCAAAT | NM_000421.3 |
| Ki-67 | TGACAAGCCCACGACTGATGAGAA | CTTTGCCTGCTGATGGTGTTCGTT | NM_002417.4 |
| GAPDH | TCGACAGTCAGCCGCATCTTCTTT | ACCAAATCCGTTGACTCCGACCTT | NM_002046.3 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nam, J.-S.; Chakraborty, C.; Sharma, A.R.; Her, Y.; Bae, K.-J.; Sharma, G.; Doss, G.P.; Lee, S.-S.; Hong, M.-S.; Song, D.-K. Effect of Wnt3a on Keratinocytes Utilizing in Vitro and Bioinformatics Analysis. Int. J. Mol. Sci. 2014, 15, 5472-5495. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms15045472
Nam J-S, Chakraborty C, Sharma AR, Her Y, Bae K-J, Sharma G, Doss GP, Lee S-S, Hong M-S, Song D-K. Effect of Wnt3a on Keratinocytes Utilizing in Vitro and Bioinformatics Analysis. International Journal of Molecular Sciences. 2014; 15(4):5472-5495. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms15045472
Chicago/Turabian StyleNam, Ju-Suk, Chiranjib Chakraborty, Ashish Ranjan Sharma, Young Her, Kee-Jeong Bae, Garima Sharma, George Priya Doss, Sang-Soo Lee, Myung-Sun Hong, and Dong-Keun Song. 2014. "Effect of Wnt3a on Keratinocytes Utilizing in Vitro and Bioinformatics Analysis" International Journal of Molecular Sciences 15, no. 4: 5472-5495. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms15045472