
Ponicidin Induces Apoptosis via JAK2 and STAT3 Signaling Pathways in Gastric Carcinoma

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Ponicidin Inhibits Proliferation of MKN28 Cells

2.2. Ponicidin Induces Apoptosis of MKN28 Cells
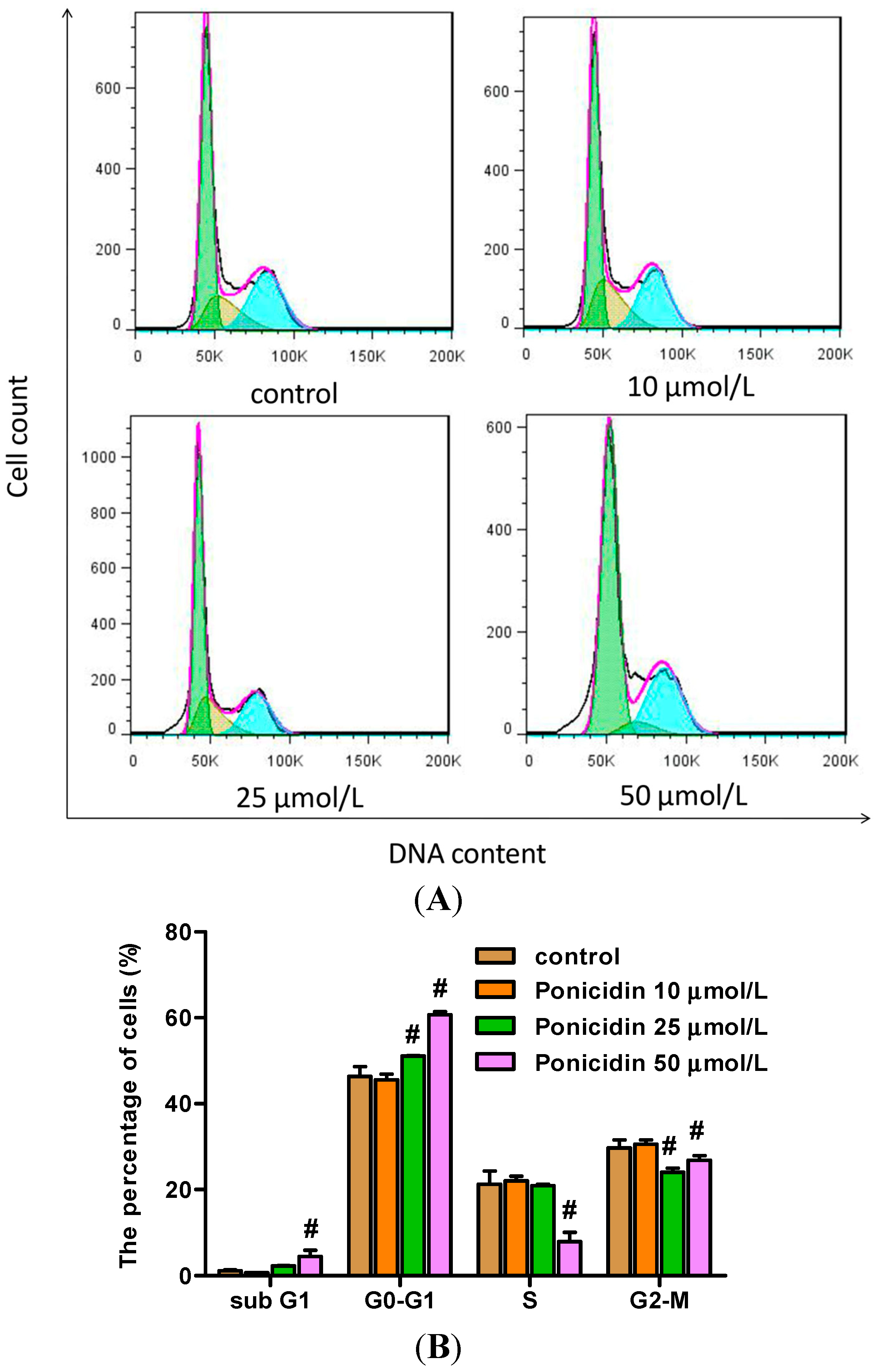
2.3. Ponicidin Arrests the Cell Cycle of MKN28 Cells


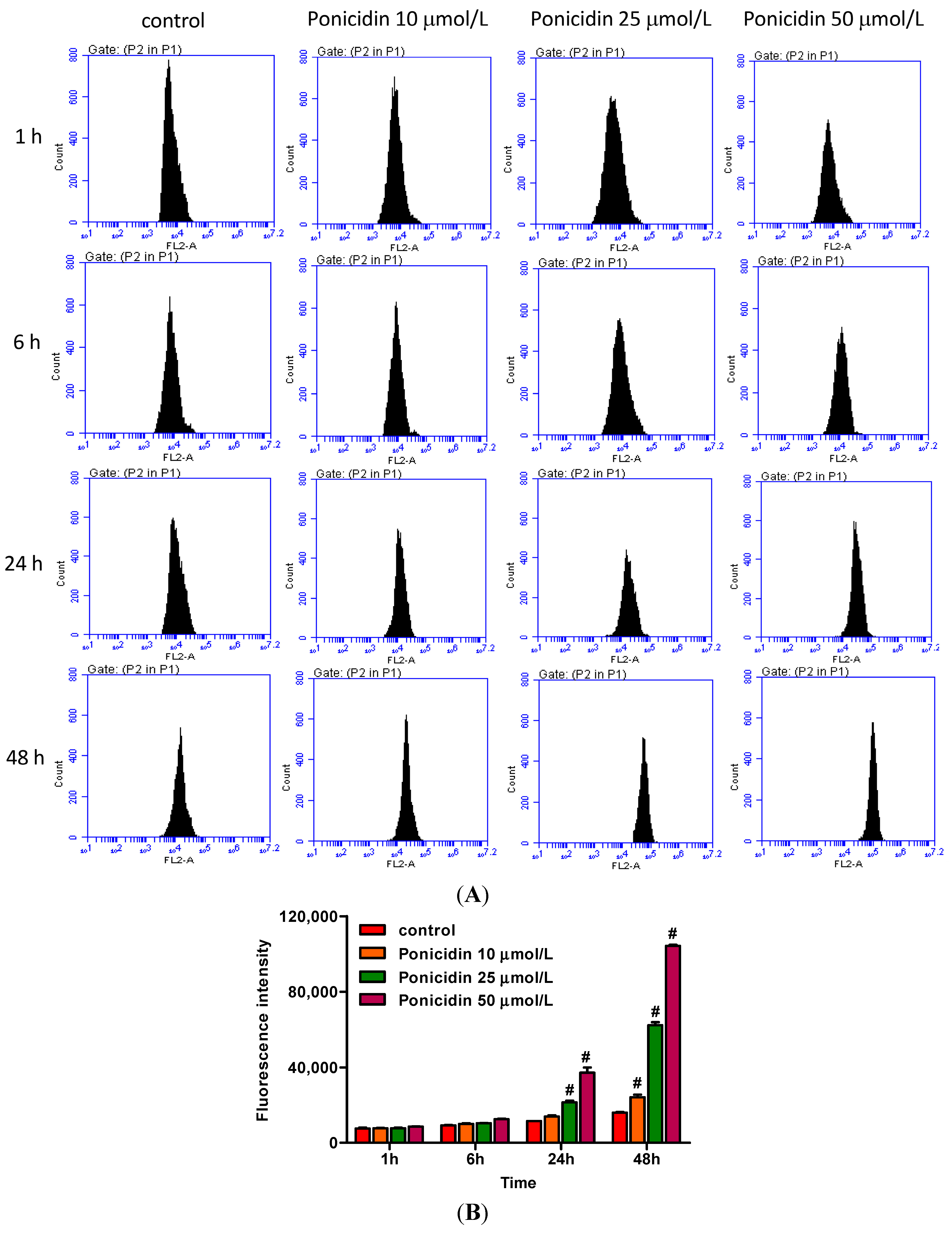
2.4. Ponicidin Induces Reactive Oxygen Species (ROS) Generation in MKN28 Cells

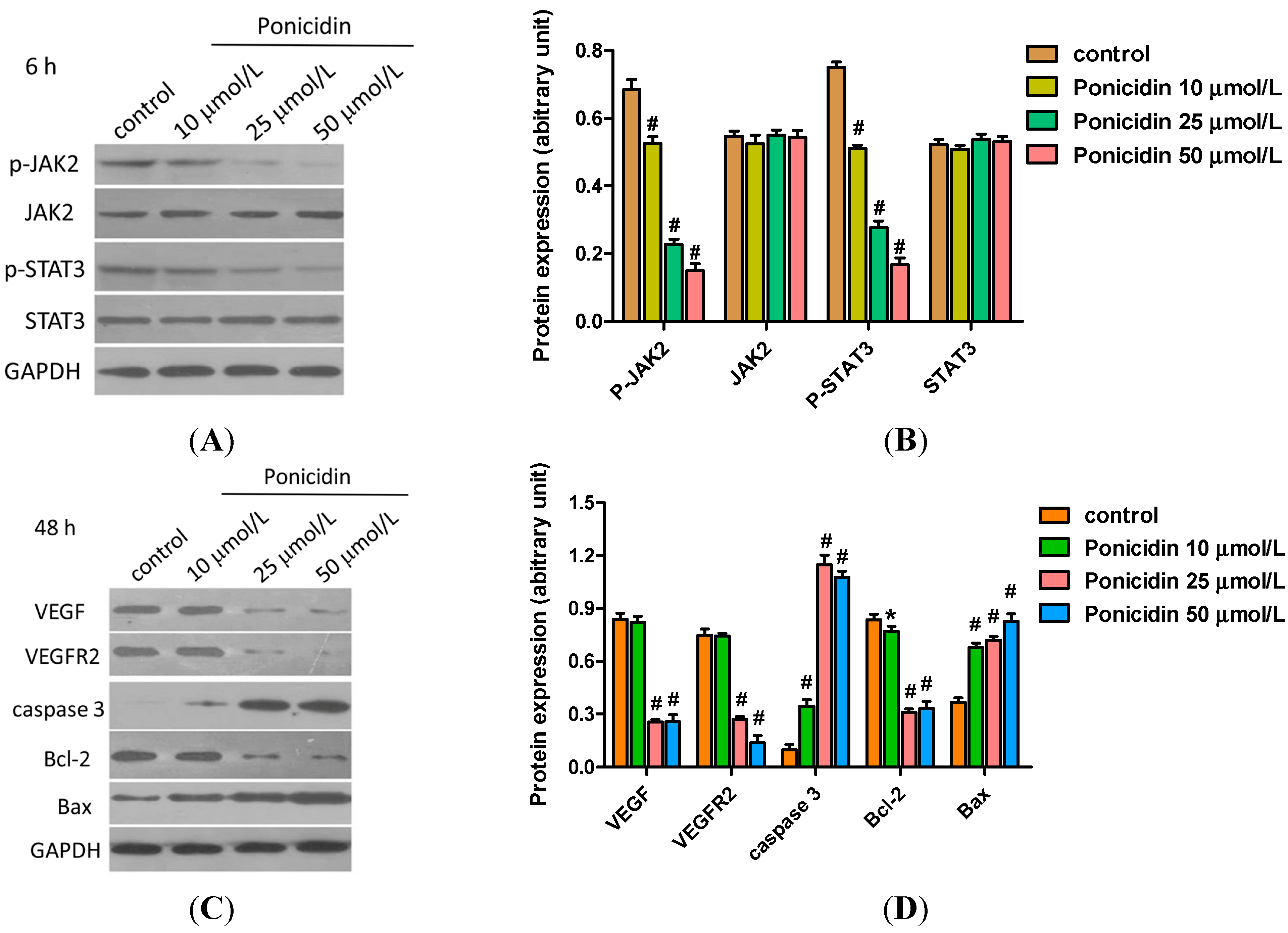
2.5. Ponicidin Effects on Proteins Expression in MKN28 Cells

2.6. Ponicidin Induces Activity of Caspase-3 in MKN28 Cells

3. Experimental Section
3.1. Reagents
3.2. Cell Culture
3.3. Cell Viability Assay
3.4. Flow Cytometry (FCM) Detection
3.5. Western Blot
3.6. Caspase-3 Activation Assay
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Coupland, V.H.; Lagergren, J.; Luchtenborg, M.; Jack, R.H.; Allum, W.; Holmberg, L.; Hanna, G.B.; Pearce, N.; Moller, H. Hospital volume, proportion resected and mortality from oesophageal and gastric cancer: A population-based study in England, 2004–2008. Gut 2013, 62, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Camargo, M.C.; Kim, W.H.; Chiaravalli, A.M.; Kim, K.M.; Corvalan, A.H.; Matsuo, K.; Yu, J.; Sung, J.J.; Herrera-Goepfert, R.; Meneses-Gonzalez, F.; et al. Improved survival of gastric cancer with tumour Epstein-Barr virus positivity: An international pooled analysis. Gut 2014, 63, 236–243. [Google Scholar] [PubMed]
- Shibata, W.; Ariyama, H.; Westphalen, C.B.; Worthley, D.L.; Muthupalani, S.; Asfaha, S.; Dubeykovskaya, Z.; Quante, M.; Fox, J.G.; Wang, T.C. Stromal cell-derived factor-1 overexpression induces gastric dysplasia through expansion of stromal myofibroblasts and epithelial progenitors. Gut 2013, 62, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, X.; Zhang, J.; Chen, Y.; Xu, J.; Xia, X.; He, S.; Qiang, F.; Li, A.; Shu, Y. CHIP functions as a novel suppressor of tumour angiogenesis with prognostic significance in human gastric cancer. Gut 2013, 62, 496–508. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N. Vascular endothelial growth factor and the regulation of angiogenesis. Recent Prog. Horm. Res. 1999, 55, 15–35. [Google Scholar]
- Chen, S.H.; Murphy, D.A.; Lassoued, W.; Thurston, G.; Feldman, M.D.; Lee, W.M. Activated STAT3 is a mediator and biomarker of VEGF endothelial activation. Cancer Biol. Ther. 2008, 7, 1994–2003. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Han, Z. C. STAT3: A critical transcription activator in angiogenesis. Med. Res. Rev. 2008, 28, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Sawamiphak, S.; Seidel, S.; Essmann, C.L.; Wilkinson, G.A.; Pitulescu, M.E.; Acker, T.; Acker-Palmer, A. Ephrin-B2 regulates VEGFR2 function in developmental and tumour angiogenesis. Nature 2010, 465, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Haisma, H.J.; Kamps, G.K.; Bouma, A.; Geel, T.M.; Rots, M.G.; Kariath, A.; Bellu, A.R. Selective targeting of adenovirus to αvβ3 integrins, VEGFR2 and Tie2 endothelial receptors by angio-adenobodies. Int. J. Pharm. 2010, 391, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Choi, S.; Lee, Y.; Lee, H.J.; Kim, K.H.; Ahn, K.S.; Bae, H.; Lee, H.J.; Lee, E.O.; Ahn, K.S.; et al. Herbal compound farnesiferol C exerts antiangiogenic and antitumor activity and targets multiple aspects of VEGFR1 (Flt1) or VEGFR2 (Flk1) signaling cascades. Mol. Cancer Ther. 2010, 9, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.Y.; Sun, D.; Liu, X.Y.; Pan, Y.; Liang, H. STAT3 correlates with lymph node metastasis and cell survival in gastric cancer. World J. Gastroenterol. 2010, 16, 5380–5387. [Google Scholar] [CrossRef] [PubMed]
- Saxena, N.K.; Sharma, D.; Ding, X.; Lin, S.; Marra, F.; Merlin, D.; Anania, F.A. Concomitant activation of the JAK/STAT, PI3K/AKT, and ERK signaling is involved in leptin-mediated promotion of invasion and migration of hepatocellular carcinoma cells. Cancer Res. 2007, 67, 2497–2507. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Zhang, Z.G.; Tian, X.Q.; Sun, D.F.; Liang, Q.C.; Zhang, Y.J.; Lu, R.; Chen, Y.X.; Fang, J.Y. Inhibition of JAK1, 2/STAT3 signaling induces apoptosis, cell cycle arrest, and reduces tumor cell invasion in colorectal cancer cells. Neoplasia 2008, 10, 287–297. [Google Scholar] [PubMed]
- Han, Q.B.; Li, M.L.; Li, S.H.; Mou, Y.K.; Lin, Z.W.; Sun, H.D. Ent-kauranediterpenoids from Isodonrubescens var. lushanensis. Chem. Pharm. Bull. 2003, 51, 790–793. [Google Scholar] [CrossRef]
- Zhang, J.X.; Han, Q.B.; Zhao, A.H.; Sun, H.D. Diterpenoids from Isodon japonica. Fitoterapia 2003, 74, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Osawa, K.; Yasuda, H.; Maruyama, T.; Morita, H.; Takeya, K.; Itokawa, H. Antibacterial trichorabdalditerpenes from Rabdosiatrichocarpa. Phytochemistry 1994, 36, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- Li, S.H.; Niu, X.M.; Peng, L.Y.; Zhang, H.J.; Yao, P.; Sun, H.D. Ent-kauranediterpenoids from the leaves of Isodonxerophilus. Planta Med. 2002, 68, 946–948. [Google Scholar] [CrossRef] [PubMed]
- Meade-Tollin, L.C.; Wijeratne, E.M.; Cooper, D.; Guild, M.; Jon, E.; Fritz, A.; Zhou, G.X.; Whitesell, L.; Liang, J.Y.; Gunatilaka, A.A. Ponicidin and oridonin are responsible for the antiangiogenic activity of Rabdosiarubescens, a constituent of the herbal supplement PC SPES. J. Nat. Prod. 2004, 67, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Correa, P.; Haenszel, W. Epidemiology of gastric cancer. Epidemiol. Cancer Digest. Tract 1982, 21, 59–84. [Google Scholar]
- Correa, P.; Haenszel, W.; Cuello, C.; Zavala, D.; Fontham, E.; Zarama, G.; Tannenbaum, S.; Collazos, T.; Ruiz, B. Gastric precancerous process in a high risk population: Cohort follow-up. Cancer Res. 1990, 50, 4737–4740. [Google Scholar] [PubMed]
- Correa, P.; Cuello, C.; Duque, E.; Burbano, L.C.; Garcia, F.T.; Bolanos, O.; Brown, C.; Haenszel, W. Gastric cancer in Colombia. III. Natural history of precursor lesions. J. Natl. Cancer Inst. 1976, 57, 1027–1035. [Google Scholar] [PubMed]
- Scott, N.; Diament, R.; Murday, V.; McMahon, M.; Lansdown, M.; Rathbone, B.; Wyatt, J.; Dixon, M.; Quirke, P. Helicobacter gastritis and intestinal metaplasia in a gastric cancer family. Lancet 1990, 335, 728. [Google Scholar]
- Talley, N.J.; Zinsmeister, A.R.; Weaver, A.; DiMagno, E.P.; Carpenter, H.A.; Perez-Perez, G.I.; Blaser, M.J. Gastric adenocarcinoma and Helicobacter pylori infection. J. Natl. Cancer Inst. 1991, 83, 1734–1739. [Google Scholar] [CrossRef] [PubMed]
- Martindale, J.L.; Holbrook, N.J. Cellular response to oxidative stress: Signaling for suicide and survival. J. Cell. Physiol. 2002, 192, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gloire, G.; Legrand-Poels, S.; Piette, J. NF-κB activation by reactive oxygen species: Fifteen years later. Biol. Pharmacol. 2006, 72, 1493–1505. [Google Scholar]
- Budihardjo, I.; Oliver, H.; Lutter, M.; Luo, X.; Wang, X. Biochemical pathways of caspase activation during apoptosis. Annu. Rev. Cell Dev. Biol. 1999, 15, 269–290. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Guo, J.; Wang, Z.; Xiao, B.; Lee, H.J.; Lee, E.O.; Kim, S.H.; Lu, J. Decursin and decursinolangelate inhibit estrogen-stimulated and estrogen-independent growth and survival of breast cancer cells. Breast Cancer Res. 2007, 9, R77. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Agarwal, R. Mechanisms of action of novel agents for prostate cancer chemoprevention. Endocr.-Relat. Cancer 2006, 13, 751–778. [Google Scholar] [CrossRef] [PubMed]
- Sagar, S.M.; Yance, D.; Wong, R.K. Natural health products that inhibit angiogenesis: A potential source for investigational new agents to treat cancer—Part 1. Curr. Oncol. 2006, 13, 14–26. [Google Scholar]
- Lee, H.J.; Lee, E.O.; Rhee, Y.H.; Ahn, K.S.; Li, G.X.; Jiang, C.; Lu, J.; Kim, S.H. An oriental herbal cocktail, ka-mi-kae-kyuk-tang, exerts anti-cancer activities by targeting angiogenesis, apoptosis and metastasis. Carcinogenesis 2006, 27, 2455–2463. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Huang, R.W.; Lin, D.J.; Peng, J.; Zhang, M.H.; Pan, X.L.; Hou, M.; Wu, X.Y.; Lin, Q.; Chen, F. Ponicidin, an ent-kaurane diterpenoid derived from a constituent of the herbal supplement PC-SPES, Rabdosia rubescens, induces apoptosis by activation of caspase-3 and mitochondrial events in lung cancer cells in vitro. Cancer Investig. 2006, 24, 136–148. [Google Scholar] [CrossRef]
- Zhang, J.F.; Liu, P.Q.; Chen, G.H.; Lu, M.Q.; Cai, C.J.; Yang, Y.; Li, H. Ponicidin inhibits cell growth on hepatocellular carcinoma cells by induction of apoptosis. Digest. Liver Dis. 2007, 39, 160–166. [Google Scholar] [CrossRef]
- Yu, H.; Pardoll, D.; Jove, R. STATs in cancer inflammation and immunity: A leading role for STAT3. Nat. Rev. Cancer 2009, 9, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Jove, R. The STATs of cancer—Newmolecular targets come of age. Nat. Rev. Cancer 2004, 4, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Niu, G.; Wright, K.L.; Huang, M.; Song, L.; Haura, E.; Turkson, J.; Zhang, S.; Wang, T.; Sinibaldi, D.; Coppola, D.; et al. Constitutive STAT3 activity up-regulates VEGF expression and tumor angiogenesis. Oncogene 2002, 21, 2000–2008. [Google Scholar] [CrossRef] [PubMed]
- Real, P.J.; Sierra, A.; de Juan, A.; Segovia, J.C.; Lopez-Vega, J.M.; Fernandez-Luna, J.L. Resistance to chemotherapy via STAT3-dependent overexpression of Bcl-2 in metastatic breast cancer cells. Oncogene 2002, 21, 7611–7618. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Niu, G.; Kortylewski, M.; Burdelya, L.; Shain, K.; Zhang, S.; Bhattacharya, R.; Gabrilovich, D.; Heller, R.; Coppola, D.; et al. Regulation of the innate and adaptive immune responses by STAT3 signaling in tumor cells. Nat. Med. 2004, 10, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Rossi, D.; Gaidano, G. Messengers of cell death: Apoptotic signaling in health and disease. Haematologica 2003, 88, 212–218. [Google Scholar] [PubMed]
- Tsuruo, T.; Naito, M.; Tomida, A.; Fujita, N.; Mashima, T.; Sakamoto, H.; Haga, N. Molecular targeting therapy of cancer: Drug resistance, apoptosis and survival signal. Cancer Sci. 2003, 94, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Debatin, K.M. Apoptosis pathways in cancer and cancer therapy. Cancer Immunol. Immun. 2004, 53, 153–159. [Google Scholar] [CrossRef]
- Kirkin, V.; Joos, S.; Zörnig, M. The role of Bcl-2 family members in tumorigenesis. BBA-Mol. Cell Res. 2004, 1644, 229–249. [Google Scholar]
- Manion, M.K.; Hockenbery, D.M. Targeting Bcl-2-related proteins in cancer therapy. Cancer Biol. Ther. 2003, 2, S105–S114. [Google Scholar] [CrossRef] [PubMed]
- Takaishi, S.; Okumura, T.; Tu, S.; Wang, S.S.; Shibata, W.; Vigneshwaran, R.; Gordon, S.A.; Shimada, Y.; Wang, T.C. Identification of gastric cancer stem cells using the cell surface marker CD44. Stem Cells 2009, 27, 1006–1020. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, L.F.; Ye, M.; Gu, H.H.; Cao, Y. Induction of apoptosis by epigallocatechin-3-gallate via mitochondrial signal transduction pathway. Prev. Med. 2004, 39, 1172–1179. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.-F.; Lu, Y.-M.; Qu, G.-Q.; Liu, Y.; Chen, W.-X.; Liao, X.-H.; Kong, W.-M. Ponicidin Induces Apoptosis via JAK2 and STAT3 Signaling Pathways in Gastric Carcinoma. Int. J. Mol. Sci. 2015, 16, 1576-1589. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16011576
Liu Y-F, Lu Y-M, Qu G-Q, Liu Y, Chen W-X, Liao X-H, Kong W-M. Ponicidin Induces Apoptosis via JAK2 and STAT3 Signaling Pathways in Gastric Carcinoma. International Journal of Molecular Sciences. 2015; 16(1):1576-1589. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16011576
Chicago/Turabian StyleLiu, Yuan-Fei, Yun-Min Lu, Guo-Qiang Qu, Yuan Liu, Wei-Xiong Chen, Xiao-Hong Liao, and Wu-Ming Kong. 2015. "Ponicidin Induces Apoptosis via JAK2 and STAT3 Signaling Pathways in Gastric Carcinoma" International Journal of Molecular Sciences 16, no. 1: 1576-1589. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16011576