
Molecular Characterization and Biological Effects of a C-Type Lectin-Like Receptor in Large Yellow Croaker (Larimichthys crocea)

Abstract
:1. Introduction
2. Results
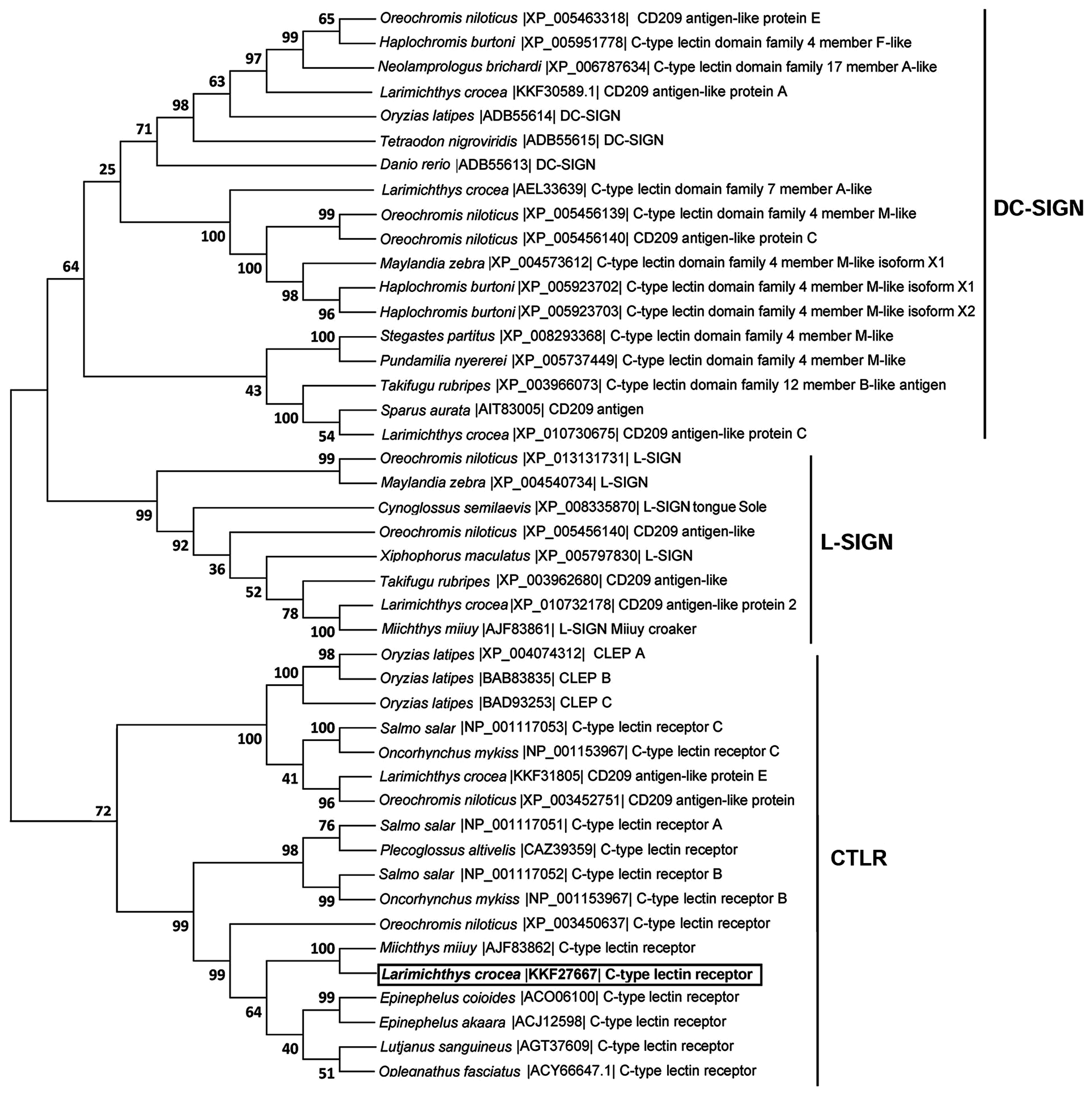
2.1. cDNA and Gene Structures of LycCTLR


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Common Name | Molecules | Identities (%) | NCBI Accession Number |
|---|---|---|---|---|
| C-Type Lectin Receptor | ||||
| Miichthys miiuy | Miiuy croaker | C-type lectin-like receptor | 88.4 | AJF83862 |
| Oplegnathus fasciatus | Barred knifejaw | C-type lectin receptor | 62.3 | ACY66647 |
| Lutjanus sanguineus | Humphead snapper | C-type lectin receptor | 61.9 | AGT37609 |
| Epinephelus coioides | Orange-spotted grouper | C-type lectin receptor | 59.5 | ACO06100 |
| Epinephelus akaara | Hong Kong grouper | C-type lectin receptor | 58.1 | ACJ12598 |
| Oreochromis niloticus | Tilapia | C-type lectin receptor | 56.6 | XP_003450637 |
| Salmo salar | Salmon A | C-type lectin receptor A | 45.6 | NP_001117051 |
| Oncorhynchus mykiss | Trout B | C-type lectin receptor B | 42.8 | NP_001153967 |
| Plecoglossus altivelis | Ayu | C-type lectin receptor | 35 | CAZ39359 |
| Species | Common Name | Molecules | Identities (%) | NCBI Accession Number |
| DC-SIGN | ||||
| Oryzias latipes | Medaka | DC-SIGN | 20.0 | ADB55614 |
| Tetraodon nigroviridis | Tetraodon | DC-SIGN | 19.5 | ADB55615 |
| Oreochromis niloticus | Tilapia | DC-SIGN | 19.1 | XP_005463318 |
| Larimichthys crocea | Large yellow croaker | DC-SIGN | 18.7 | KKF30589 |
| Danio rerio | Zebrafish | DC-SIGN | 14.0 | ADB55613 |
| Species | Common Name | Molecules | Identities (%) | NCBI Accession Number |
| l -SIGN | ||||
| Takifugu rubripes | Fugu | l-SIGN | 18.1 | XP_003962680 |
| Cynoglossus semilaevis | Tongue sole | l-SIGN | 15.9 | XP_008335870 |
| Miichthys miiuy | Miiuy croaker | l-SIGN | 15.4 | AJF83861 |
| Oreochromis niloticus | Tilapia | l-SIGN | 14.4 | XP_005456140 |
| Larimichthys crocea | Large yellow croaker | l-SIGN | 14.4 | XP_010732178 |


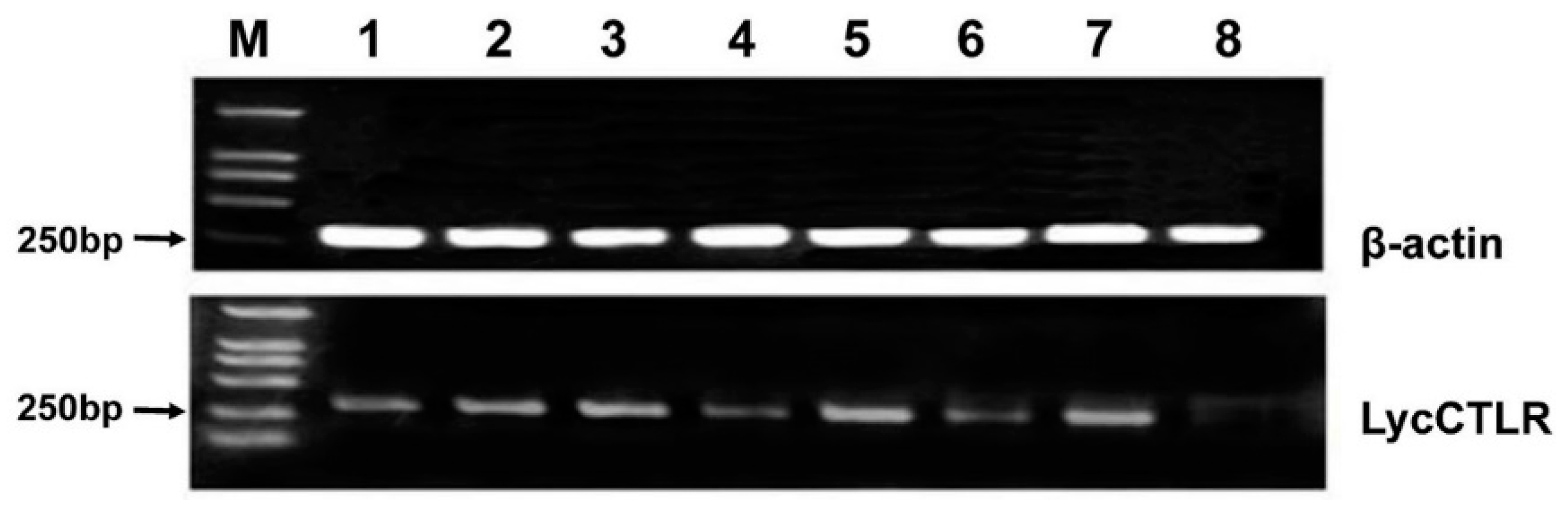
2.2. Tissue Expression Analysis of LycCTLR Gene


2.3. HemagglutinationActivity of LycCTLR
| Erythrocytes | Minimum Agglutinating Concentration of rLycCLTR (μg/mL) |
|---|---|
| Rabbit | 4 |
| Rat | 8 |
| Mouse | 64 |
| Tilapia | NA a |
2.4. Sugar Binding SpecificityAssays
| Saccharides | Minimal Inhibitory Concentration (mM) |
|---|---|
| d-galactose | 12.5 |
| d-xylose | 50 |
| l-fucose | 50 |
| Cellobiose | 50 |
| Maltotriose | 100 |
| d-fructose | 200 |
| d-glucose | 200 |
| d-mannitol | 200 |
| Maltose | 200 |
| Sobitol | 200 |
| d-(−)-arabinose | NI a |
| d-mannose | NI |
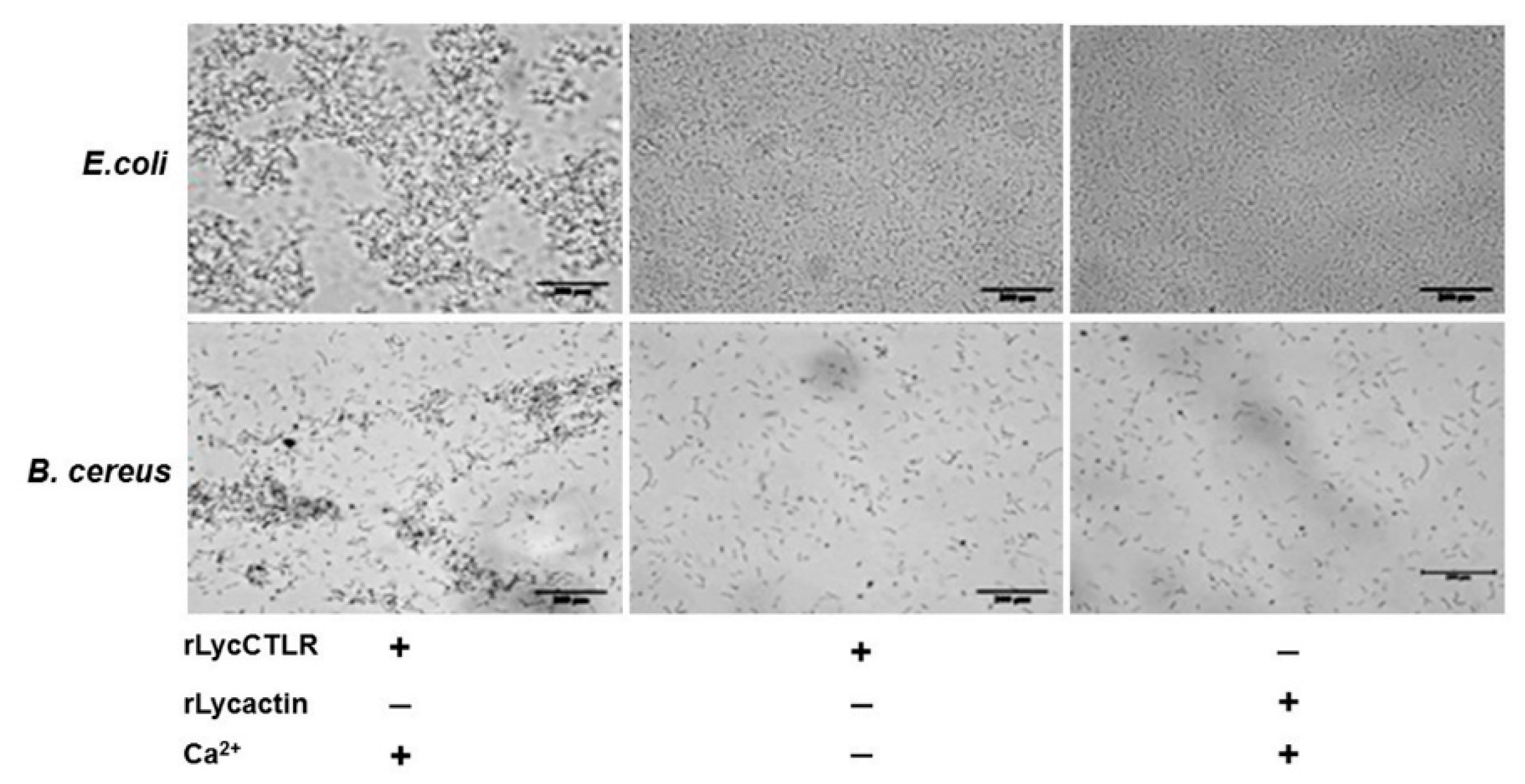
2.5. Bacterial Agglutinationassays

3. Discussion
4. Materials and Methods
4.1. Fish and Induction Experiment
4.2. Cloning of the LycCTLR
4.3. Sequence Analysis and Database
4.4. Tissue Expression Analysis of LycCTLR Gene
4.5. Expression Modulation Analysis of LycCTLR Gene
4.6. Expression and Purification of Recombinant LycCTLR
4.7. Hemagglutination Assays
4.8. Sugar Binding Assays
4.9. Bacterial Agglutination Assays
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Aoki, T.; Hikima, J.; Hwang, S.D.; Jung, T.S. Innate immunity of finfish: Primordial conservation and function of viral rna sensors in teleosts. Fish Shellfish Immunol. 2013, 35, 1689–1702. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.D.; Vojtech, L.N.; Laing, K.J. Sensing disease and danger: A survey of vertebrate PRRs and their origins. Dev. Comp. Immunol. 2011, 35, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Plato, A.; Willment, J.A.; Brown, G.D. C-type lectin-like receptors of the dectin-1 cluster: Ligands and signaling pathways. Int. Rev. Immunol. 2013, 32, 134–156. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; Gringhuis, S.I. Signalling through C-type lectin receptors: Shaping immune responses. Nat. Rev. Immunol. 2009, 9, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Tizard, I. Lectin-carbohydrate interaction in the immune system. Vet. Immunol. Immunopathol. 1996, 55, 205–223. [Google Scholar] [CrossRef]
- Ebner, S.; Ehammer, Z.; Holzmann, S.; Schwingshackl, P.; Forstner, M.; Stoitzner, P.; Huemer, G.M.; Fritsch, P.; Romani, N. Expression of C-type lectin receptors by subsets of dendritic cells in human skin. Int. Immunol. 2004, 16, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Hoving, J.C.; Wilson, G.J.; Brown, G.D. Signalling C-type lectin receptors, microbial recognition and immunity. Cell Microbiol. 2014, 16, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Kerrigan, A.M.; Brown, G.D. Syk-coupled C-type lectin receptors that mediate cellular activation via single tyrosine based activation motifs. Immunol. Rev. 2010, 234, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Mayer, W.E.; Overath, P.; Klein, J. Genes encoding putative natural killer cell C-type lectin receptors in teleostean fishes. Proc. Natl. Acad. Sci. USA 2003, 100, 7779–7784. [Google Scholar] [CrossRef] [PubMed]
- Soanes, K.H.; Figuereido, K.; Richards, R.C.; Mattatall, N.R.; Ewart, K.V. Sequence and expression of C-type lectin receptors in atlantic salmon (Salmo salar). Immunogenetics 2004, 56, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Panagos, P.G.; Dobrinski, K.P.; Chen, X.; Grant, A.W.; Traver, D.; Djeu, J.Y.; Wei, S.; Yoder, J.A. Immune-related, lectin-like receptors are differentially expressed in the myeloid and lymphoid lineages of zebrafish. Immunogenetics 2006, 58, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Asim, M.; Lan, J.; Zhao, L.; Wei, S.; Chen, N.; Liu, X.; Zhou, Y.; Lin, L. Molecular cloning and functional characterization of mannose receptor in zebra fish (Danio rerio) during infection with aeromonas sobria. Int. J. Mol. Sci. 2015, 16, 10997–11012. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, L.; Zhou, Y.; Zhao, X.; Xi, M.; Wei, S.; Fang, R.; Ji, W.; Chen, N.; Gu, Z.; et al. Molecular cloning and expression analysis of mannose receptor C type 1 in grass carp (Ctenopharyngodon idella). Dev. Comp. Immunol. 2014, 43, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, L.; Hegazy, A.M.; Wang, H.; Li, J.; Zheng, F.; Zhou, Y.; Wang, W.; Li, J.; Liu, X.; et al. Mannose receptor mediated phagocytosis of bacteria in macrophages of blunt snout bream (Megalobrama amblycephala) in a Ca2+-dependent manner. Fish Shellfish Immunol. 2015, 43, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lu, X.J.; Yang, H.Y.; Shi, Y.H. An interaction between a C-type lectin receptor and leukocyte cell-derived chemotaxin 2 of ayu, Plecoglossus altivelis. Fish Shellfish Immunol. 2010, 28, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.J.; Lu, X.J.; Chen, Q.; Chen, J. Molecular characterization and functional analysis of a novel C-type lectin receptor-like gene from a teleost fish, Plecoglossus altivelis. Fish Shellfish Immunol. 2015, 44, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Shi, Y.H.; Chen, J. Molecular characterization of a transmembrane C-type lectin receptor gene from ayu (Plecoglossus altivelis) and its effect on the recognition of different bacteria by monocytes/macrophages. Mol. Immunol. 2015, 66, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.F.; Xiang, L.X.; Wang, Q.L.; Dong, W.R.; Gong, Y.F.; Shao, J.Z. The DC-SIGN of zebrafish: Insights into the existence of a CD209 homologue in a lower vertebrate and its involvement in adaptive immunity. J. Immunol. 2009, 183, 7398–7410. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Drickamer, K. Engineering galactose-binding activity into a C-type mannose-binding protein. Nature 1992, 360, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Soanes, K.H.; Ewart, K.V.; Mattatall, N.R. Recombinant production and characterization of the carbohydrate recognition domain from atlantic salmon C-type lectin receptor c (SCLRC). Protein Expr. Purif. 2008, 59, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Yang, H.; Li, F.; Zhang, X.; Xu, X. Purification, characterization and cDNA cloning of a novel lipopolysaccharide-binding lectin from the shrimp Penaeus monodon. Dev. Comp. Immunol. 2006, 30, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Drickamer, K.; Taylor, M.E. Recent insights into structures and functions of C-type lectins in the immune system. Curr. Opin. Struc. Biol. 2015, 34, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Liu, X.; Yang, J.; Wang, S.; Sun, G.; Yang, J. Critical roles of sea cucumber C-type lectin in non-self recognition and bacterial clearance. Fish Shellfish Immunol. 2015, 45, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Ayulo, A.M.; Machado, R.A.; Scussel, V.M. Enterotoxigenic Escherichia coli and Staphylococcus aureus in fish and seafood from the southern region of brazil. Int. J. Food Microbiol. 1994, 24, 171–178. [Google Scholar] [CrossRef]
- Ceuppens, S.; Rajkovic, A.; Heyndrickx, M.; Tsilia, V.; van de Wiele, T.; Boon, N.; Uyttendaele, M. Regulation of toxin production by Bacillus cereus and its food safety implications. Crit. Rev. Microbiol. 2011, 37, 188–213. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Tian, C.; Chen, X. Molecular characterization of goose-type lysozyme homologue of large yellow croaker and its involvement in immune response induced by trivalent bacterial vaccine as an acute-phase protein. Immunol. Lett. 2007, 113, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Liu, G.; Ao, J.; Chen, X. Expression analysis of immune-relevant genes in the spleen of large yellow croaker (Pseudosciaena crocea) stimulated with poly I:C. Fish Shellfish Immunol. 2006, 21, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Ao, J.; Mu, Y.; Xiang, L.X.; Fan, D.; Feng, M.; Zhang, S.; Shi, Q.; Zhu, L.Y.; Li, T.; Ding, Y.; et al. Genome sequencing of the perciform fish Larimichthys crocea provides insights into molecular and genetic mechanisms of stress adaptation. PLoS Genet. 2015, 11, e1005118. [Google Scholar] [CrossRef] [PubMed]
- BoxShade. Available online: http://www.ch.embnet.org/software/BOX_form.html (accessed on 1 October 2015).
- Clustal Omega. Available online: http://www.ebi.ac.uk/Tools/msa/clustalo/ (accessed on 1 October 2015).
- SMART. Available online: http://smart.embl-heidelberg.de/ (accessed on 1 October 2015).
- TMHMM Server v.2.1. Available online: http://www.cbs.dtu.dk/services/TMHMM/ (accessed on 1 October 2015).
- Yu, S.; Chen, X.; Ao, J. Molecular characterization and expression analysis of β2-microglobulin in large yellow croaker Pseudosciaena crocea. Mol. Biol. Rep. 2009, 36, 1715–1723. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Li, M.; Wang, K.; Qin, Q.; Chen, X. Characterization of large yellow croaker (Pseudosciaena crocea) β-actin promoter supports β-actin gene as an internal control for gene expression modulation and its potential application in transgenic studies in fish. Fish Shellfish Immunol. 2011, 30, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Chen, Y.; Ao, J.; Chen, X. Molecular characterization and bioactivity of a CXCL13 chemokine in large yellow croaker Pseudosciaena crocea. Fish Shellfish Immunol. 2010, 28, 445–452. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ao, J.; Ding, Y.; Chen, Y.; Mu, Y.; Chen, X. Molecular Characterization and Biological Effects of a C-Type Lectin-Like Receptor in Large Yellow Croaker (Larimichthys crocea). Int. J. Mol. Sci. 2015, 16, 29631-29642. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226175
Ao J, Ding Y, Chen Y, Mu Y, Chen X. Molecular Characterization and Biological Effects of a C-Type Lectin-Like Receptor in Large Yellow Croaker (Larimichthys crocea). International Journal of Molecular Sciences. 2015; 16(12):29631-29642. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226175
Chicago/Turabian StyleAo, Jingqun, Yang Ding, Yuanyuan Chen, Yinnan Mu, and Xinhua Chen. 2015. "Molecular Characterization and Biological Effects of a C-Type Lectin-Like Receptor in Large Yellow Croaker (Larimichthys crocea)" International Journal of Molecular Sciences 16, no. 12: 29631-29642. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226175