
Isolation and Molecular Characterization of 1-Aminocyclopropane-1-carboxylic Acid Synthase Genes in Hevea brasiliensis

Abstract
:1. Introduction
2. Results
2.1. Cloning, Identification, and Structure Analysis of the HbACS Gene Family

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank Accession No. | ORF (bp) | Predicted Protein | |||
|---|---|---|---|---|---|---|
| Size (aa) | Type | Mw (kDa) | pI | |||
| HbACS1 | KJ911898 | 1443 | 480 | Type 1 | 54.05 | 6.35 |
| HbACS2 | KJ911899 | 1446 | 481 | Type 1 | 54.43 | 5.70 |
| HbACS3 | KJ911900 | 1452 | 483 | Type 1 | 54.20 | 5.74 |
| HbACS4 | KJ911901 | 1455 | 484 | Type 1 | 54.60 | 7.24 |
| HbACS5 | KJ911902 | 1398 | 465 | Type 2 | 52.66 | 8.94 |
| HbACS6 | KJ911903 | 1323 | 440 | Type 3 | 49.18 | 5.69 |
| HbACS7 | KJ911904 | 1323 | 440 | Type 3 | 49.25 | 5.58 |
| HbACS8 | KJ911905 | 1608 | 535 | Putative AAT | 59.24 | 8.23 |
| HbACS9 | KJ911906 | 1653 | 550 | Putative AAT | 60.48 | 6.23 |
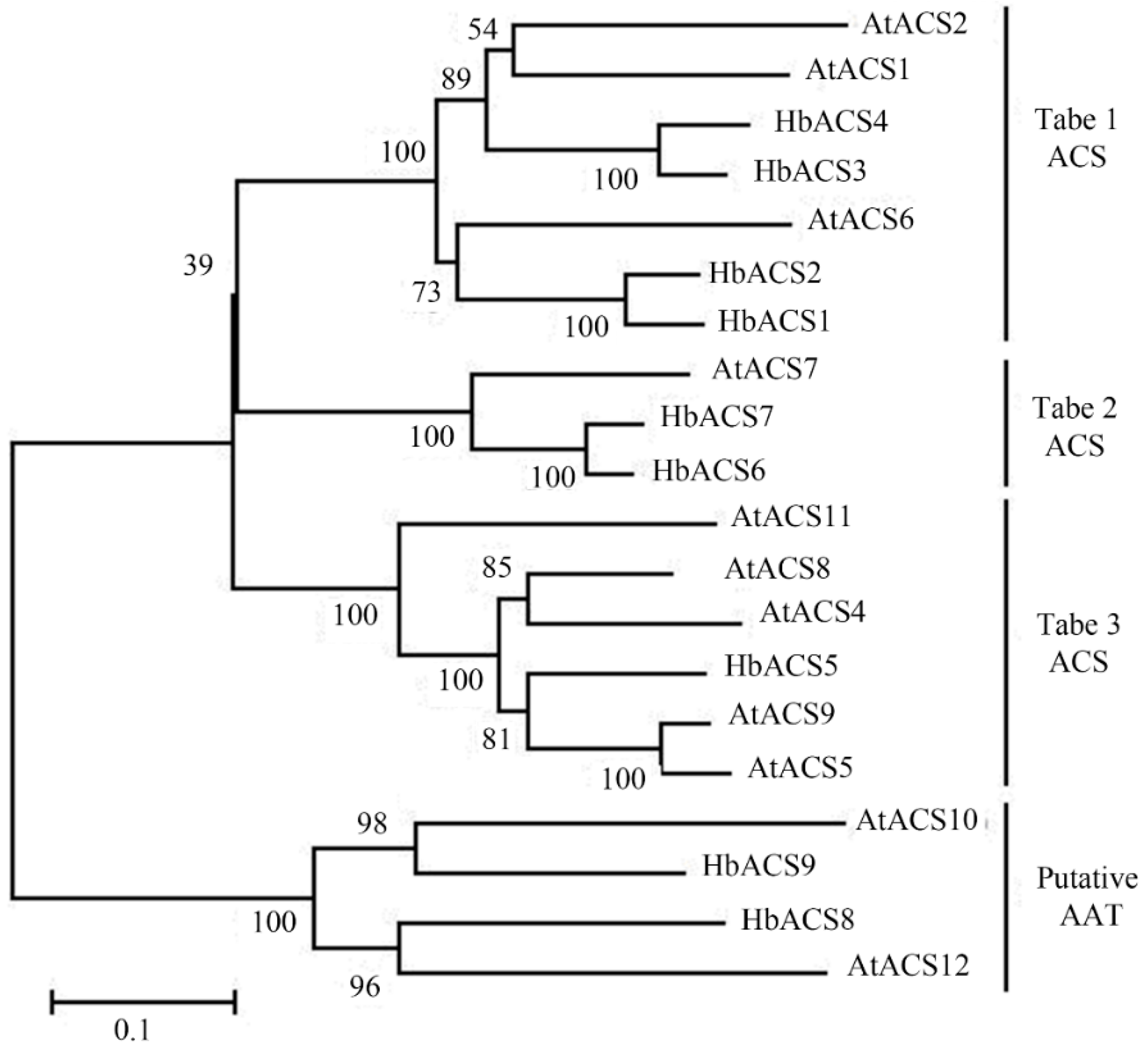
2.2. Phylogenetic Analysis

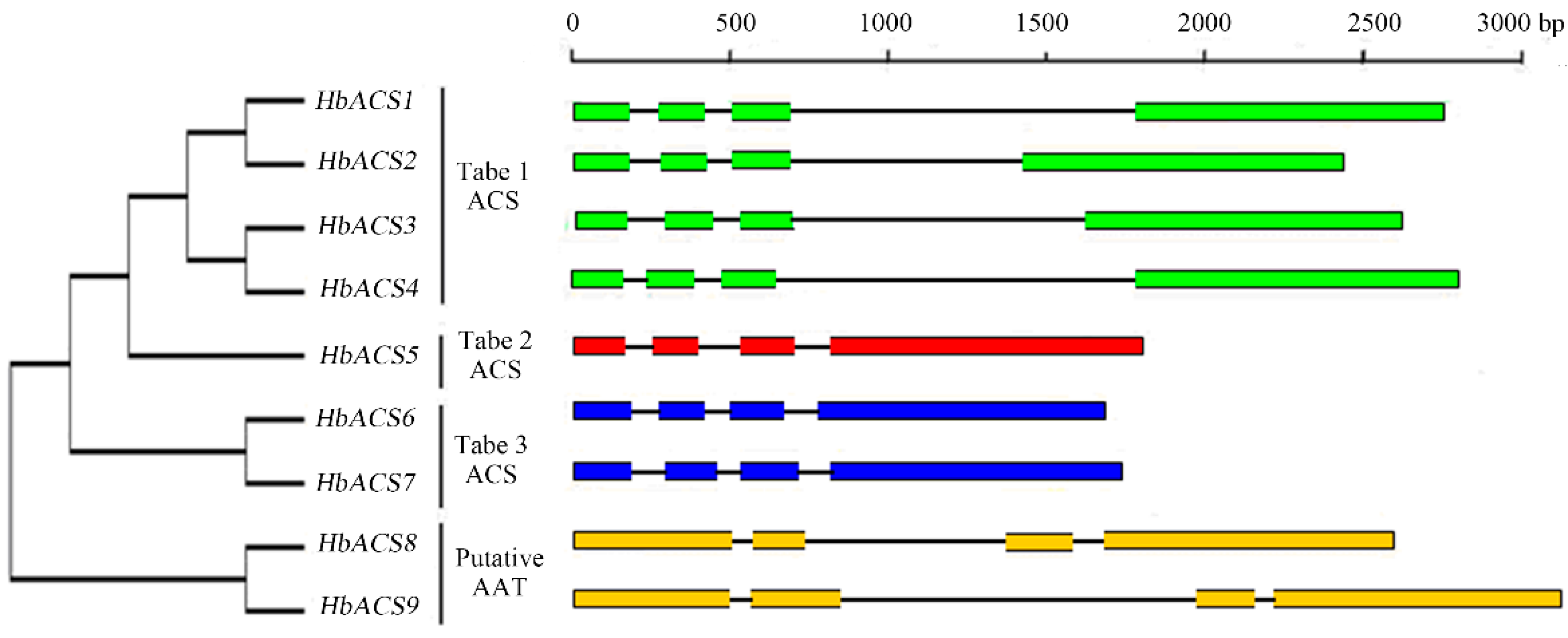
2.3. Intron and Exon Organization of HbACS Genes


| Gene | Size | Hormone Response Element | Stress Response Element | Other Element |
|---|---|---|---|---|
| HbASC1 | 823 | CGTCA-motif, TATC-box | MBS | GCN4_motif, Skn-1_motif |
| HbASC2 | 1484 | CGTCA-motif, ERE GARE-motif, TATC-box TGA-element | MBS HSE TC-rich repeats | O2-site, Skn-1_motif, as-2-box |
| HbASC3 | 1625 | ABRE, CGTCA-motif, ERE, TCA-element | AT-rich element, HSE | GCN4_motif, Skn-1_motif |
| HbASC4 | 1553 | ABRE, P-box, TGA-element | ARE, HSE, LTR, MBS, TC-rich repeats | Skn-1_motif, as-2-box, circadian |
| HbASC5 | 1575 | ABRE, AuxRR-core, GARE-motif | ARE, Box-W1, C-repeat/DRE, HSE, MBS, TC-rich repeats, W box | GCN4_motif, Skn-1_motif |
| HbASC6 | 1110 | GARE-motif, TGA-element | Box-W1, HSE, MBS, TC-rich repeats, W box | O2-site, Skn-1_motif2 |
| HbASC7 | 2036 | P-box, AuxRR-core, GARE-motif, TCA-element | ARE, HSE, MBS, TC-rich repeats | CAT-box, O2-site, RY-element, Skn-1_motif, circadian |
2.4. Cis-Elements of HbACS Promoters
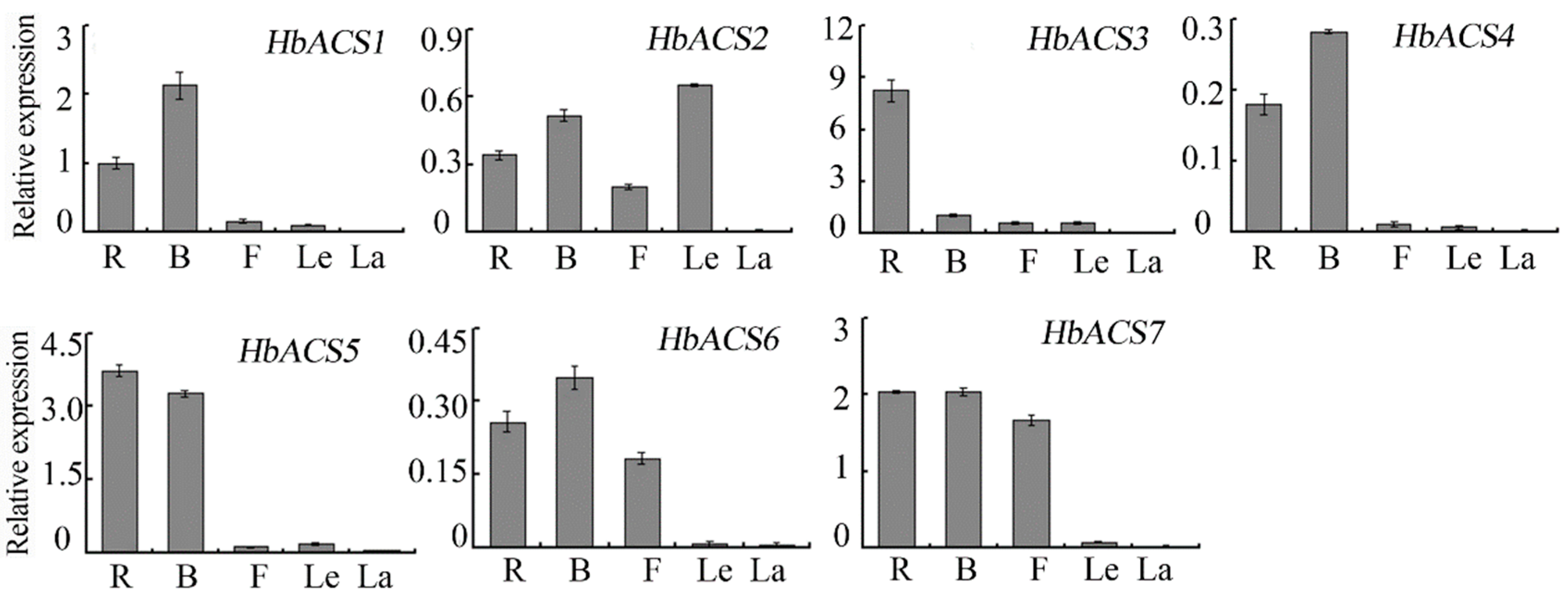
2.5. Expression Analysis of HbACS Genes in Hevea Tissues


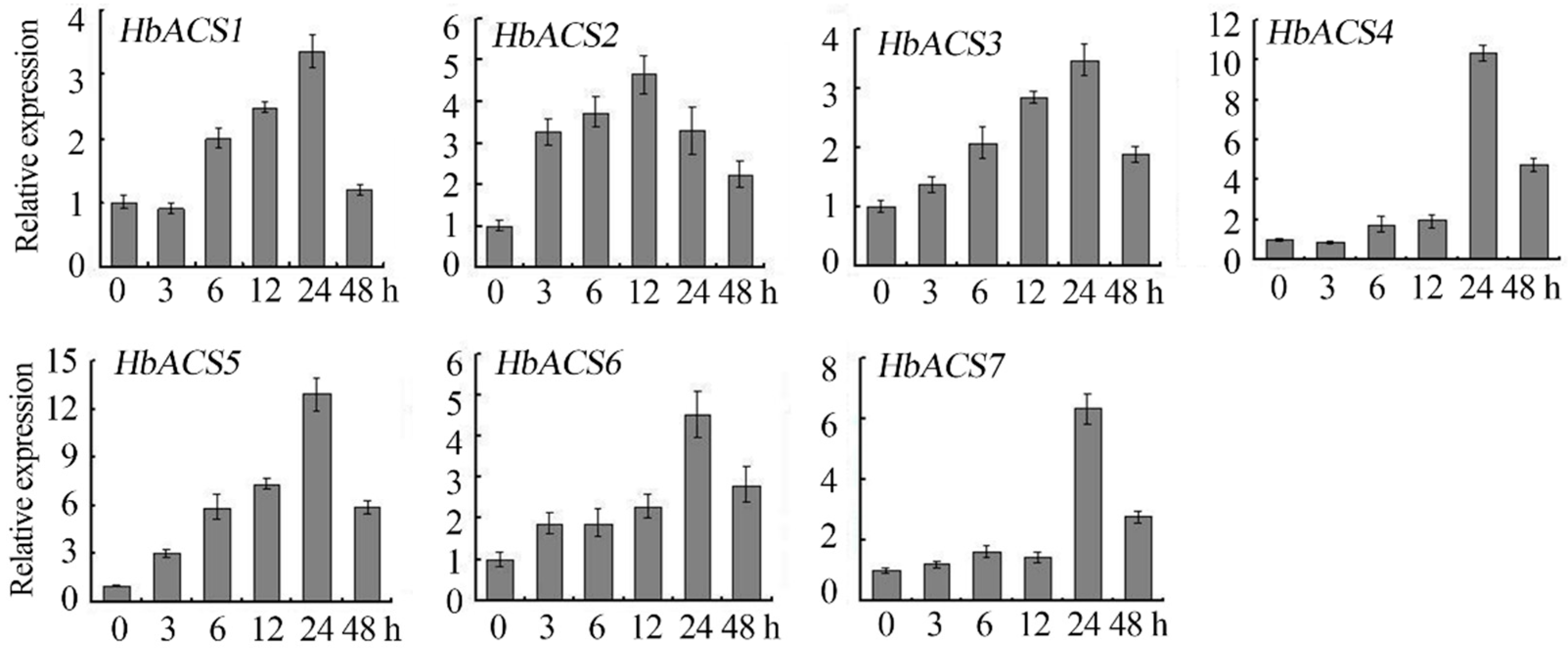
2.6. Expression Analyses of HbACS Genes in Response to Ethrel Treatment
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Nucleic Acid Extraction and cDNA Synthesis
4.3. Cloning and Identification of ACS Genes
4.4. Promoter Region Analysis of HbACS Genes
4.5. Expression Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, F.; Cui, X.; Sun, Y.; Dong, C.H. Ethylene signaling and regulation in plant growth and stress responses. Plant Cell Rep. 2013, 32, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.J.; Chang, C.L.; Wang, P.H.; Tsai, M.C.; Hsu, P.H.; Chang, I.F. A type III ACC synthase, ACS7, is involved in root gravitropism in Arabidopsis thaliana. J. Exp. Bot. 2013, 64, 4343–4360. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.O.; Yang, S.F. Ethylene biosynthesis: Identification of 1-aminocyclopropane-1-carboxylic acid as an intermediate in the conversion of methionine to ethylene. Proc. Natl. Acad. Sci. USA. 1979, 76, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.F.; Zhong, S.L.; Grierson, D. Recent advances in ethylene research. J. Exp. Bot. 2009, 60, 3311–3336. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, S. Phosphorylation of 1-aminocyclopropane-1-carboxylic acid synthase by MPK6, a stress-responsive mitogen-activated protein kinase, induces ethylene biosynthesis in Arabidopsis. Plant Cell 2004, 16, 3386–3399. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.R.; Roy, S.; Sengupta, D.N. C-Terminal phosphorylation is essential for regulation of ethylene synthesizing ACC synthase enzyme. Plant Signal. Behav. 2013, 8, 491–511. [Google Scholar]
- Yamagami, T.; Tsuchisaka, A.; Yamada, K.; Haddon, W.F.; Harden, L.A.; Theologis, A. Biochemical diversity among the 1-amino-cyclopropane-1-carboxylate synthase isozymes encoded by the Arabidopsis gene family. J. Biol. Chem. 2003, 278, 49102–49112. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.P.; Lin, T.Y.; Wang, N.N.; Shih, M.C. Differential expression of genes encoding 1-aminocyclopropane-1-carboxylate synthase in Arabidopsis during hypoxia. Plant Mol. Biol. 2005, 58, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.N.; Shih, M.C.; Li, N. The GUS reporter-aided analysis of the promoter activities of Arabidopsis ACC synthase genes AtACS4, AtACS5, and AtACS7 induced by hormones and stresses. J. Exp. Bot. 2005, 56, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Theologis, A. Cloning the mRNA encoding 1-aminocyclopropane-1-carboxylate synthase, the key enzyme for ethylene biosynthesis in plants. Proc. Natl. Acad. Sci. USA 1989, 86, 6621–6625. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, I.; Kim, W.S.; Jayasankar, S.; Svircev, A.M.; Brown, D.C. Differential regulation of four members of the ACC synthase gene family in plum. J. Exp. Bot. 2008, 59, 2009–2027. [Google Scholar] [CrossRef] [PubMed]
- Kamiyoshihara, Y.; Iwata, M.; Fukaya, T.; Tatsuki, M.; Mori, H. Turnover of LeACS2, a wound-inducible1-aminocyclopropane-1-carboxylic acid synthase in tomato, is regulated by phosphorylation/dephosphorylation. Plant J. 2010, 64, 140–150. [Google Scholar] [PubMed]
- Harada, T.; Murakoshi, Y.; Torii, Y.; Tanase, K.; Onozaki, T.; Morita, S.; Masumura, T.; Satoh, S. Analysis of genomic DNA of DcACS1, a 1-aminocyclopropane-1-carboxylate synthase gene, expressed in senescing petals of carnation (Dianthus caryophyllus) and its orthologous genes in D. superbus var. longicalycinus. Plant Cell Rep. 2011, 30, 519–527. [Google Scholar] [CrossRef]
- D’ Auzac, J.; Jacob, J.L.; Prevot, J.C.; Clement, A.; Gallois, R.; Crestin, H.; Lacote, R.; Pujade Renaud, V.; Gohet, E. The regulation of cis-polyisoprene production natural rubber from Hevea brasiliensis. Recent Res. Dev. Plant Physiol. 1997, 1, 273–332. [Google Scholar]
- Hao, B.Z.; Wu, J.L. Laticifer differentiation in Hevea brasiliensis: Induction by exogenous jasmonic acid and linolenic acid. Ann. Bot. 2000, 85, 37–43. [Google Scholar] [CrossRef]
- Dusotoit-Coucaud, A.; Brunel, N.; Kongsawadworakul, P.; Viboonjun, U.; Lacointe, A.; Julien, J.L.; Chrestin, H.; Sakr, S. Sucrose importation into laticifers of Hevea brasiliensis, in relation to ethylene stimulation of latex production. Ann. Bot. 2009, 104, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.W.; Yang, S.Q. Ethylene physiology of Hevea brasiliensis. Trop. Crop Res. 1991, 12, 69–77. [Google Scholar]
- Zhu, J.H.; Zhang, Z.L. Ethylene stimulation of latex production in Hevea brasiliensis. Plant Signal. Behav. 2009, 4, 1–3. [Google Scholar] [CrossRef]
- Duan, C.; Argout, X.; Gébelin, V.; Summo, M.; Dufayard, J.F.; Leclercq, J.; Kuswanhadi; Piyatrakul, P.; Pirrello, J.; Rio, M.; et al. Identification of the Hevea brasiliensis AP2/ERF superfamily by RNA sequencing. BMC Genomics 2013, 14, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuswanhadi; Leclercq, J.; Rio, M.; Tregear, J.; Ducamp-Collin, M.N.; Montoro, P. Isolation of three members of the multigene family encoding ACC oxidases in Hevea brasiliensis and investigation of their responses to ethylene stimulation and wounding. J. Rubber Res. 2010, 13, 185–205. [Google Scholar]
- Piyatrakul, P.; Putranto, R.A.; Martin, F.; Rio, M.; Dessailly, F.; Leclercq, J.; Dufayard, J.F.; Lardet, L.; Montoro, P. Some ethylene biosynthesis and AP2/ERF genes reveal a specific pattern of expression during somatic embryogenesis in Hevea brasiliensis. BMC Plant Biol. 2012, 12, 244. [Google Scholar] [CrossRef]
- Triwitayakorn, K.; Chatkulkawin, P.; Kanjanawattanawong, S.; Sraphet, S.; Yoocha, T.; Sangsrakru, D.; Chanprasert, J.; Ngamphiw, C.; Jomchai, N.; Therawattanasuk, K.; et al. Transcriptome sequencing of Hevea brasiliensis for development of microsatellite markers and construction of a genetic linkage map. DNA Res. 2011, 18, 471–482. [Google Scholar] [CrossRef]
- Xia, Z.; Xu, H.; Zhai, J.; Li, D.; Luo, H.; He, C.; Huang, X. RNA-Seq analysis and de novo transcriptome assembly of Hevea brasiliensis. Plant Mol. Biol. 2011, 77, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Deng, Z.; Qin, B.; Liu, X.; Men, Z. De novo assembly and characterization of bark transcriptome using Illumina sequencing and development of EST-SSR markers in rubber tree (Hevea brasiliensis Muell. Arg.). BMC Genomics 2012, 13, 192. [Google Scholar] [CrossRef] [PubMed]
- Lertpanyasampatha, M.; Gao, L.; Kongsawadworakul, P.; Viboonjun, U.; Chrestin, H.; Liu, R.; Chen, X.; Narangajavana, J. Genome-wide analysis of microRNAs in rubber tree (Hevea brasiliensis L.) using high-throughput sequencing. Planta 2012, 236, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.Y.; Usharraj, A.O.; Misra, B.B.; Thottathil, G.P.; Jayasekaran, K.; Feng, Y.; Hou, S.; Ong, S.Y.; Ng, F.L.; Lee, L.S.; et al. Draft genome sequence of the rubber tree Hevea brasiliensis. BMC Genomics 2013, 14, 75. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.S.P.; Tecson-Mendoza, E.M.; Laurena, A.C.; Botella, J.R. Hybrid “Sinta” Papaya exhibits unique ACC synthase 1 cDNA isoforms. J. Biochem. Mol. Biol. 2005, 38, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.H.; Wang, M.H. Genome-wide analysis of 1-amino-cyclopropane-1-carboxylate synthase gene family in Arabidopsis, rice, grapevine and poplar. Afr. J. Biotechnol. 2012, 11, 1106–1118. [Google Scholar] [CrossRef]
- Chae, H.S.; Kieber, J.J. Eto Brute? Role of ACS turnover in regulating ethylene biosynthesis. Trends Plant Sci. 2005, 10, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Tatsuki, M.; Mori, H. Phosphorylation of tomato 1-aminocyclopropane-1-carboxylic acid synthase, LE-ACS2, at the C-terminal region. J. Biol. Chem. 2001, 276, 28051–28057. [Google Scholar] [CrossRef] [PubMed]
- Lyzenga, W.J.; Stone, S.L. Regulation of ethylene biosynthesis through protein degradation. Plant Signal. Behav. 2012, 7, 1438–1442. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.L.; Woodson, W.R. Differential expression of three members of the 1-aminocyclopropane-1-carboxylate synthase gene family in carnation. Plant Physiol. 1999, 119, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Chen, Z.; Gao, J.; Gong, Z. Abscisic acid inhibits root growth in Arabidopsis through ethylene biosynthesis. Plant J. 2014, 79, 44–55. [Google Scholar] [CrossRef]
- Joseph, R.E. Reentry of the ethylene MPK6 module. Plant Cell. 2004, 16, 3169–3173. [Google Scholar] [CrossRef]
- Van de Poel, B.; van der Straeten, D. 1-Aminocyclopropane-1-carboxylic acid (ACC) in plants: More than just theprecursor of ethylene. Front. Plant Sci. 2014, 5, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, G.C.; Flores-Vergara, M.A.; Krasynanski, S.; Kumar, S.; Thompson, W.F. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nat. Protoc. 2006, 1, 2320–2325. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Qi, J.; Li, H.; Zhang, C.; Wang, Y. A convenient and efficient protocol for isolating high quality RNA from latex of Hevea brasiliensis (para rubber tree). J. Biochem. Biophys. Methods 2007, 70, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.H.; Luo, M.W.; Zhang, C.F.; Zeng, H.C. Extraction of high-quality RNA from rubber tree leaves. Biosci. Biotechnol. Biochem. 2012, 76, 1394–1396. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qin, Y.; Xiao, X.; Tang, C. Screening of valid reference genes for real-time RT-PCR data normalization in Hevea brasiliensis and expression validation of a sucrose transporter gene HbSUT3. Plant Sci. 2011, 181, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Tang, C.; Fang, Y.; Yang, M.; Zhou, B.; Qi, J.; Zhang, Y. Structure and expression profile of the sucrose synthase gene family in the rubber tree: Indicative of roles in stress response and sucrose utilization in the laticifers. FEBS J. 2014, 281, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Remans, T.; Smeets, K.; Opdenakker, K.; Mathijsen, D.; Vangronsveld, J.; Cuypers, A. Normalisation of real-time RT-PCR gene expression measurements in Arabidopsis thaliana exposed to increased metal concentrations. Planta 2008, 227, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.-H.; Xu, J.; Chang, W.-J.; Zhang, Z.-L. Isolation and Molecular Characterization of 1-Aminocyclopropane-1-carboxylic Acid Synthase Genes in Hevea brasiliensis. Int. J. Mol. Sci. 2015, 16, 4136-4149. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16024136
Zhu J-H, Xu J, Chang W-J, Zhang Z-L. Isolation and Molecular Characterization of 1-Aminocyclopropane-1-carboxylic Acid Synthase Genes in Hevea brasiliensis. International Journal of Molecular Sciences. 2015; 16(2):4136-4149. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16024136
Chicago/Turabian StyleZhu, Jia-Hong, Jing Xu, Wen-Jun Chang, and Zhi-Li Zhang. 2015. "Isolation and Molecular Characterization of 1-Aminocyclopropane-1-carboxylic Acid Synthase Genes in Hevea brasiliensis" International Journal of Molecular Sciences 16, no. 2: 4136-4149. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16024136