
Nerve Demyelination Increases Metabotropic Glutamate Receptor Subtype 5 Expression in Peripheral Painful Mononeuropathy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
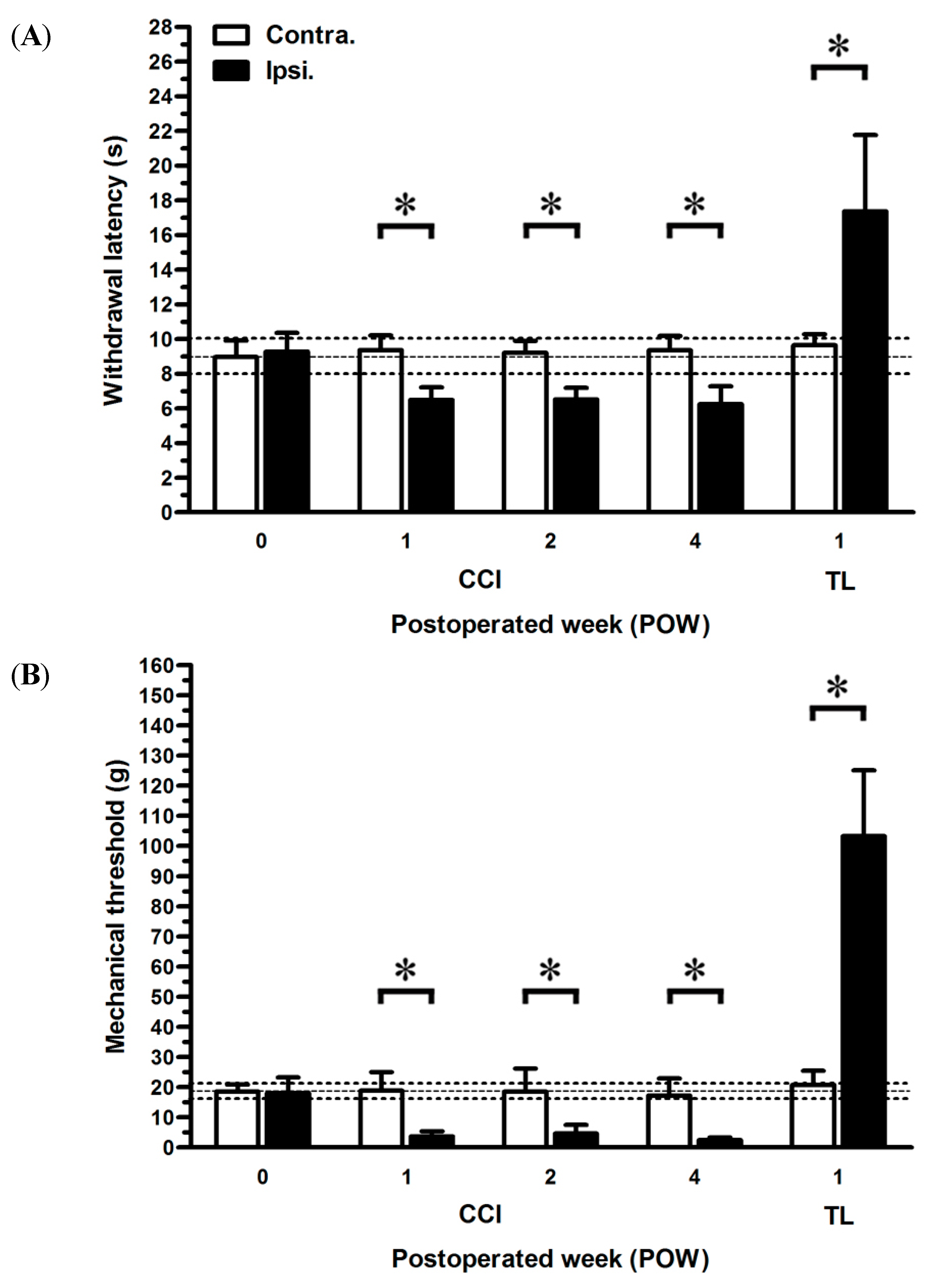
2.1. Neuropathic Pain Behaviors Following CCI

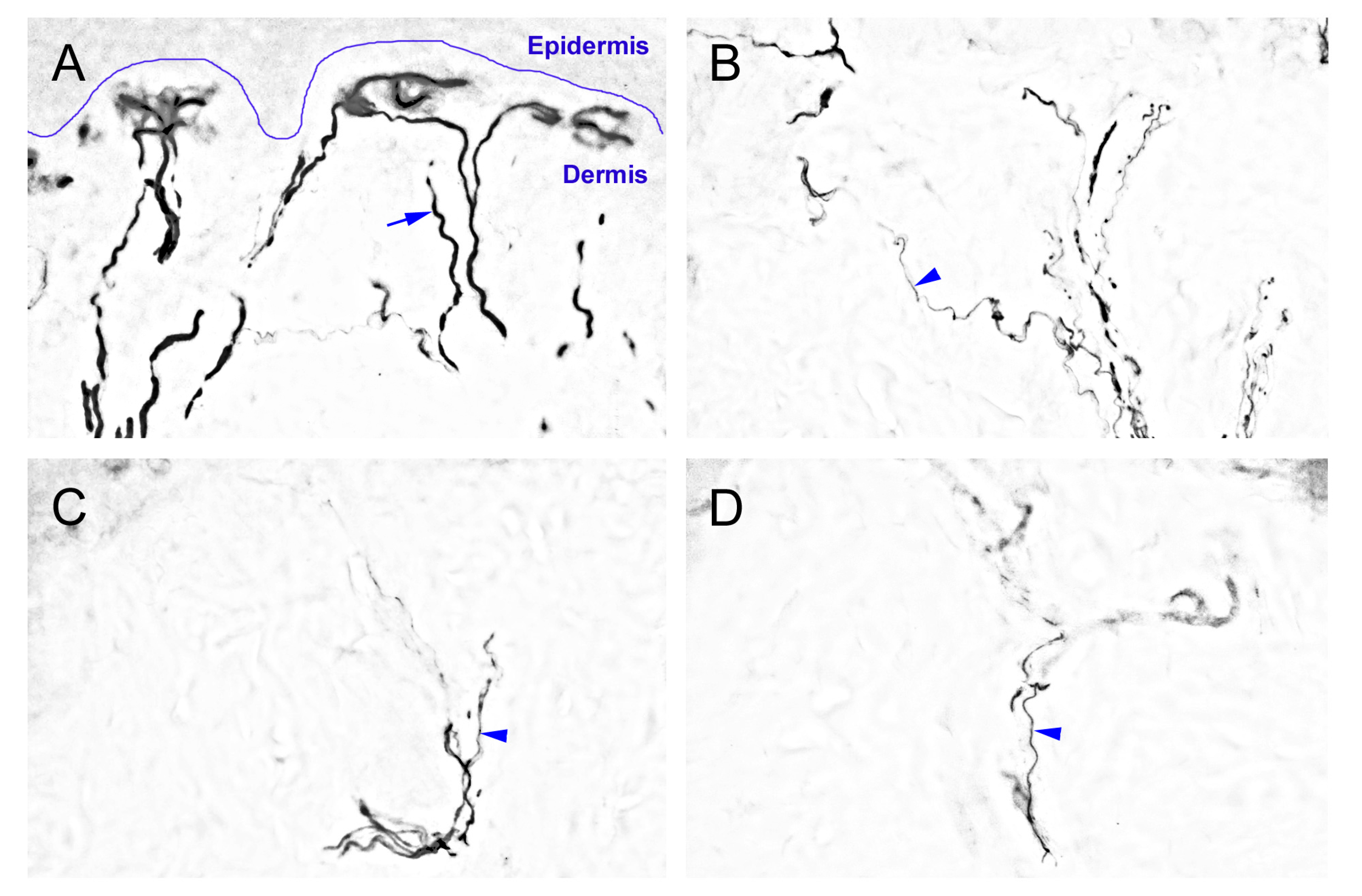
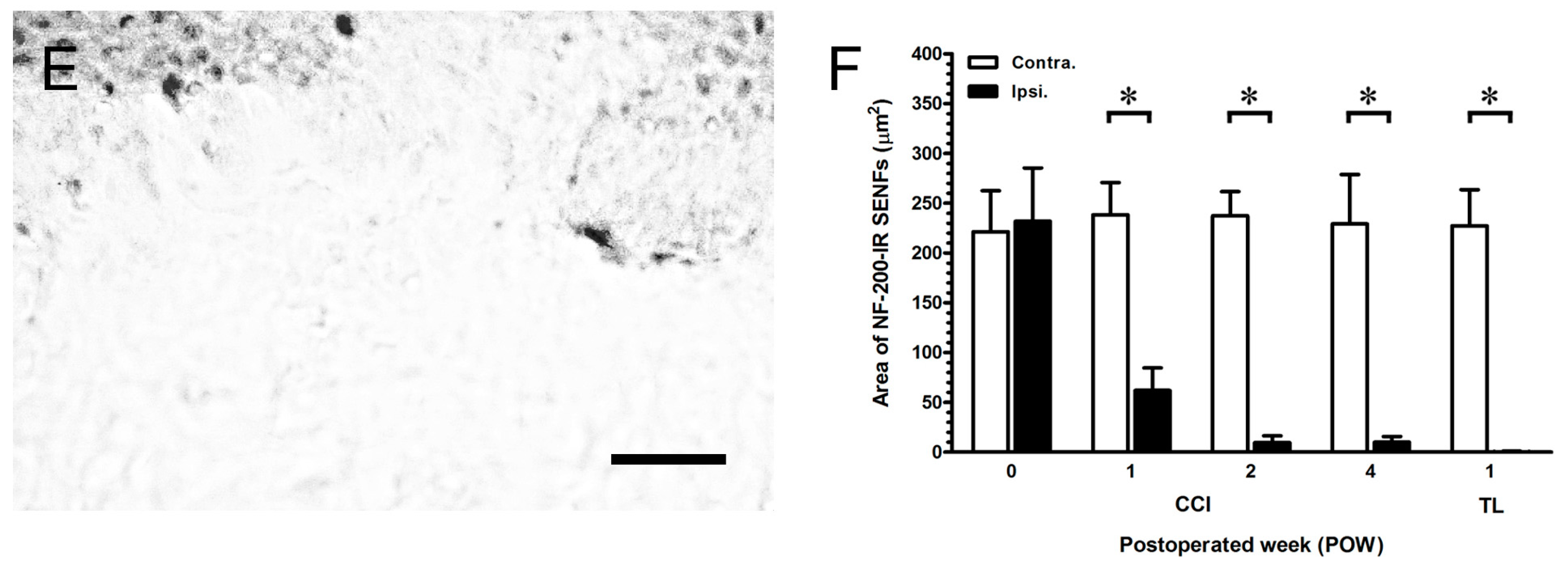
2.2. The Degeneration of Neurofilament 200 (NF-200)-Immunoreactive (IR) SENFs in Dermis after CCI


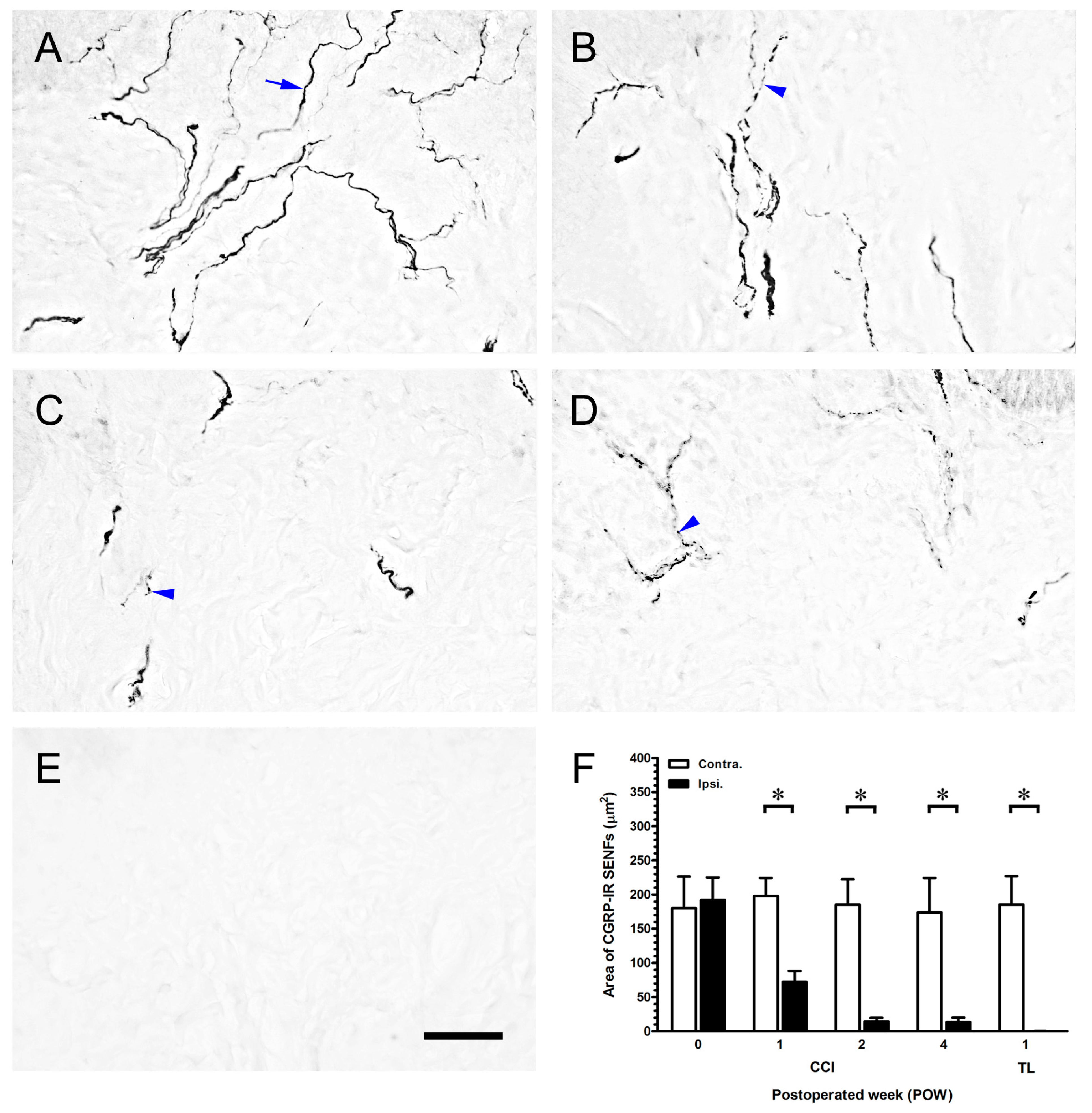
2.3. CCI-Induced the Significant Loss of Dermal Calcitonin Gene-Related Peptide (CGRP)-IR SENFs

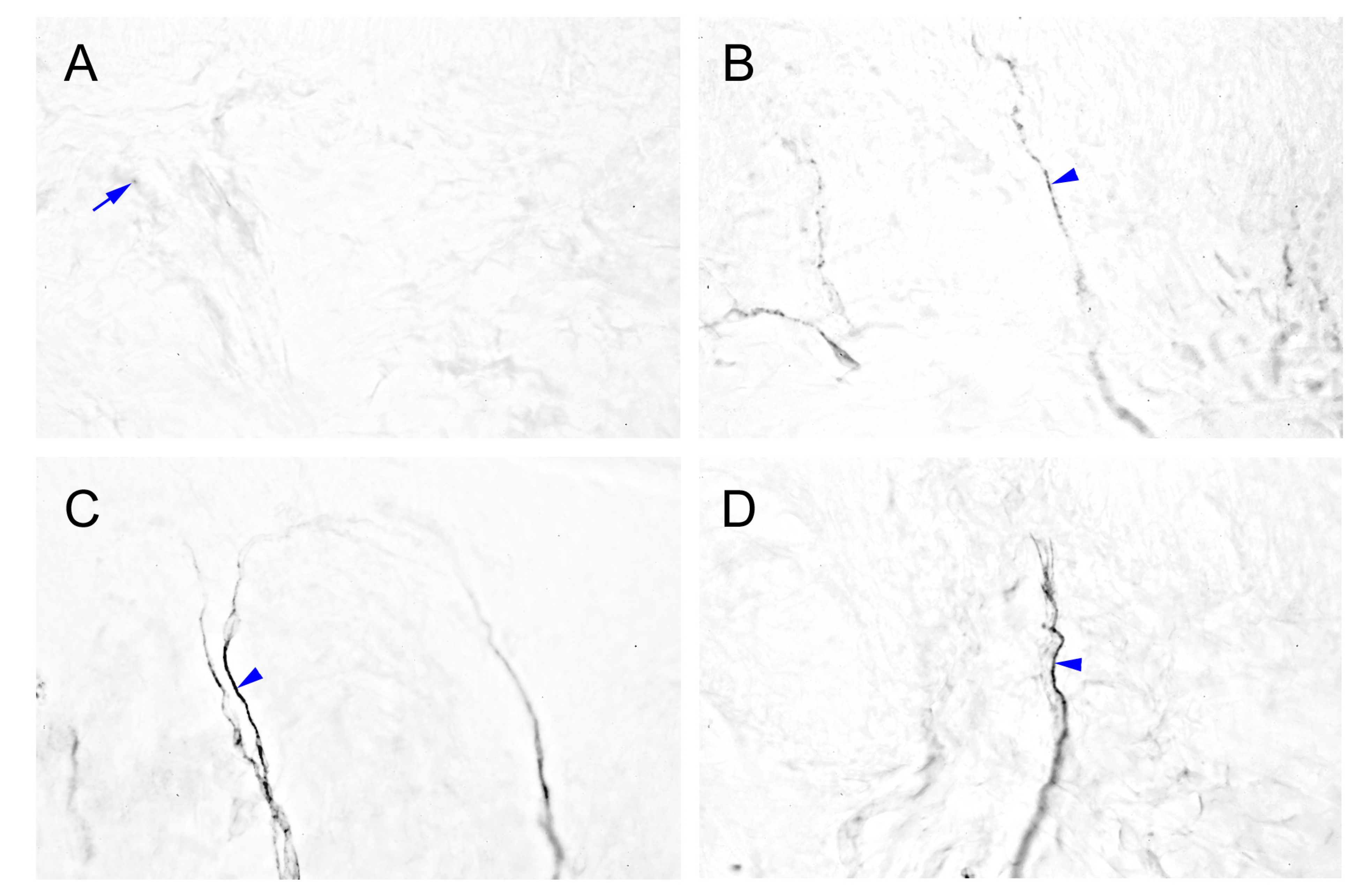
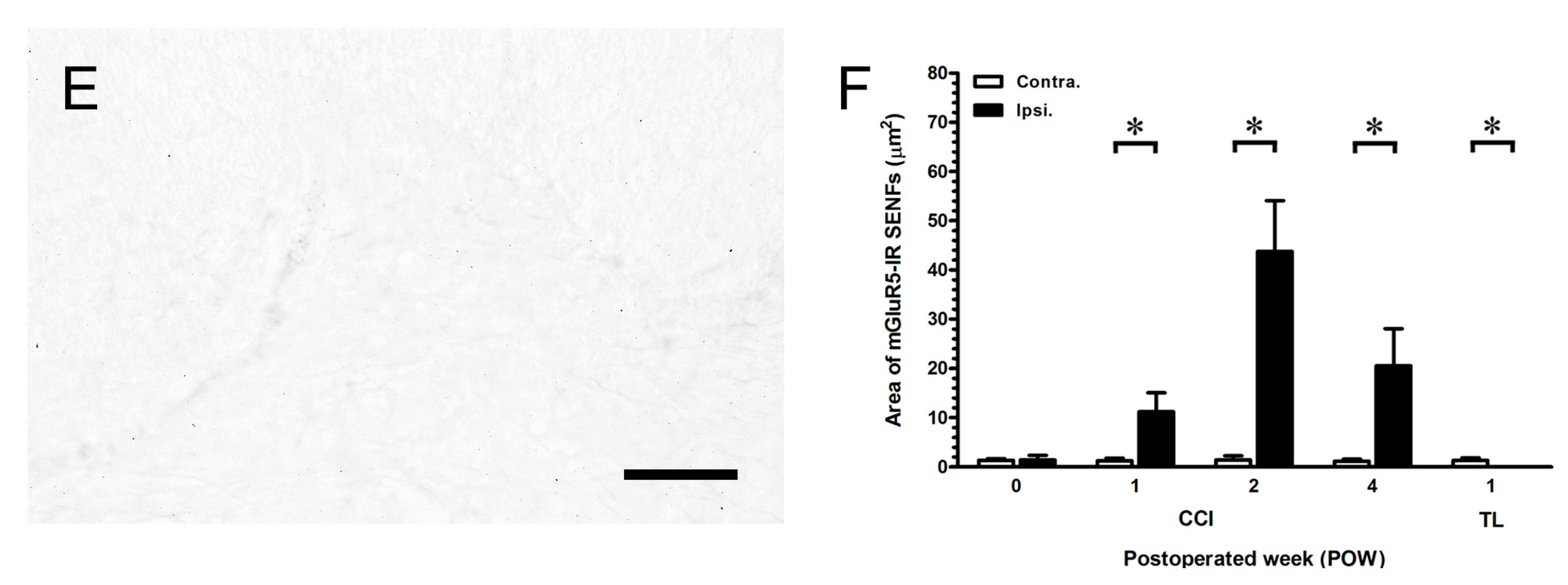
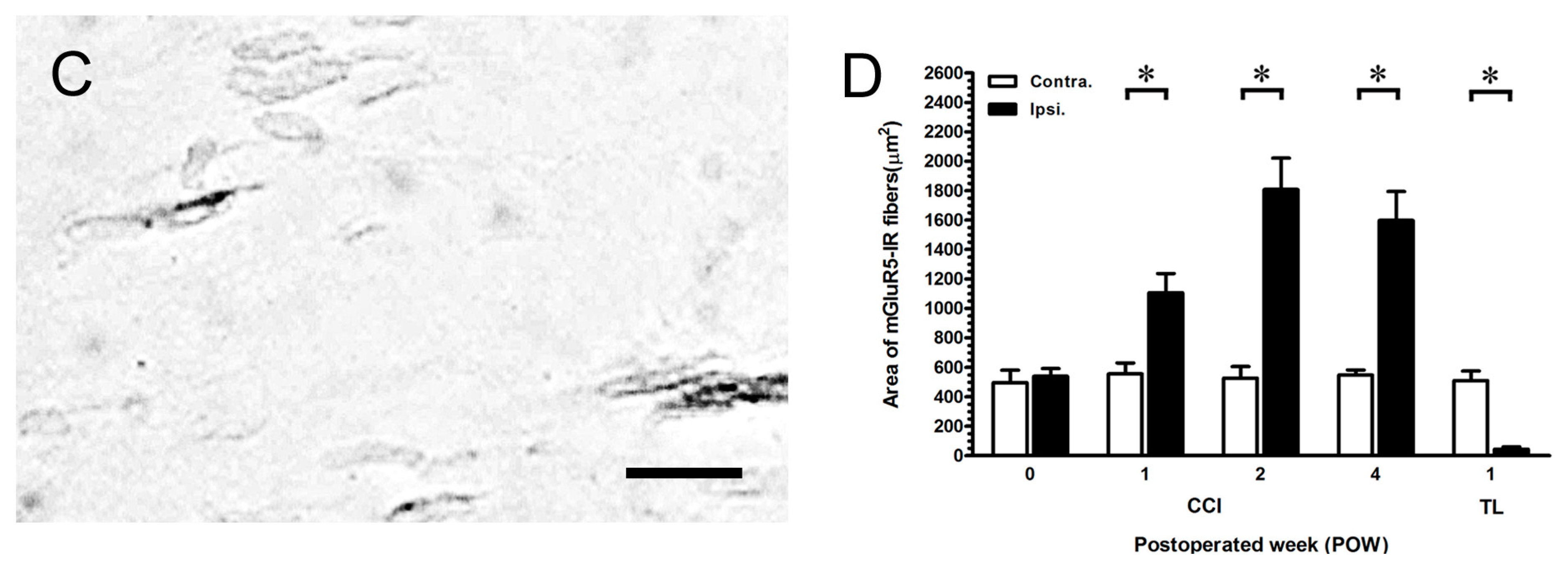
2.4. Intensely Increased mGluR5-IR SENFs in Dermis Following CCI


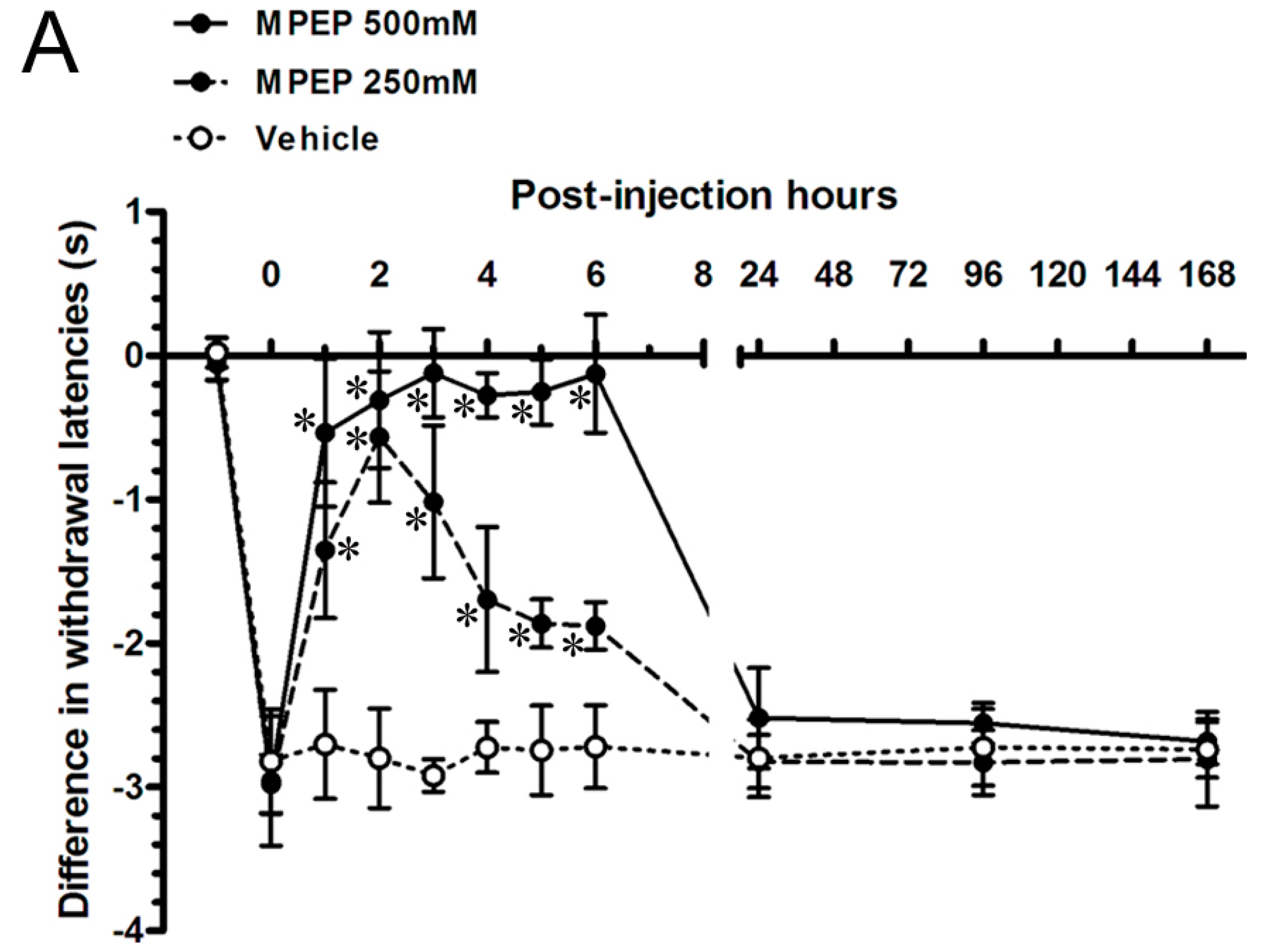
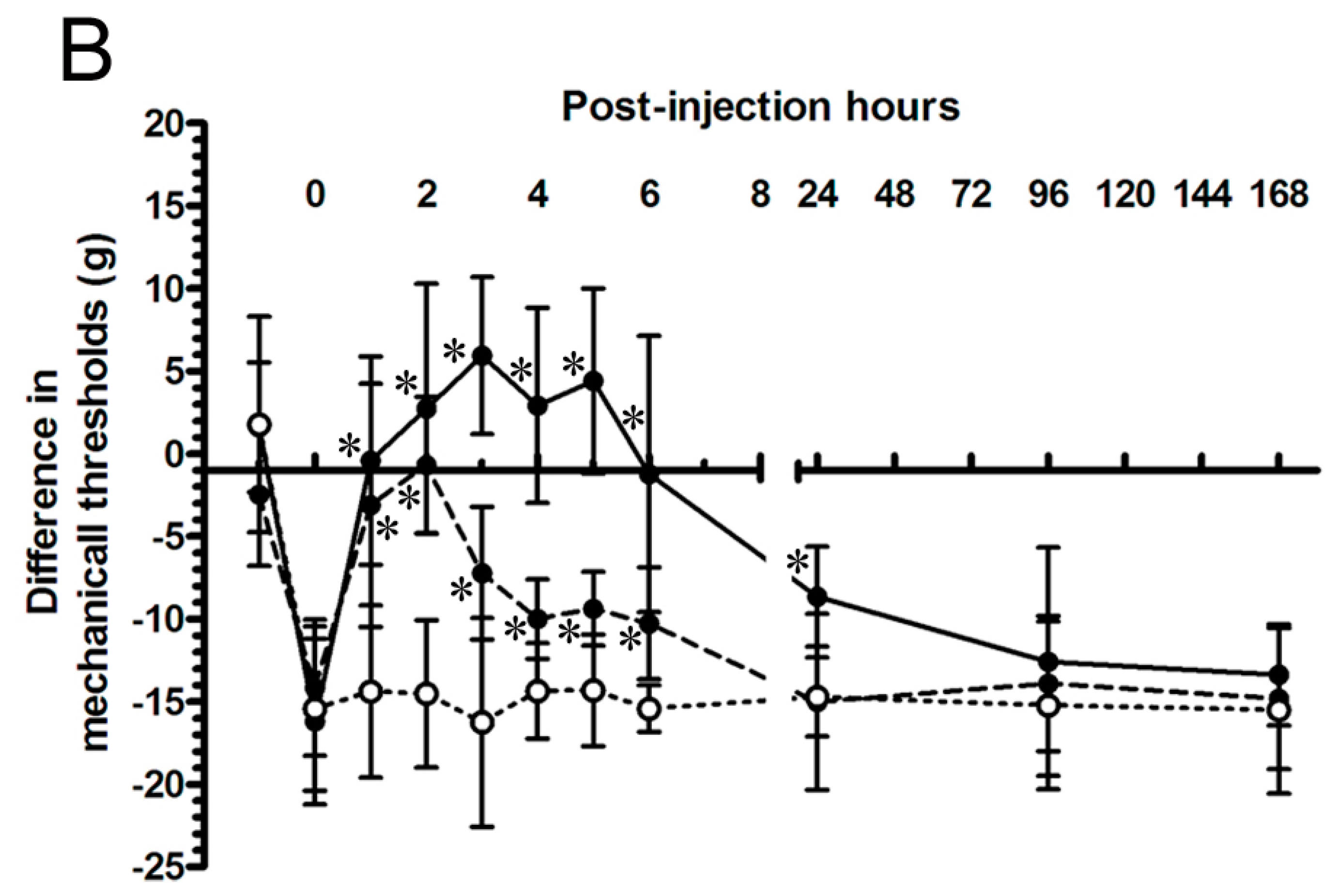
2.5. MPEP Attenuated the Neuropathic Pain Behaviors by Intraplantar Injections
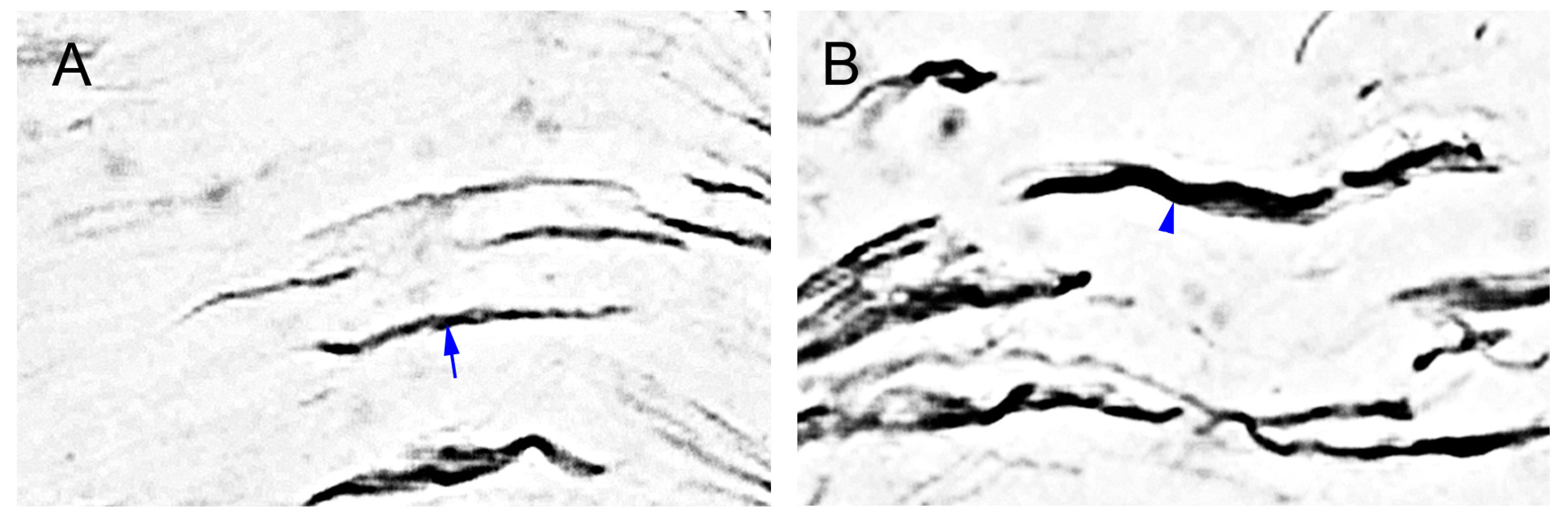
2.6. MGluR5-IR Fibers Increased along the Distal CCI Stumps of Sciatic Nerve




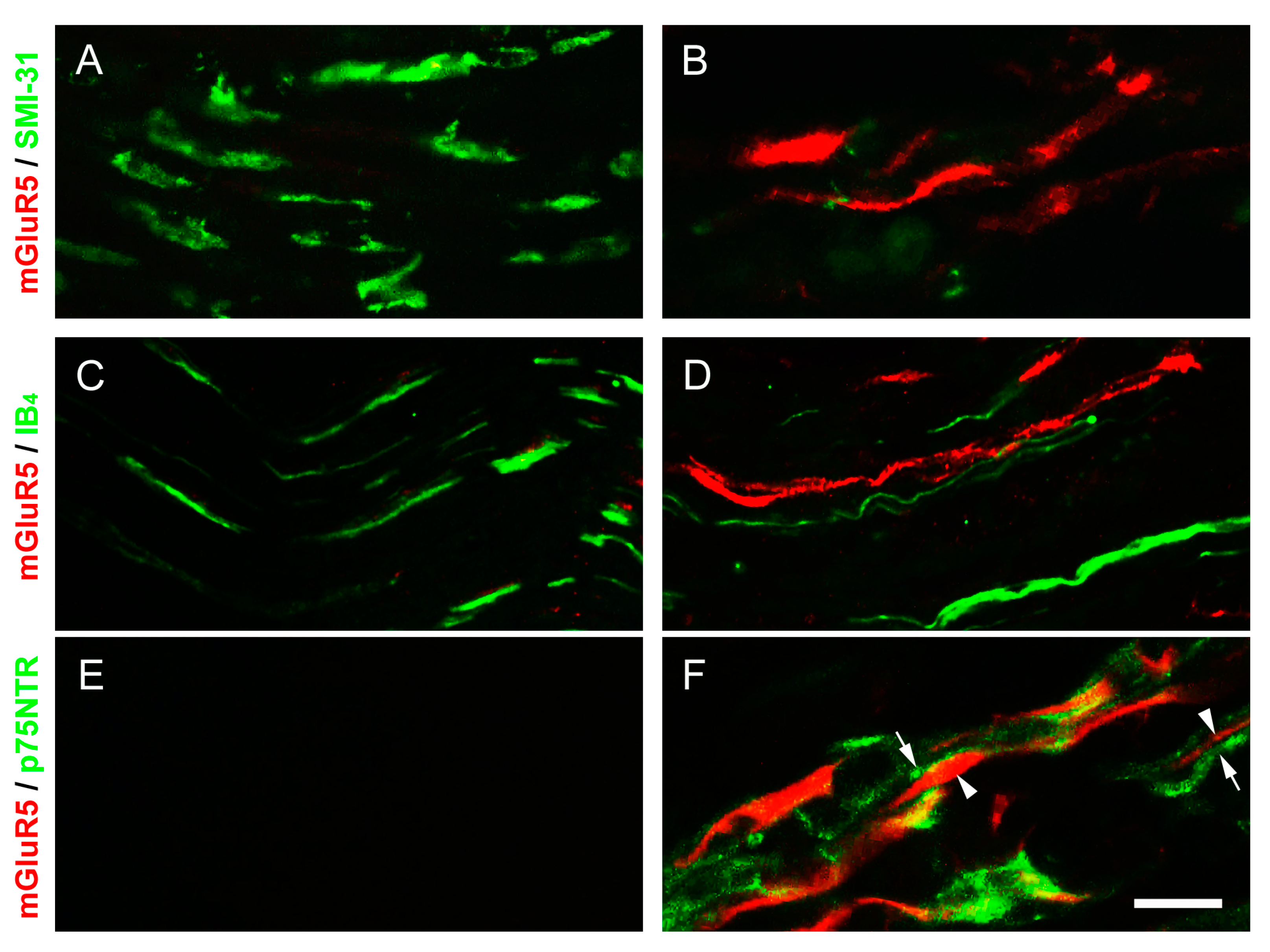
2.7. p75 Neurotrophin Receptor (p75NTR)-IR Reactive Schwann Cells Surrounded the mGluR5-IR Fibers at the Distal CCI Stumps of Sciatic Nerve

3. Discussion
3.1. Different Degrees of Nerve Compression Used to Assess Behaviors
3.2. The Important Relationships between the Peripheral Hypersensitivities and Cutaneous Denervations
3.3. The Efficient Role of mGluR5 in Neuropathic Pain Behaviors
3.4. The mGluR5 Localization in Periphery
4. Experimental Section
4.1. Animals
4.2. Surgeries
4.3. Neuropathic Pain Behaviors
4.3.1. Thermal Hyperalgesia
4.3.2. Mechanical Allodynia
4.4. Immunohistochemistry of Footpad Skin
4.4.1. Procedures
4.4.2. Quantitation
4.5. Pharmacological Intervention
4.5.1. Drugs
4.5.2. Behavioral Assessments
4.6. Immunohistochemistry of Sciatic Nerve
4.6.1. Procedures
4.6.2. Quantitation
4.7. Double-Labeled Immunofluorescence
4.8. Study Designs
4.9. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Basbaum, A.I. Distinct neurochemical features of acute and persistent pain. Proc. Natl. Acad. Sci. USA 1999, 96, 7739–7743. [Google Scholar] [PubMed]
- Jacobs, W.C.; van Tulder, M.; Arts, M.; Rubinstein, S.M.; van Middelkoop, M.; Ostelo, R.; Verhagen, A.; Koes, B.; Peul, W.C. Surgery vs. conservative management of sciatica due to a lumbar herniated disc: A systematic review. Eur. Spine J. 2011, 20, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.J.; Xie, Y.K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 1988, 33, 87–107. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Chung, J.M. An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain 1992, 50, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Seltzer, Z.; Dubner, R.; Shir, Y. A novel behavioral model of neuropathic pain disorders produced in rats by partial sciatic nerve injury. Pain 1990, 43, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Niederberger, E.; Kühlein, H.; Geisslinger, G. Update on the pathobiology of neuropathic pain. Expert Rev. Proteomic 2008, 5, 799–818. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Gautron, M.; Jazat, F.; Mayes, M.; Guilbaud, G. The spectrum of fiber loss in a model of neuropathic pain in the rat: An electron microscopic study. Pain 1991, 47, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Lindenlaub, T.; Sommer, C. Epidermal innervation density after partial sciatic nerve lesion and pain-related behavior in the rat. Acta Neuropathol. 2002, 104, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Bisby, M.A. Calcitonin gene-related peptide, substance P and protein gene product 9.5 immunoreactive axonal fibers in the rat footpad skin following partial sciatic nerve injuries. J. Neurocytol. 2000, 29, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Tseng, T.J.; Chen, C.C.; Hsieh, Y.L.; Hsieh, S.T. Effects of decompression on neuropathic pain behaviors and skin reinnervation in chronic constriction injury. Exp. Neurol. 2007, 204, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Casanova-Molla, J.; Morales, M.; Solà-Valls, N.; Bosch, A.; Calvo, M.; Grau-Junyent, J.M.; Valls-Solé, J. Axonal fluorescence quantitation provides a new approach to assess cutaneous innervation. J. Neurosci. Methods 2011, 200, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Myers, M.I.; Peltier, A.C.; Li, J. Evaluating dermal myelinated nerve fibers in skin biopsy. Muscle Nerve 2013, 47, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Provitera, V.; Nolano, M.; Pagano, A.; Caporaso, G.; Stancanelli, A.; Santoro, L. Myelinated nerve endings in human skin. Muscle Nerve 2007, 35, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Coggeshall, R.E.; Carlton, S.M. Ultrastructural analysis of NMDA, AMPA, and kainate receptors on unmyelinated and myelinated axons in the periphery. J. Comp. Neurol. 1998, 391, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Neugebauer, V. Peripheral metabotropic glutamate receptors: Fight the pain where it hurts. Trends Neurosci. 2001, 24, 550–552. [Google Scholar] [CrossRef] [PubMed]
- Dickenson, A.H.; Chapman, V.; Green, G.M. The pharmacology of excitatory and inhibitory amino acid-mediated events in the transmission and modulation of pain in the spinal cord. Gen. Pharmacol. 1997, 28, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Knöpfel, T.; Kuhn, R.; Allgeier, H. Metabotropic glutamate receptors: Novel targets for drug development. J. Med. Chem. 1995, 38, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- Bhave, G.; Karim, F.; Carlton, S.M.; Gereau, R.W., IV. Peripheral group I metabotropic glutamate receptors modulate nociception in mice. Nat. Neurosci. 2001, 4, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.S.; Sung, K.W.; Lee, S.E.; Shin, H.K. Capsaicin prevents the hyperalgesia induced by peripheral group I mGluRs activation. Neurosci. Lett. 2011, 500, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Neugebauer, V.; Carlton, S.M. Peripheral metabotropic glutamate receptors as drug targets for pain relief. Expert Opin. Ther. Targets 2002, 6, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Walk, D. Role of skin biopsy in the diagnosis of peripheral neuropathic pain. Curr. Pain Headache Rep. 2009, 13, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Urban, M.O. Effect of metabotropic glutamate receptor subtype 5 antagonist, 2-methyl-6-(phenylethynyl)-pyridine on mechanosensory afferents innervating rat hind paw following inflammation. Neurosci. Lett. 2005, 390, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Nicola, D.; Valle-Argos, B.; Suardíaz, M.; Taylor, J.S.; Nieto-Sampedro, M. Role of IL-15 in spinal cord and sciatic nerve after chronic constriction injury: Regulation of macrophage and T-cell infiltration. J. Neurochem. 2008, 107, 1741–1752. [Google Scholar] [CrossRef] [PubMed]
- Gould, H.J., III; Soignier, R.D.; Cho, S.R.; Hernandez, C.; Diamond, I.; Taylor, B.K.; Paul, D. Ranolazine attenuates mechanical allodynia associated with demyelination injury. Pain Med. 2014, 15, 1771–1780. [Google Scholar] [CrossRef] [PubMed]
- Gabay, E.; Tal, M. Pain behavior and nerve electrophysiology in the CCI model of neuropathic pain. Pain 2004, 110, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.; Heckman, H.M.; Myers, R.R. Wallerian degeneration and hyperalgesia after peripheral nerve injury are glutathione-dependent. Pain 1998, 77, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Tseng, T.J.; Hsieh, Y.L.; Ko, M.H.; Hsieh, S.T. Redistribution of voltage-gated sodium channels after nerve decompression contributes to relieve neuropathic pain in chronic constriction injury. Brain Res. 2014, 1589, 15–25. [Google Scholar] [CrossRef]
- Li, L.; Qin, H.; Shi, W.; Gao, G. Local Nogo-66 administration reduces neuropathic pain after sciatic nerve transection in rat. Neurosci. Lett. 2007, 424, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.W.; Tseng, T.J.; Lin, W.M.; Hsieh, S.T. Cutaneous nerve terminal degeneration in painful mononeuropathy. Exp. Neurol. 2001, 170, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Devor, M. Ectopic mechanosensitivity in injured sensory axons arises from the site of spontaneous electrogenesis. Eur. J. Pain 1998, 2, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Devor, M.; Raber, P. Autotomy after nerve injury and its relation to spontaneous discharge originating in nerve-end neuromas. Behav. Neural. Biol. 1983, 37, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Kim, D.W.; Sang Nam, T.; Se Paik, K.; Leem, J.W. Peripheral glutamate receptors contribute to mechanical hyperalgesia in a neuropathic pain model of the rat. Neuroscience 2004, 128, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Chien, H.F.; Tseng, T.J.; Lin, W.M.; Yang, C.C.; Chang, Y.C.; Chen, R.C.; Hsieh, S.T. Quantitative pathology of cutaneous nerve terminal degeneration in the human skin. Acta Neuropathol. 2001, 102, 455–461. [Google Scholar] [PubMed]
- Sommer, C. Skin biopsy as a diagnostic tool. Curr. Opin. Neurol. 2008, 21, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Lauria, G.; Cornblath, D.R.; Johansson, O.; McArthur, J.C.; Mellgren, S.I.; Nolano, M.; Rosenberg, N.; Sommer, C. EFNS guidelines on the use of skin biopsy in the diagnosis of peripheral neuropathy: European Federation of Neurological Societies. Eur. J. Neurol. 2005, 12, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Lauria, G.; Lombardi, R. Skin biopsy in painful and immune-mediated neuropathies. J. Peripher. Nerv. Syst. 2012, 17, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Schüning, J.; Scherens, A.; Haussleiter, I.S.; Schwenkreis, P.; Krumova, E.K.; Richter, H.; Maier, C. Sensory changes and loss of intraepidermal nerve fibers in painful unilateral nerve injury. Clin. J. Pain 2009, 25, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Lauria, G.; Cazzato, D.; Porretta-Serapiglia, C.; Casanova-Molla, J.; Taiana, M.; Penza, P.; Lombardi, R.; Faber, C.G.; Merkies, I.S. Morphometry of dermal nerve fibers in human skin. Neurology 2011, 77, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Lawson, S.N.; Waddell, P.J. Soma neurofilament immunoreactivity is related to cell size and fibre conduction velocity in rat primary sensory neurons. J. Physiol. 1991, 435, 41–63. [Google Scholar] [CrossRef] [PubMed]
- Ruscheweyh, R.; Forsthuber, L.; Schoffnegger, D.; Sandkühler, J. Modification of classical neurochemical markers in identified primary afferent neurons with Aβ-, Aδ-, and C-fibers after chronic constriction injury in mice. J. Comp. Neurol. 2007, 502, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Peleshok, J.C.; Ribeiro-da-Silva, A. Delayed reinnervation by nonpeptidergic nociceptive afferents of the glabrous skin of the rat hindpaw in a neuropathic pain model. J. Comp. Neurol. 2011, 519, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Hudson, L.J.; Bevan, S.; McNair, K.; Gentry, C.; Fox, A.; Kuhn, R.; Winter, J. Metabotropic glutamate receptor 5 upregulation in A-fibers after spinal nerve injury: 2-Methyl-6-(phenylethynyl)-pyridine (MPEP) reverses the induced thermal hyperalgesia. J. Neurosci. 2002, 22, 2660–2668. [Google Scholar] [PubMed]
- Urban, M.O.; Hama, A.T.; Bradbury, M.; Anderson, J.; Varney, M.A.; Bristow, L. Role of metabotropic glutamate receptor subtype 5 (mGluR5) in the maintenance of cold hypersensitivity following a peripheral mononeuropathy in the rat. Neuropharmacology 2003, 44, 983–993. [Google Scholar] [CrossRef] [PubMed]
- De Novellis, V.; Siniscalco, D.; Galderisi, U.; Fuccio, C.; Nolano, M.; Santoro, L.; Cascino, A.; Roth, K.A.; Rossi, F.; Maione, S. Blockade of glutamate mGlu5 receptors in a rat model of neuropathic pain prevents early over-expression of pro-apoptotic genes and morphological changes in dorsal horn lamina II. Neuropharmacology 2004, 46, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Hama, A.T. Acute activation of the spinal cord metabotropic glutamate subtype-5 receptor leads to cold hypersensitivity in the rat. Neuropharmacology 2003, 44, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Fisher, K.; Lefebvre, C.; Coderre, T.J. Antinociceptive effects following intrathecal pretreatment with selective metabotropic glutamate receptor compounds in a rat model of neuropathic pain. Pharmacol. Biochem. Behav. 2002, 73, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Osikowicz, M.; Mika, J.; Makuch, W.; Przewlocka, B. Glutamate receptor ligands attenuate allodynia and hyperalgesia and potentiate morphine effects in a mouse model of neuropathic pain. Pain 2008, 139, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.Z.; Hsieh, G.; Ei-Kouhen, O.; Wilson, S.G.; Mikusa, J.P.; Hollingsworth, P.R.; Chang, R.; Moreland, R.B.; Brioni, J.; Decker, M.W.; et al. Role of central and peripheral mGluR5 receptors in post-operative pain in rats. Pain 2005, 114, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Dogrul, A.; Ossipov, M.H.; Lai, J.; Malan, T.P., Jr.; Porreca, F. Peripheral and spinal antihyperalgesic activity of SIB-1757, a metabotropic glutamate receptor (mGLUR(5)) antagonist, in experimental neuropathic pain in rats. Neurosci. Lett. 2000, 292, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Carlton, S.M.; Dougherty, P.M.; Pover, C.M.; Coggeshall, R.E. Neuroma formation and numbers of axons in a rat model of experimental peripheral neuropathy. Neurosci. Lett. 1991, 131, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Coggeshall, R.E.; Dougherty, P.M.; Pover, C.M.; Carlton, S.M. Is large myelinated fiber loss associated with hyperalgesia in a model of experimental peripheral neuropathy in the rat? Pain 1993, 52, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Guilbaud, G.; Gautron, M.; Jazat, F.; Ratinahirana, H.; Hassig, R.; Hauw, J.J. Time course of degeneration and regeneration of myelinated nerve fibres following chronic loose ligatures of the rat sciatic nerve: Can nerve lesions be linked to the abnormal pain-related behaviours? Pain 1993, 53, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Gautron, M.; Jazat, F.; Ratinahirana, H.; Hauw, J.J.; Guilbaud, G. Alterations in myelinated fibres in the sciatic nerve of rats after constriction: Possible relationships between the presence of abnormal small myelinated fibres and pain-related behavior. Neurosci. Lett. 1990, 111, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Munger, B.L.; Bennett, G.J.; Kajander, K.C. An experimental painful peripheral neuropathy due to nerve constriction. I. Axonal pathology in the sciatic nerve. Exp. Neurol. 1992, 118, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, C.S.; Sherman, D.L.; Fleetwood-Walker, S.M.; Cottrell, D.F.; Tait, S.; Garry, E.M.; Wallace, V.C.; Ure, J.; Griffiths, I.R.; Smith, A.; et al. Peripheral demyelination and neuropathic pain behavior in periaxin-deficient mice. Neuron 2000, 26, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.L.; Xie, Z.L.; Wu, Y.W.; Duan, W.R.; Xie, Y.K. Early demyelination of primary A-fibers induces a rapid-onset of neuropathic pain in rat. Neuroscience 2012, 200, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Campana, W.M.; Li, X.; Shubayev, V.I.; Angert, M.; Cai, K.; Myers, R.R. Erythropoietin reduces Schwann cell TNF-α, Wallerian degeneration and pain-related behaviors after peripheral nerve injury. Eur. J. Neurosci. 2006, 23, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Shamash, S.; Reichert, F.; Rotshenker, S. The cytokine network of Wallerian degeneration: Tumor necrosis factor-α, interleukin-1α, and interleukin-1β. J. Neurosci. 2002, 22, 3052–3060. [Google Scholar] [PubMed]
- IASP Committee. Ethical standards for investigations of experimental pain in animals. Pain 1980, 9, 141–143. [Google Scholar]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef] [PubMed]
- Gasparini, F.; Lingenhöhl, K.; Stoehr, N.; Flor, P.J.; Heinrich, M.; Vranesic, I.; Biollaz, M.; Allgeier, H.; Heckendorn, R.; Urwyler, S.; et al. 2-Methyl-6-(phenylethynyl)-pyridine (MPEP), a potent, selective and systemically active mGlu5 receptor antagonist. Neuropharmacology 1999, 38, 1493–1503. [Google Scholar] [CrossRef] [PubMed]
- Sakurada, T.; Mizoguchi, H.; Kuwahata, H.; Katsuyama, S.; Komatsu, T.; Morrone, L.A.; Corasaniti, M.T.; Bagetta, G.; Sakurada, S. Intraplantar injection of bergamot essential oil induces peripheral antinociception mediated by opioid mechanism. Pharmacol. Biochem. Behav. 2011, 97, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Thornton, M.R.; Mantovani, C.; Birchall, M.A.; Terenghi, G. Quantification of N-CAM and N-cadherin expression in axotomized and crushed rat sciatic nerve. J. Anat. 2005, 206, 69–78. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, M.-H.; Hsieh, Y.-L.; Hsieh, S.-T.; Tseng, T.-J. Nerve Demyelination Increases Metabotropic Glutamate Receptor Subtype 5 Expression in Peripheral Painful Mononeuropathy. Int. J. Mol. Sci. 2015, 16, 4642-4665. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16034642
Ko M-H, Hsieh Y-L, Hsieh S-T, Tseng T-J. Nerve Demyelination Increases Metabotropic Glutamate Receptor Subtype 5 Expression in Peripheral Painful Mononeuropathy. International Journal of Molecular Sciences. 2015; 16(3):4642-4665. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16034642
Chicago/Turabian StyleKo, Miau-Hwa, Yu-Lin Hsieh, Sung-Tsang Hsieh, and To-Jung Tseng. 2015. "Nerve Demyelination Increases Metabotropic Glutamate Receptor Subtype 5 Expression in Peripheral Painful Mononeuropathy" International Journal of Molecular Sciences 16, no. 3: 4642-4665. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16034642