
The Wnt11 Signaling Pathway in Potential Cellular EMT and Osteochondral Differentiation Progression in Nephrolithiasis Formation

 ,
,
Abstract
:1. Introduction
2. Results
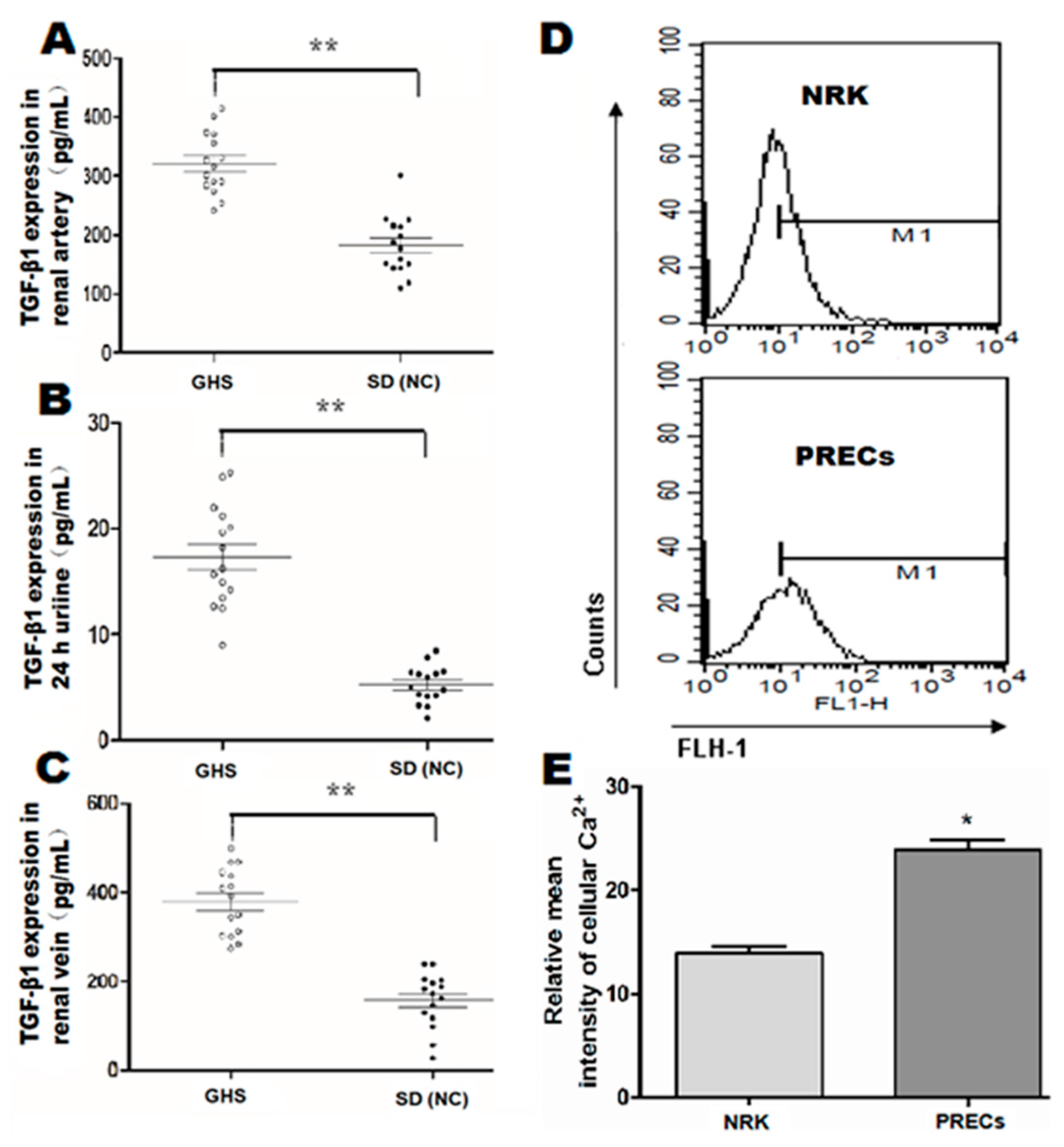
2.1. Basal Levels of TGF-β1 in GHS Rats and Intracellular Ca2+ Concentration in Isolated PRECs

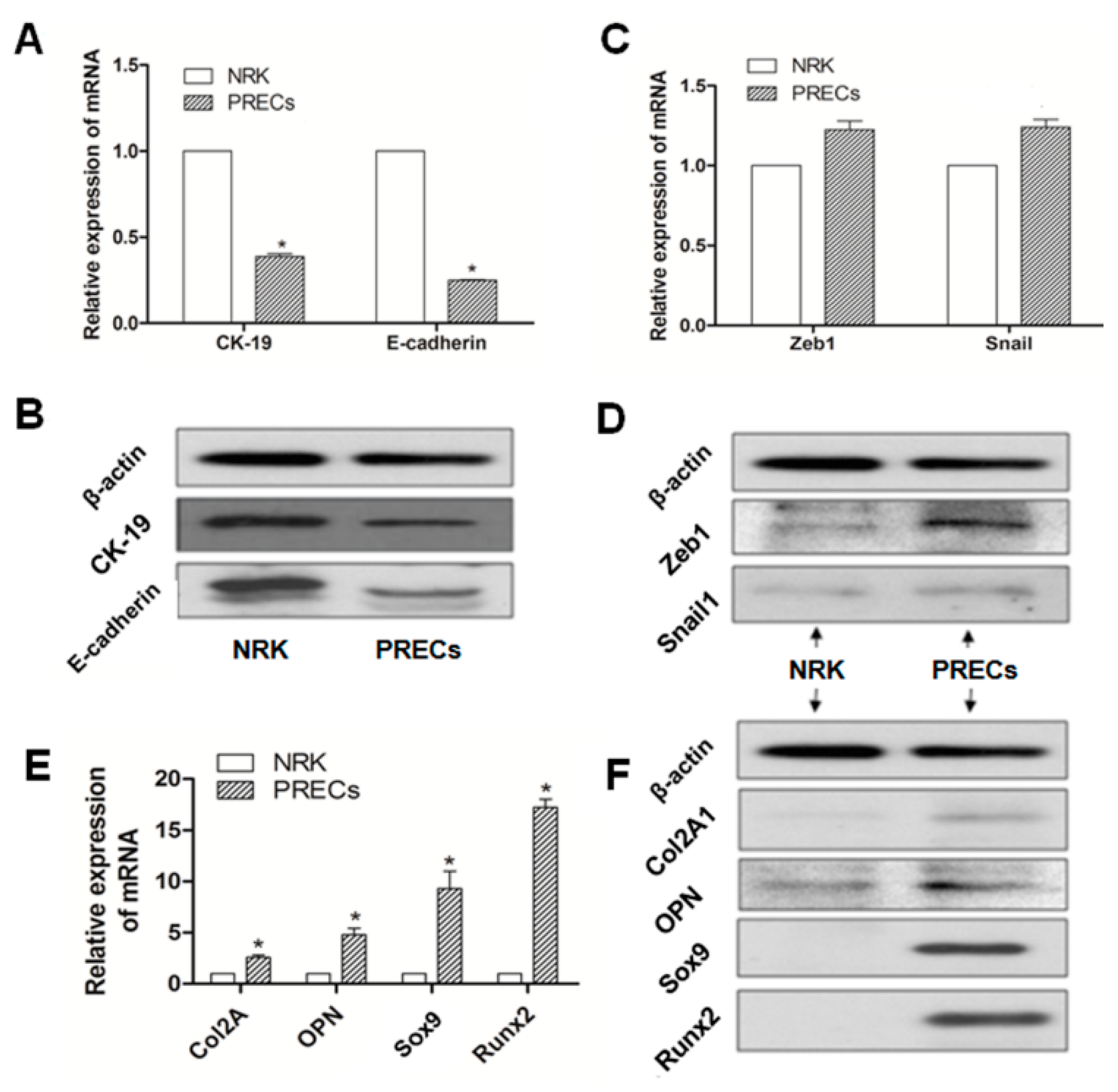
2.2. Different Expression of Specific Bio-Markers in EMT and Osteochondral Differentiation between PRECs and NRK Cells

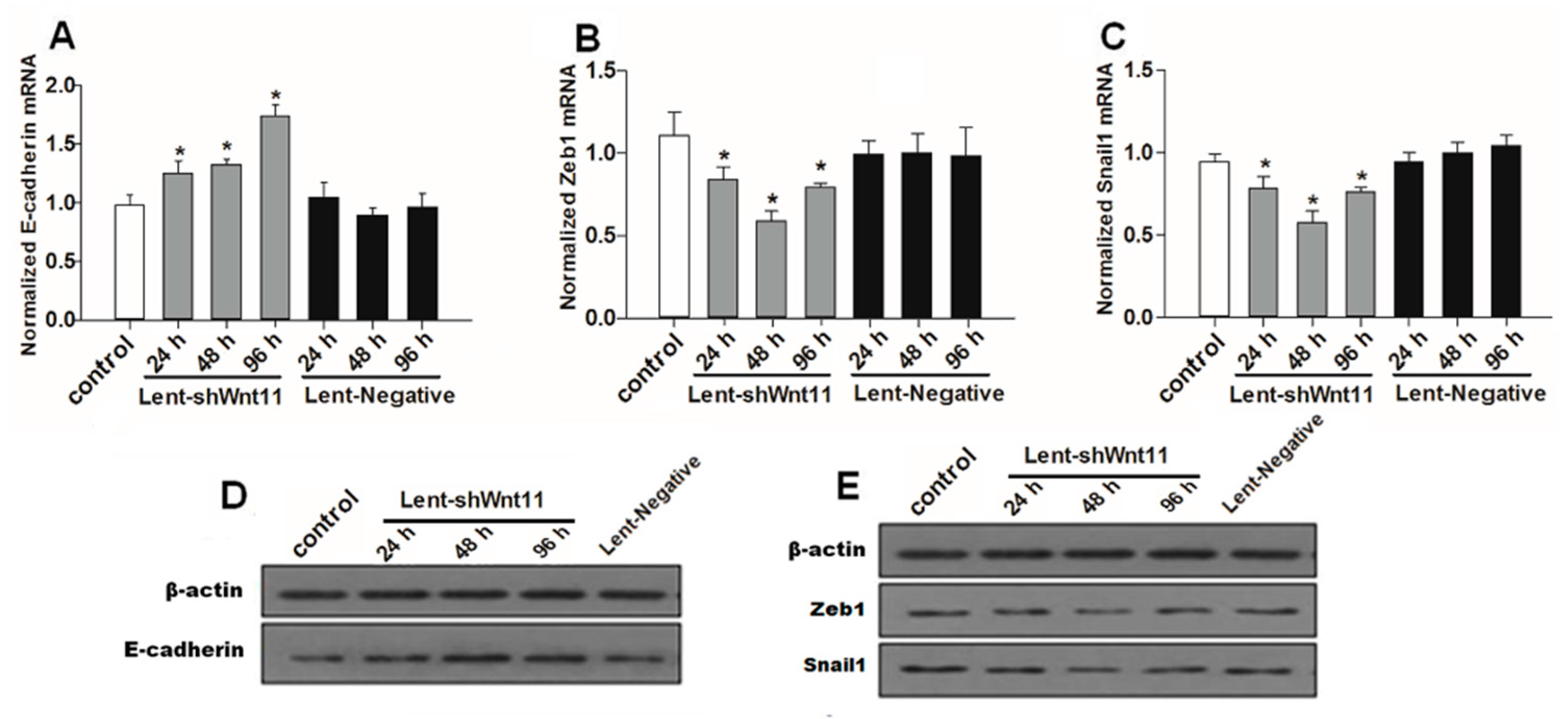
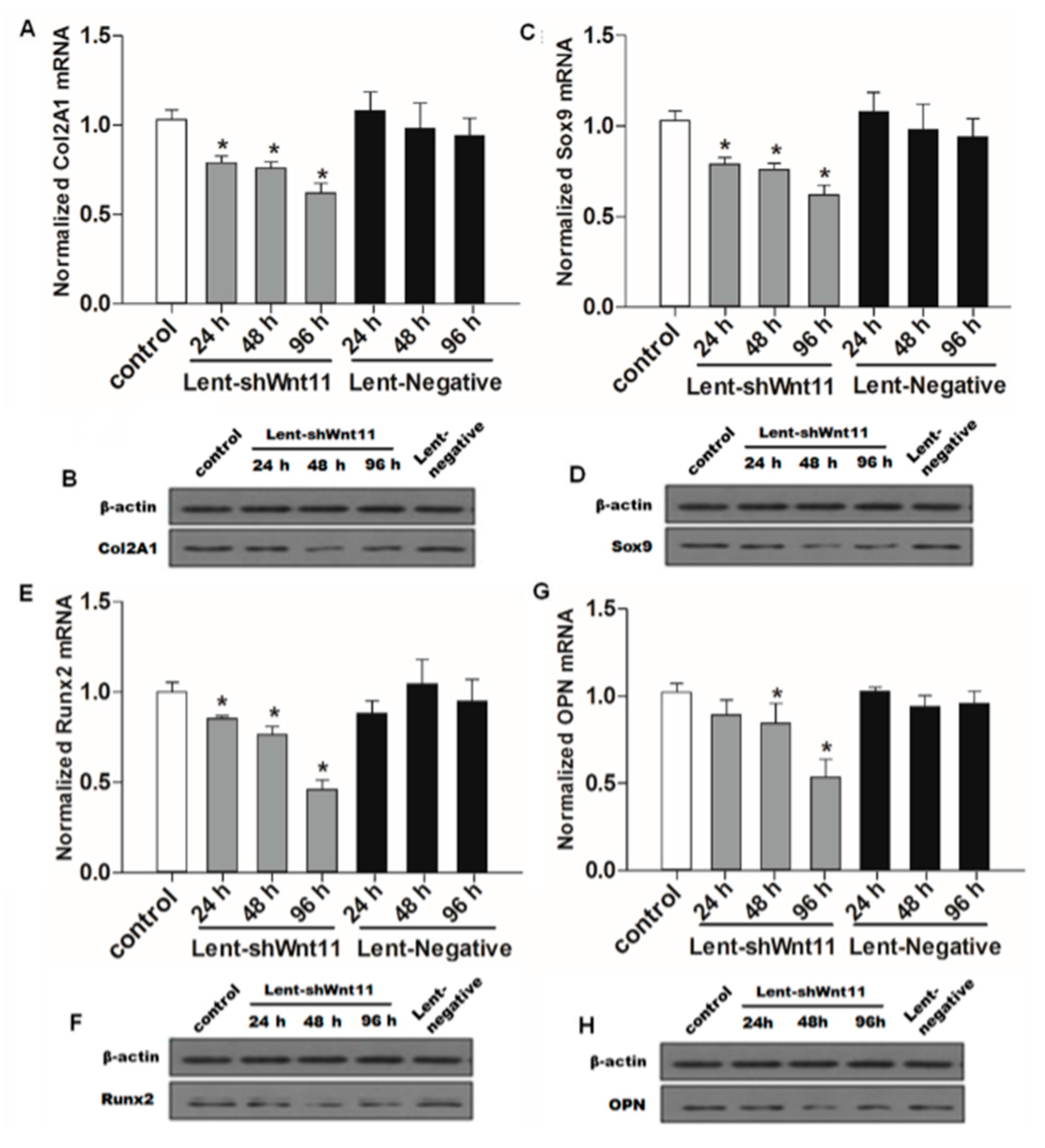
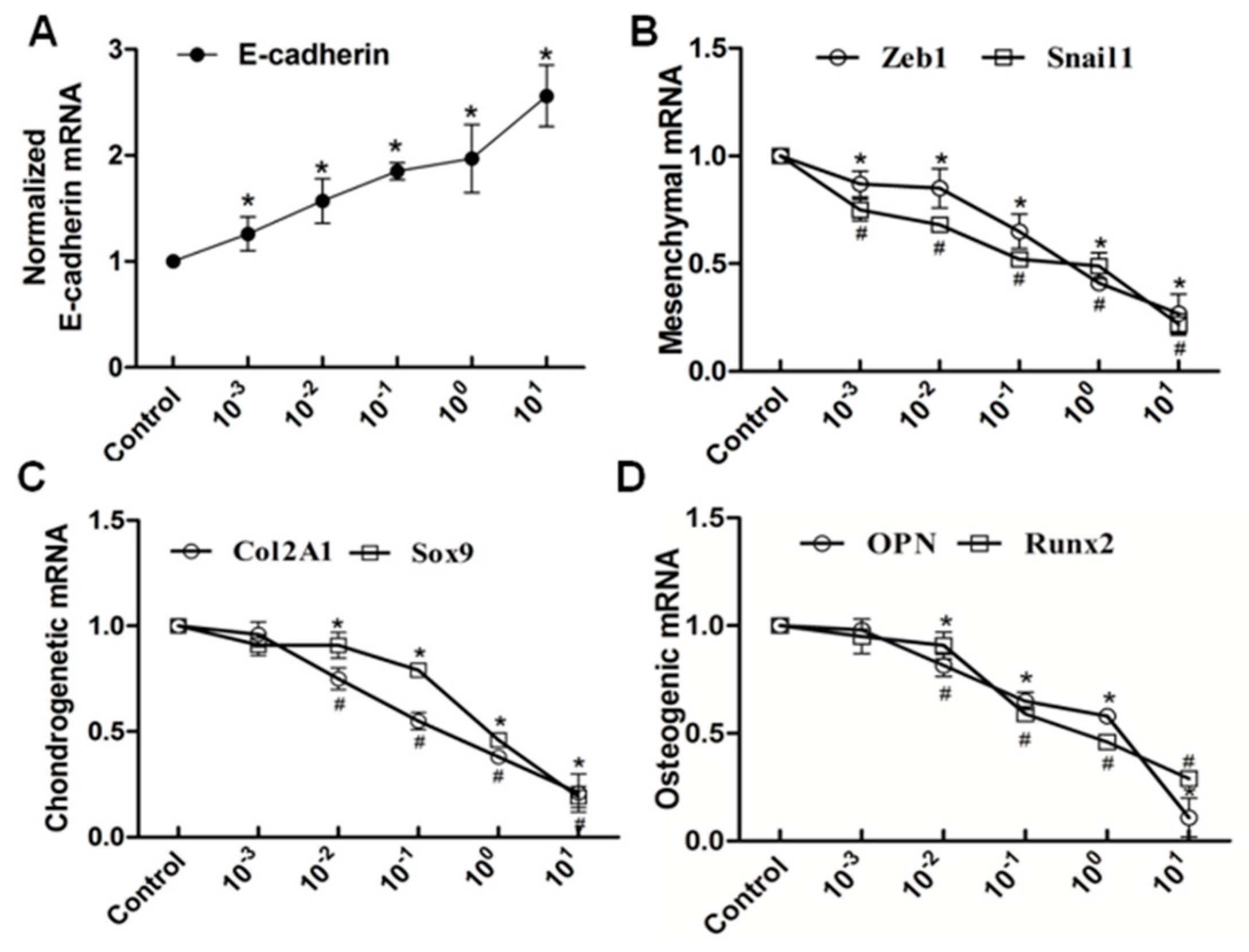
2.3. Wnt11 Knockdown Attenuated the Expression of Osteogenic/Chondrogenetic Factors and Reversed the EMT Process in PRECs



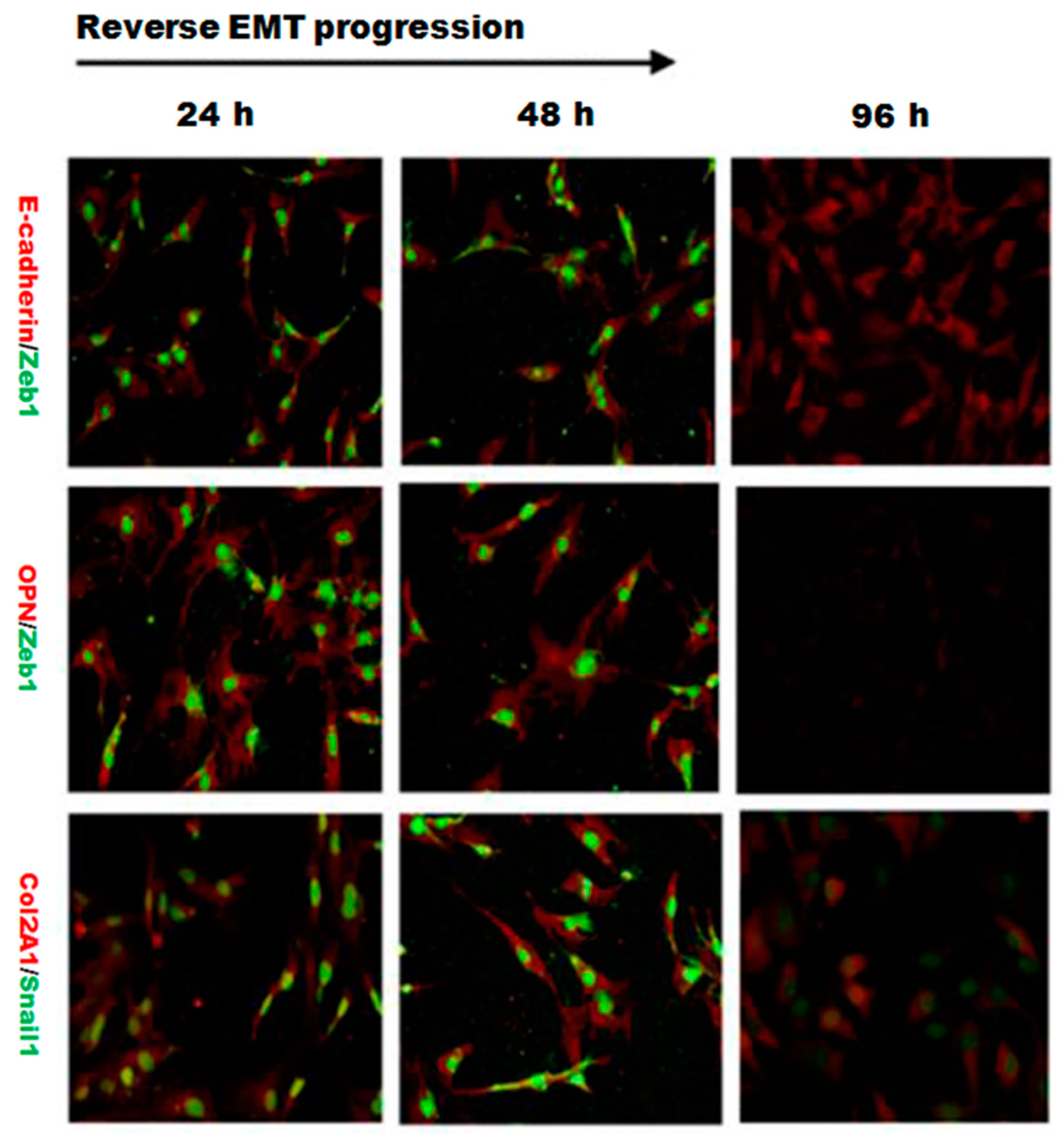
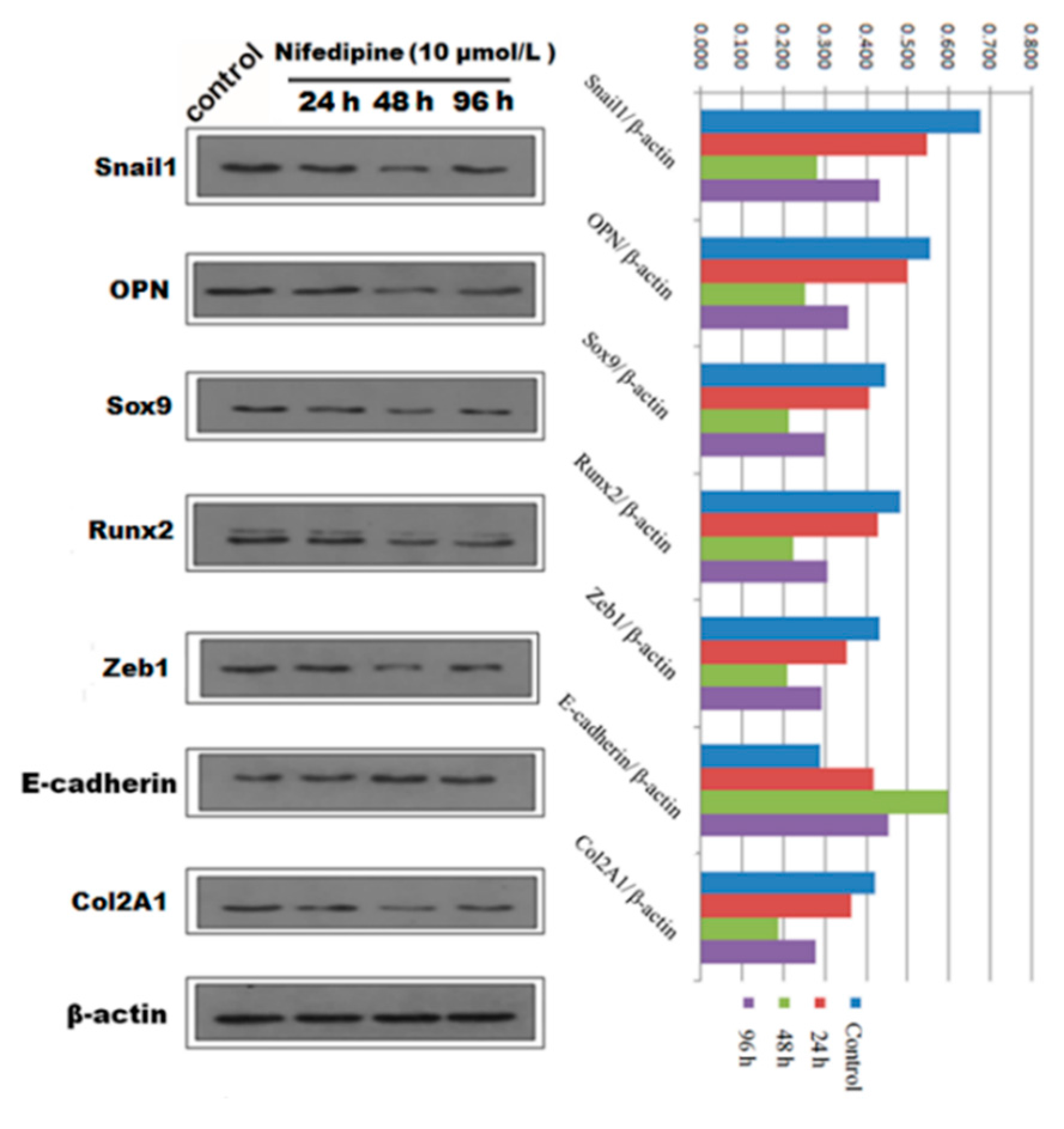
2.4. EMT Process Was Abrogated by L-Type Calcium Channel Blocker Nifedipine in PRECs


2.5. The EMT Process Was Inhibited by SiCav1.2 in PRECs
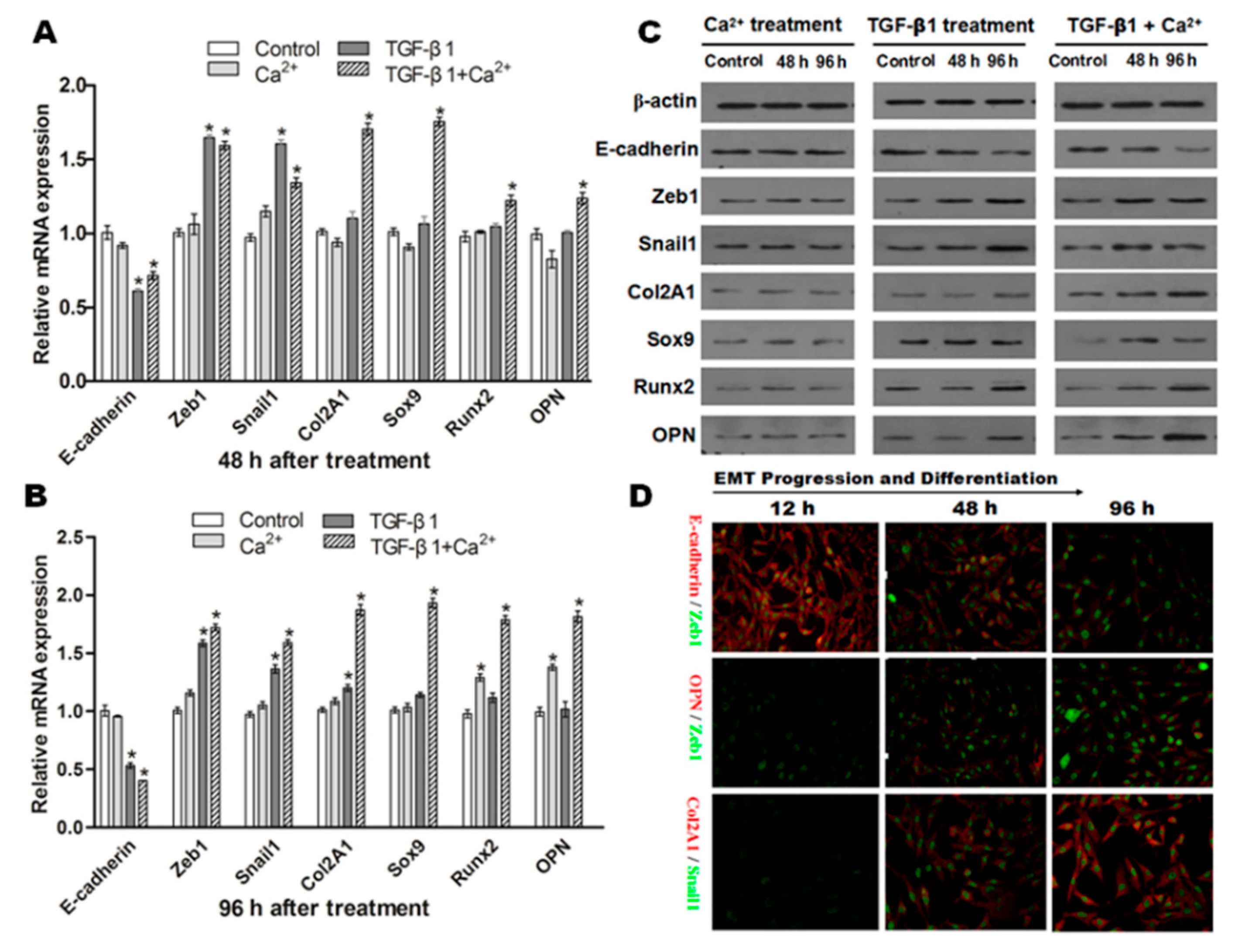
2.6. Effects of Incubation of Ca2+ and/or TGF-β1 in NRK Cells

3. Discussion
4. Materials and Methods
4.1. Animals and Cells Preparation
4.2. Enzyme-Linked Immunosorbent Assay
4.3. Evaluation of Intracellular Ca2+ by Flow Cytometry Assay
4.4. Real-Time RT PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| E-cadherin | GAGGCCAAGCAGCAGTACATT | CGATACGTGATCTTCTGTTCCA |
| CK19 | TAATGGCGAGCTGGAGGTGA | TCGATCTGTAGGACTATCTTGGAG |
| Zeb1 | CCGCAGGGTTACTCTTGTGT | TTGGCACTTGGTGGGACTAC |
| Snail1 | CGGAAGCCCAACTATAGCGA | AGAGTCCCAGATGAGGGTGG |
| Col2A1 | GAAGAGCGGAGACTACTGGATTG | CTGGACGTTAGCGGTGTTGGGAG |
| Sox9 | CACCAGAACTCCGGCTCCTA | TGTGGGTCTGCGGGATGGAA |
| Runx2 | AGCCACCTTCACTTACACCC | CCATTGGGAACTGATAGGAC |
| OPN | GGTTTGCTTTTGCCTGTTCG | GTGGCTACAGCATCTGAGTGTTTG |
| OCN | GTGCAGACCTAGCAGACACCAT | TTCACCACCTTACTGCCCTC |
| β-actin | CACGATGGAGGGGCCGGACTCATC | TAAAGACCTCTATGCCAACACAGT |
4.5. Western Blot Analysis
4.6. Immunofluorescence Staining
4.7. shRNA-Mediated Gene Knockdown
4.8. Treatment of Nifedipine in PRECs and Incubation of Ca2+ (Calcium Chloride) and/or TGF-β1 in NRK Cells
4.9. Orai1 Gene and Cav1.2 Subunit of L-Type Ca2+ Channel Knockdown by siRNA
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Worcester, E.M.; Coe, F.L. Does idiopathic hypercalciuria trigger calcium-sensing receptor-mediated protection from urinary supersaturation? J. Am. Soc. Nephrol. 2009, 20, 1657–1659. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.J.; Bai, S.; Karnauskas, A.J.; Bushinsky, D.A.; Favus, M.J. Regulation of renal calcium receptor gene expression by 1,25-dihydroxyvitamin D3 in genetic hypercalciuric stone-forming rats. J. Am. Soc. Nephrol. 2005, 16, 1300–1308. [Google Scholar] [CrossRef] [PubMed]
- Walker, V.; Cook, P.; Griffin, D.G. Male hypercalciuric stone formers with low renal calcium reabsorption. J. Clin. Pathol. 2014, 67, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Worcester, E.M.; Bergsland, K.J.; Gillen, D.L.; Coe, F.L. Evidence for increased renal tubule and parathyroid gland sensitivity to serum calcium in human idiopathic hypercalciuria. Am. J. Physiol. Ren. Physiol. 2013, 305, F853–F860. [Google Scholar] [CrossRef] [PubMed]
- Yoon, V.; Adams-Huet, B.; Sakhaee, K.; Maalouf, N.M. Hyperinsulinemia and urinary calcium excretion in calcium stone formers with idiopathic hypercalciuria. J. Clin. Endocrinol. Metab. 2013, 98, 2589–2594. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Boyette, L.B.; Creasey, O.A.; Guzik, L.; Lozito, T.; Tuan, R.S. Human bone marrow-derived mesenchymal stem cells display enhanced clonogenicity but impaired differentiation with hypoxic preconditioning. Stem Cells Transl. Med. 2014, 3, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Ho, I.A.; Toh, H.C.; Ng, W.H.; Teo, Y.L.; Guo, C.M.; Hui, K.M.; Lam, P.Y. Human bone marrow-derived mesenchymal stem cells suppress human glioma growth through inhibition of angiogenesis. Stem Cells 2013, 31, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, C.; Li, J.; Zhu, Y.; Zhang, X. High extracellular magnesium inhibits mineralized matrix deposition and modulates intracellular calcium signaling in human bone marrow-derived mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2014, 450, 1390–1395. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Liu, Y.; Lipsky, S.; Cho, M. Physical manipulation of calcium oscillations facilitates osteodifferentiation of human mesenchymal stem cells. FASEB J. 2007, 21, 1472–1480. [Google Scholar] [CrossRef] [PubMed]
- Zeisberg, E.M.; Tarnavski, O.; Zeisberg, M.; Dorfman, A.L.; McMullen, J.R.; Gustafsson, E.; Chandraker, A.; Yuan, X.; Pu, W.T.; Roberts, A.B.; et al. Endothelial-to-mesenchymal transition contributes to cardiac fibrosis. Nat. Med. 2007, 13, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Yi, P.; Gao, S.; Gu, Z.; Huang, T.; Wang, Z. P44/WDR77 restricts the sensitivity of proliferating cells to TGF β signaling. Biochem. Biophys. Res. Commun. 2014, 450, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Cai, Y.; Soofi, A.; Dressler, G.R. Activation of Wnt11 by transforming growth factor-β drives mesenchymal gene expression through non-canonical Wnt protein signaling in renal epithelial cells. J. Biol. Chem. 2012, 287, 21290–21302. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Wang, S.; Tang, J.; He, D.; Cui, L.; Liu, Z.; Guo, B.; Huang, L.; Lu, Y.; Hu, H. Does crystal deposition in genetic hypercalciuric rat kidney tissue share similarities with bone formation? Urology 2014, 83, 507–509. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Wang, S.; Jia, Z.; Cui, L.; Lu, Y.; Hu, H.; Qin, B. Calcium ions promote primary renal epithelial cell differentiation into cells with bone-associated phenotypes via transforming growth factor-β1-induced epithelial-mesenchymal transition in idiopathic hypercalciuria patients. Mol. Med. Rep. 2015, 11, 2199–2206. [Google Scholar] [CrossRef] [PubMed]
- Moe, O.W.; Bonny, O. Genetic hypercalciuria. J. Am. Soc. Nephrol. 2005, 16, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Strutz, F.M. EMT and proteinuria as progression factors. Kidney Int. 2009, 75, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Epithelial to mesenchymal transition in renal fibrogenesis: Pathologic significance, molecular mechanism, and therapeutic intervention. J. Am. Soc. Nephrol. 2004, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Iseki, K. Chronic kidney disease in Japan. Int. Med. 2008, 47, 681–689. [Google Scholar] [CrossRef]
- Saucier, N.A.; Sinha, M.K.; Liang, K.V.; Krambeck, A.E.; Weaver, A.L.; Bergstralh, E.J.; Li, X.; Rule, A.D.; Lieske, J.C. Risk factors for CKD in persons with kidney stones: A case-control study in Olmsted County, Minnesota. Am. J. Kidney Dis. 2010, 55, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Zacharias, A.L.; Gage, P.J. Canonical Wnt/β-catenin signaling is required for maintenance but not activation of Pitx2 expression in neural crest during eye development. Dev. Dyn. 2010, 239, 3215–3225. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Dai, C.; Li, Y.; Zeng, G.; Monga, S.P.; Liu, Y. Wnt/β-catenin signaling promotes renal interstitial fibrosis. J. Am. Soc. Nephrol. 2009, 20, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, M.A.; Joseph, J.D.; Wade, H.E.; Eaton, M.L.; Kunder, R.S.; Kazmin, D.; Chang, C.Y.; McDonnell, D.P. WNT11 expression is induced by estrogen-related receptor α and β-catenin and acts in an autocrine manner to increase cancer cell migration. Cancer Res. 2010, 70, 9298–9308. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.W.; Tadjuidje, E.; White, J.; Wells, J.; Mayhew, C.; Wylie, C.; Heasman, J. Wnt11/5a complex formation caused by tyrosine sulfation increases canonical signaling activity. Curr. Biol. 2009, 19, 1573–1580. [Google Scholar] [CrossRef] [PubMed]
- Kanlaya, R.; Sintiprungrat, K.; Thongboonkerd, V. Secreted products of macrophages exposed to calcium oxalate crystals induce epithelial mesenchymal transition of renal tubular cells via RhoA-dependent TGF-β1 pathway. Cell. Biochem. Biophys. 2013, 67, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Vuruskan, H.; Caliskan, Z.; Kordan, Y.; Ozakin, C.; Yavascaoglu, I.; Oktay, B. Elevated plasma concentrations of transforming growth factor-β 1 in patients with unilateral ureteral obstruction. Urol. Res. 2005, 33, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Westfall, T.A.; Brimeyer, R.; Twedt, J.; Gladon, J.; Olberding, A.; Furutani-Seiki, M.; Slusarski, D.C. Wnt-5/pipetail functions in vertebrate axis formation as a negative regulator of Wnt/β-catenin activity. J. Cell Biol. 2003, 162, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.J.; Nauli, S.M. Mechanosensory calcium signaling. Adv. Exp. Med. Biol. 2012, 740, 1001–1015. [Google Scholar] [PubMed]
- Monteil, A.; Chemin, J.; Bourinet, E.; Mennessier, G.; Lory, P.; Nargeot, J. Molecular and functional properties of the human α1G subunit that forms T-type calcium channels. J. Biol. Chem. 2000, 275, 6090–6100. [Google Scholar] [CrossRef] [PubMed]
- Andreasen, D.; Jensen, B.L.; Hansen, P.B.; Kwon, T.H.; Nielsen, S.; Skott, O. The α1G-subunit of a voltage-dependent Ca2+ channel is localized in rat distal nephron and collecting duct. Am. J. Physiol. Ren. Physiol. 2000, 279, F997–F1005. [Google Scholar]
- Zhao, P.L.; Wang, X.T.; Zhang, X.M.; Cebotaru, V.; Cebotaru, L.; Guo, G.; Morales, M.; Guggino, S.E. Tubular and cellular localization of the cardiac L-type calcium channel in rat kidney. Kidney Int. 2002, 61, 1393–1406. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Halagappa, V.K.; Riazi, S.; Hu, X.; Ecelbarger, C.A. Reduced expression of insulin receptors in the kidneys of insulin-resistant rats. J. Am. Soc. Nephrol. 2007, 18, 2661–2671. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Hu, X.; Riazi, S.; Tiwari, S.; Wade, J.B.; Ecelbarger, C.A. Regulation of blood pressure, the epithelial sodium channel (ENaC), and other key renal sodium transporters by chronic insulin infusion in rats. Am. J. Physiol. Ren. Physiol. 2006, 290, F1055–F1064. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, D.; Lu, Y.; Hu, H.; Zhang, J.; Qin, B.; Wang, Y.; Xing, S.; Xi, Q.; Wang, S. The Wnt11 Signaling Pathway in Potential Cellular EMT and Osteochondral Differentiation Progression in Nephrolithiasis Formation. Int. J. Mol. Sci. 2015, 16, 16313-16329. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160716313
He D, Lu Y, Hu H, Zhang J, Qin B, Wang Y, Xing S, Xi Q, Wang S. The Wnt11 Signaling Pathway in Potential Cellular EMT and Osteochondral Differentiation Progression in Nephrolithiasis Formation. International Journal of Molecular Sciences. 2015; 16(7):16313-16329. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160716313
Chicago/Turabian StyleHe, Deng, Yuchao Lu, Henglong Hu, Jiaqiao Zhang, Baolong Qin, Yufeng Wang, Shuai Xing, Qilin Xi, and Shaogang Wang. 2015. "The Wnt11 Signaling Pathway in Potential Cellular EMT and Osteochondral Differentiation Progression in Nephrolithiasis Formation" International Journal of Molecular Sciences 16, no. 7: 16313-16329. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160716313