
The Role of bFGF in the Excessive Activation of Astrocytes Is Related to the Inhibition of TLR4/NFκB Signals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
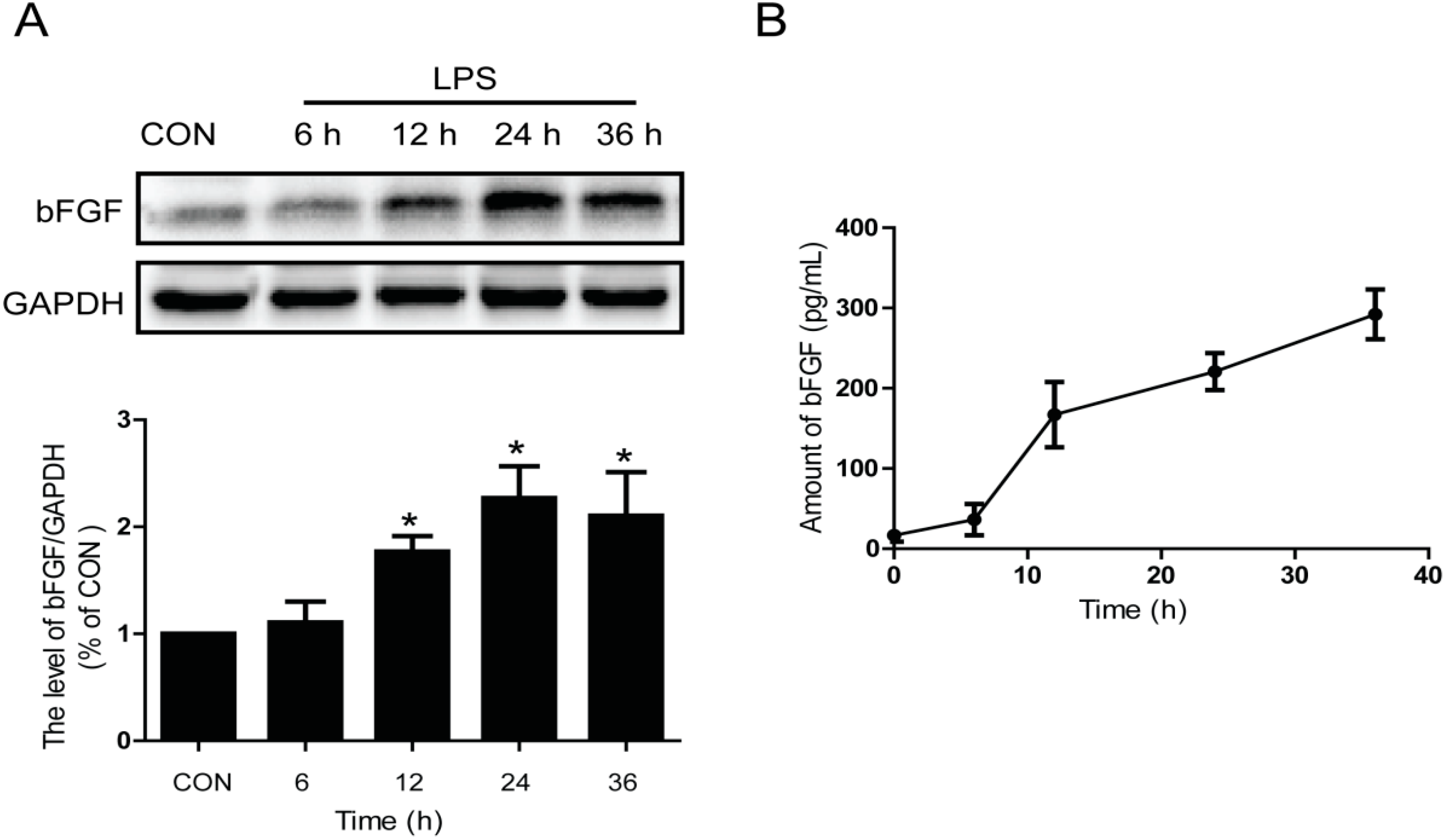
2.1. LPS Stimulates the Expression and Release of Endogenous bFGF in Primary Cultured Astrocytes

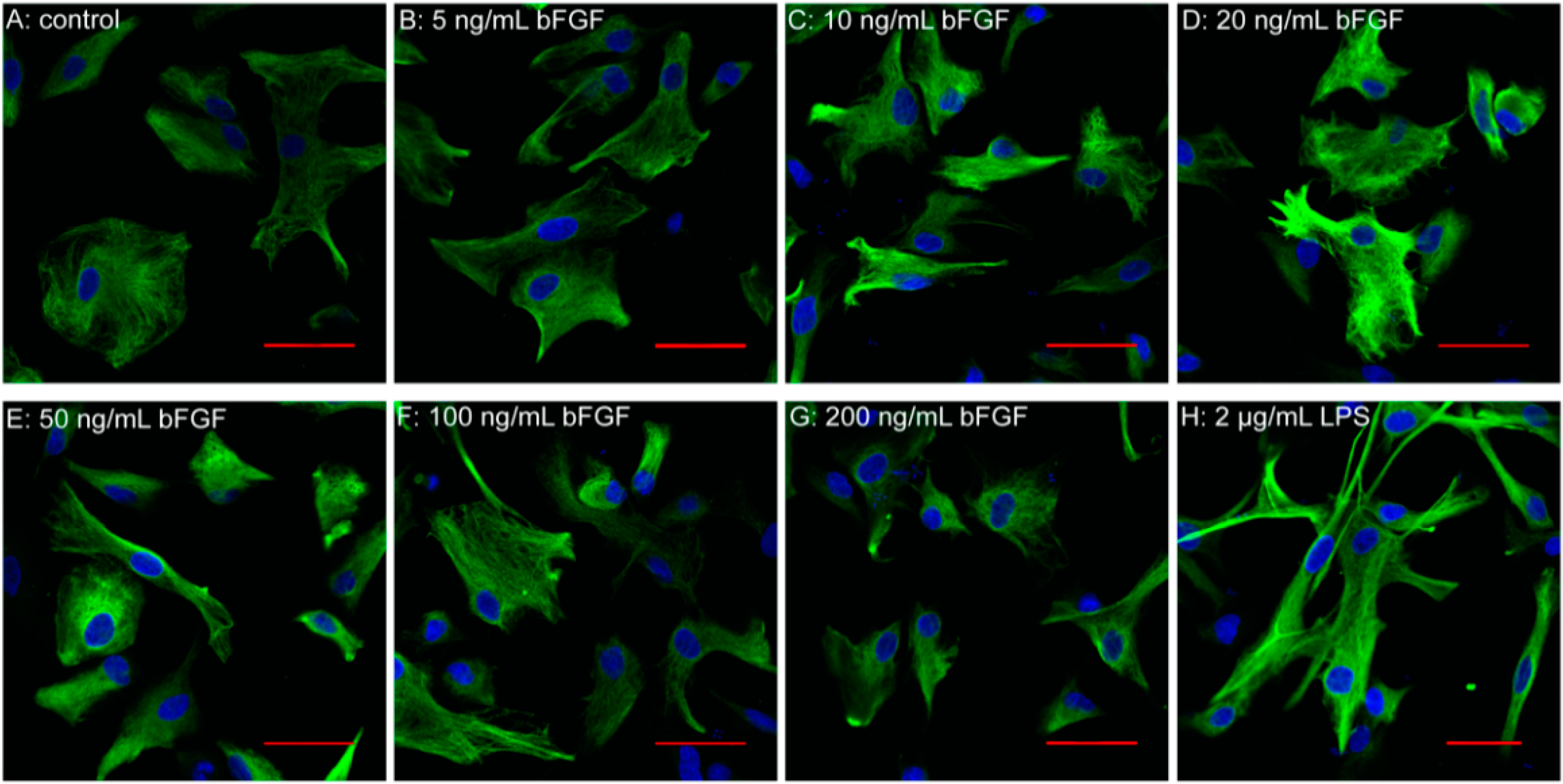
2.2. Exogenous bFGF Attenuates the Activation of Astrocytes in a High Concentration

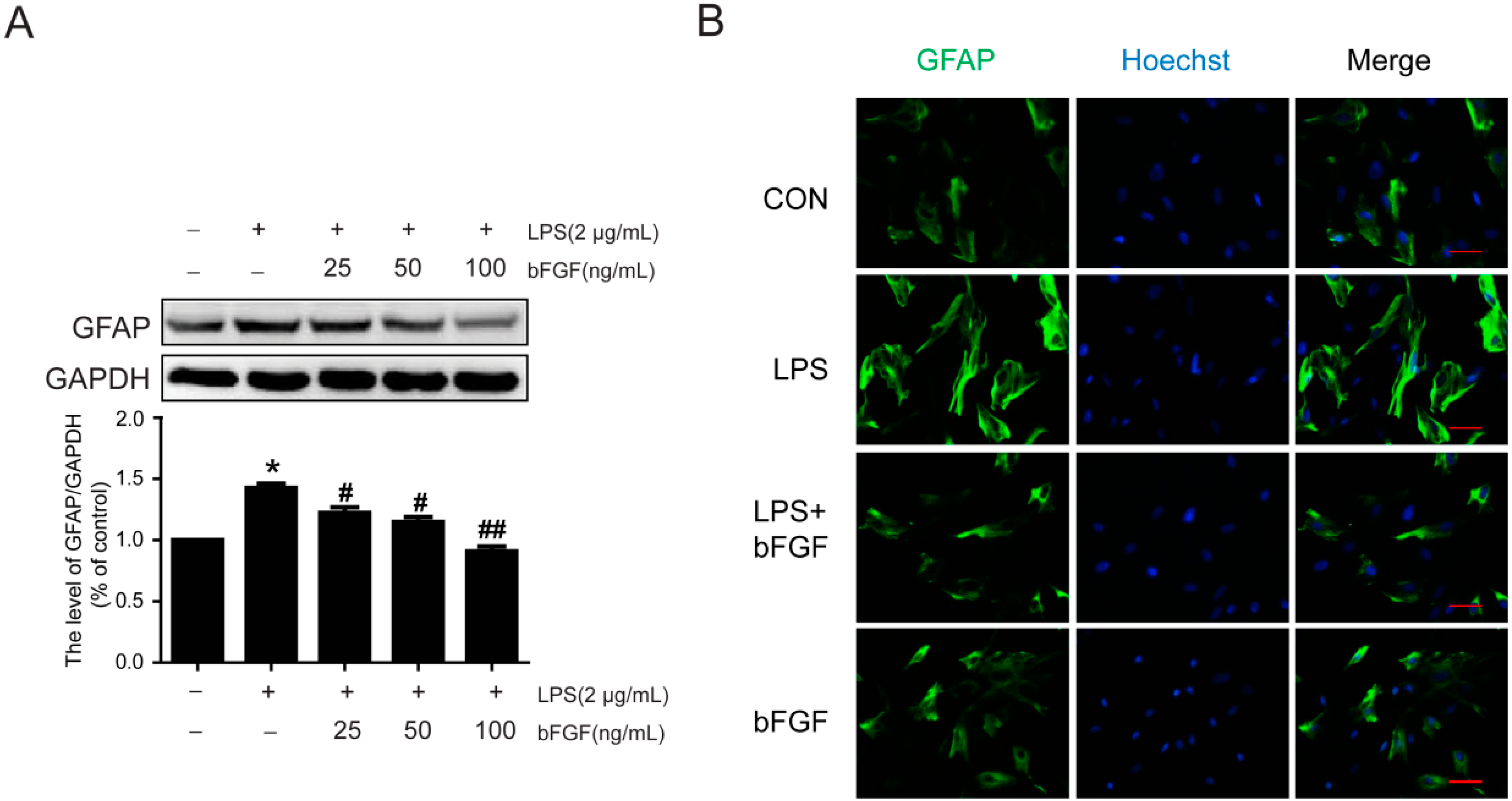
2.3. Exogenous bFGF Reduces the Expression of GFAP and Changes the Morphology in LPS Induced Astrocytes

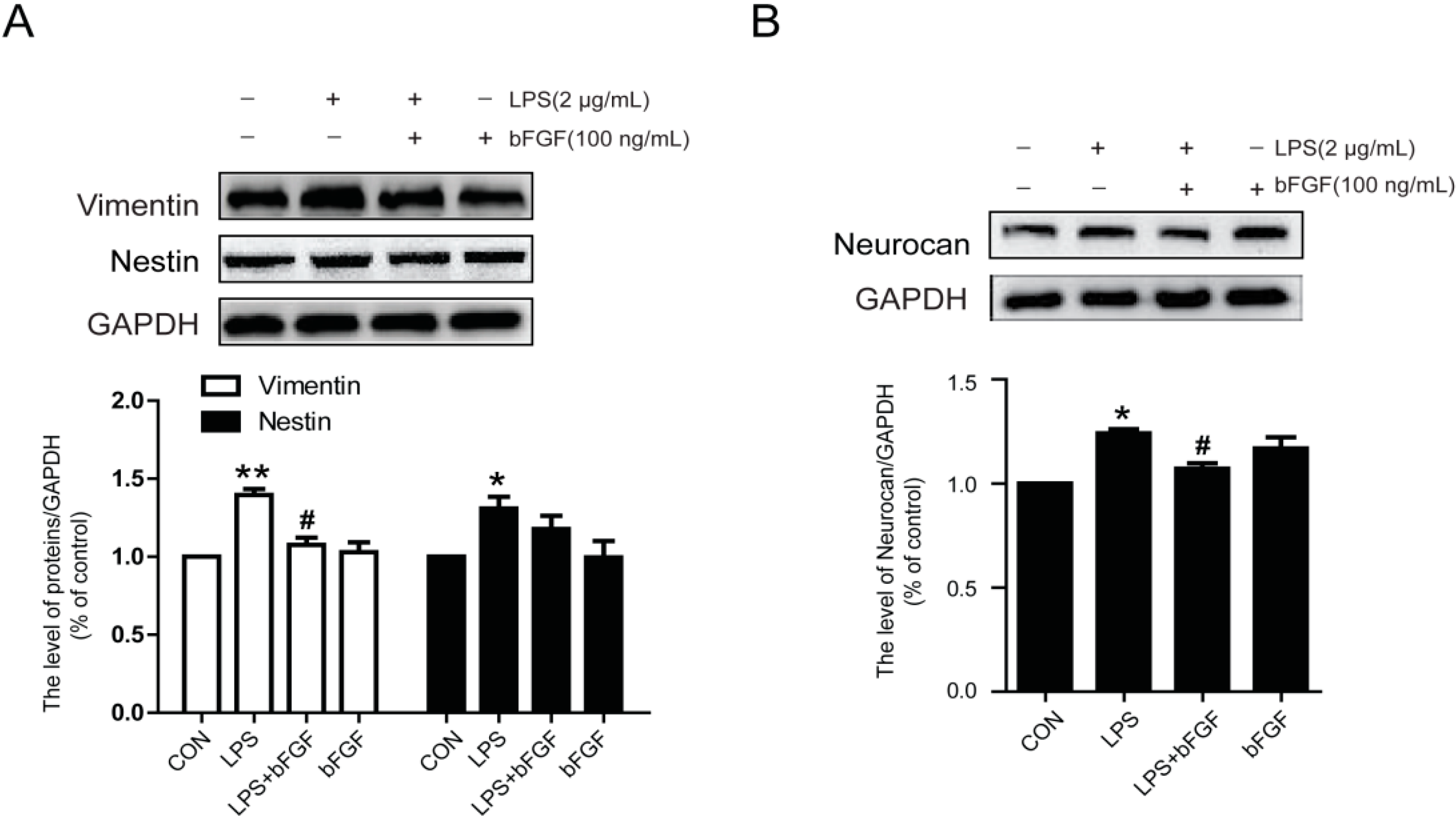
2.4. Exogenous bFGF Inhibits the Expression of Vimentin and Neurocan in LPS-Treated Astrocytes

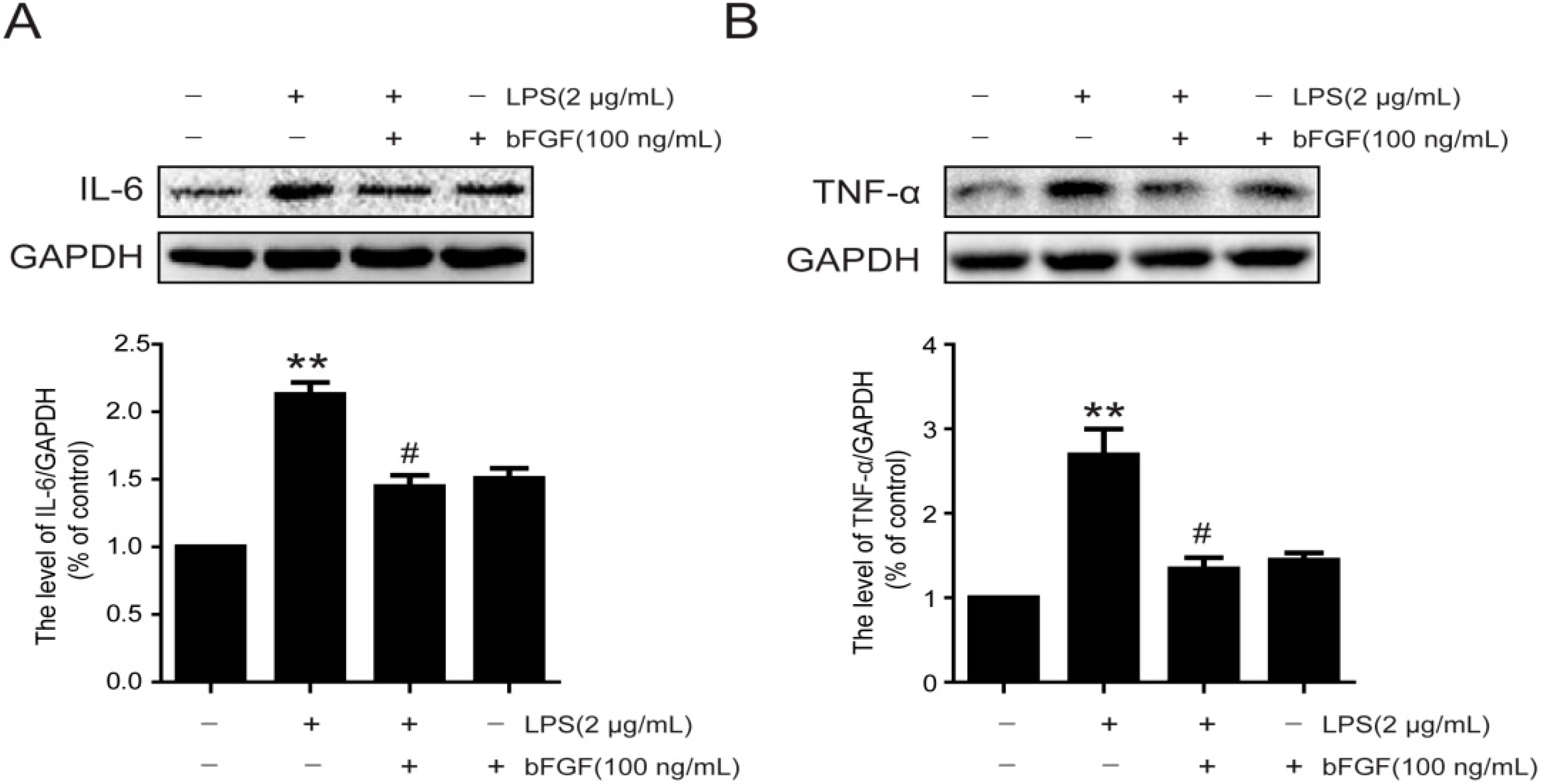
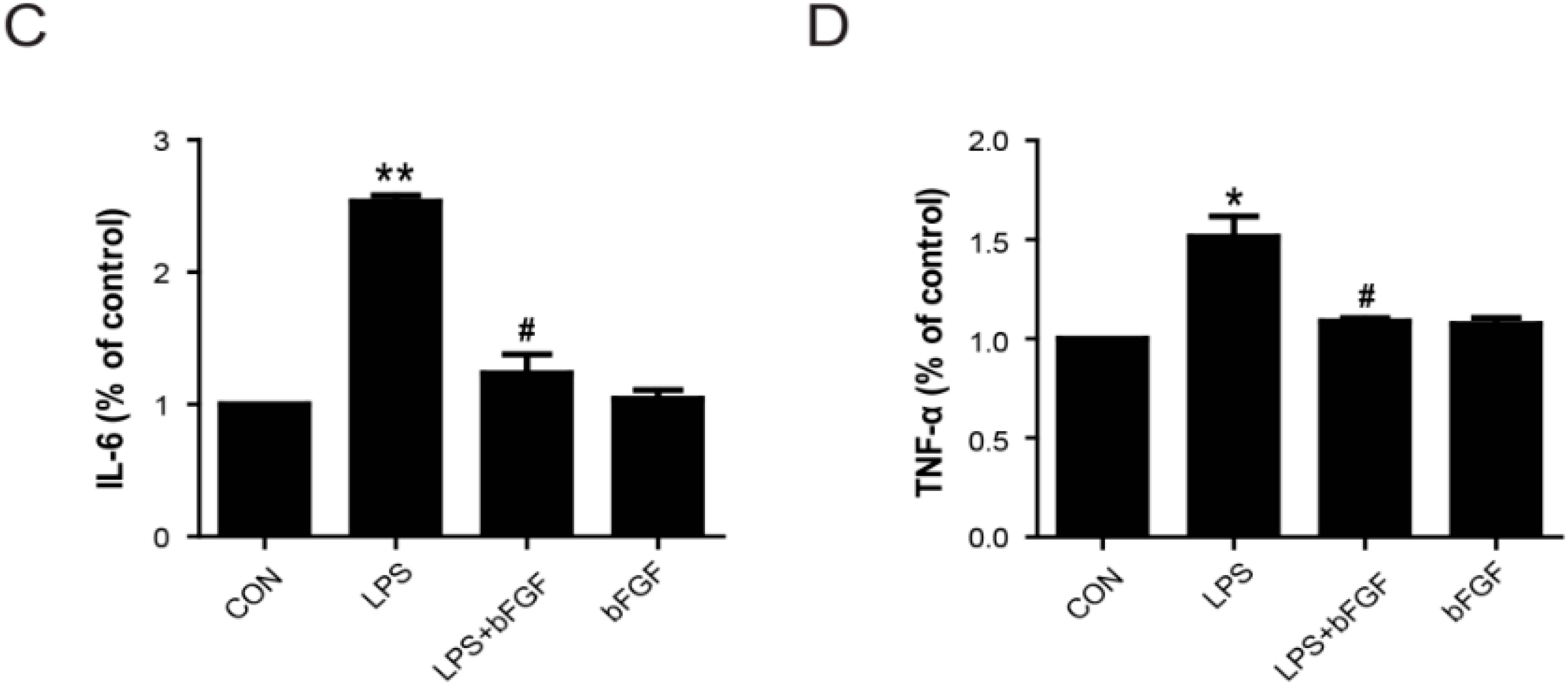
2.5. Exogenous bFGF Inhibits the Expression of Pro-Inflammatory Cytokines in LPS-stimulated Astrocytes


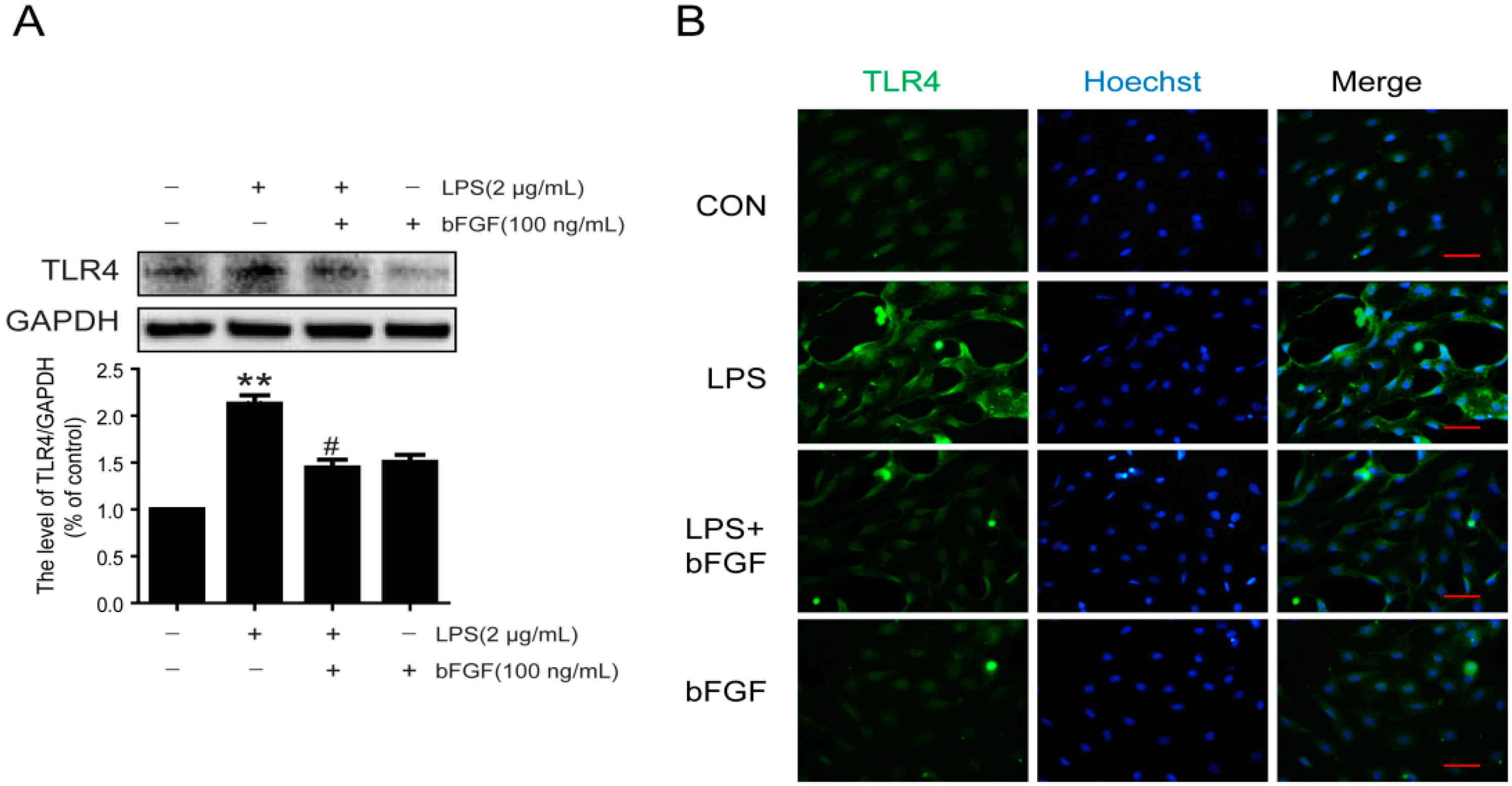
2.6. Exogenous bFGF Reduces TLR4 Expression Induced by LPS

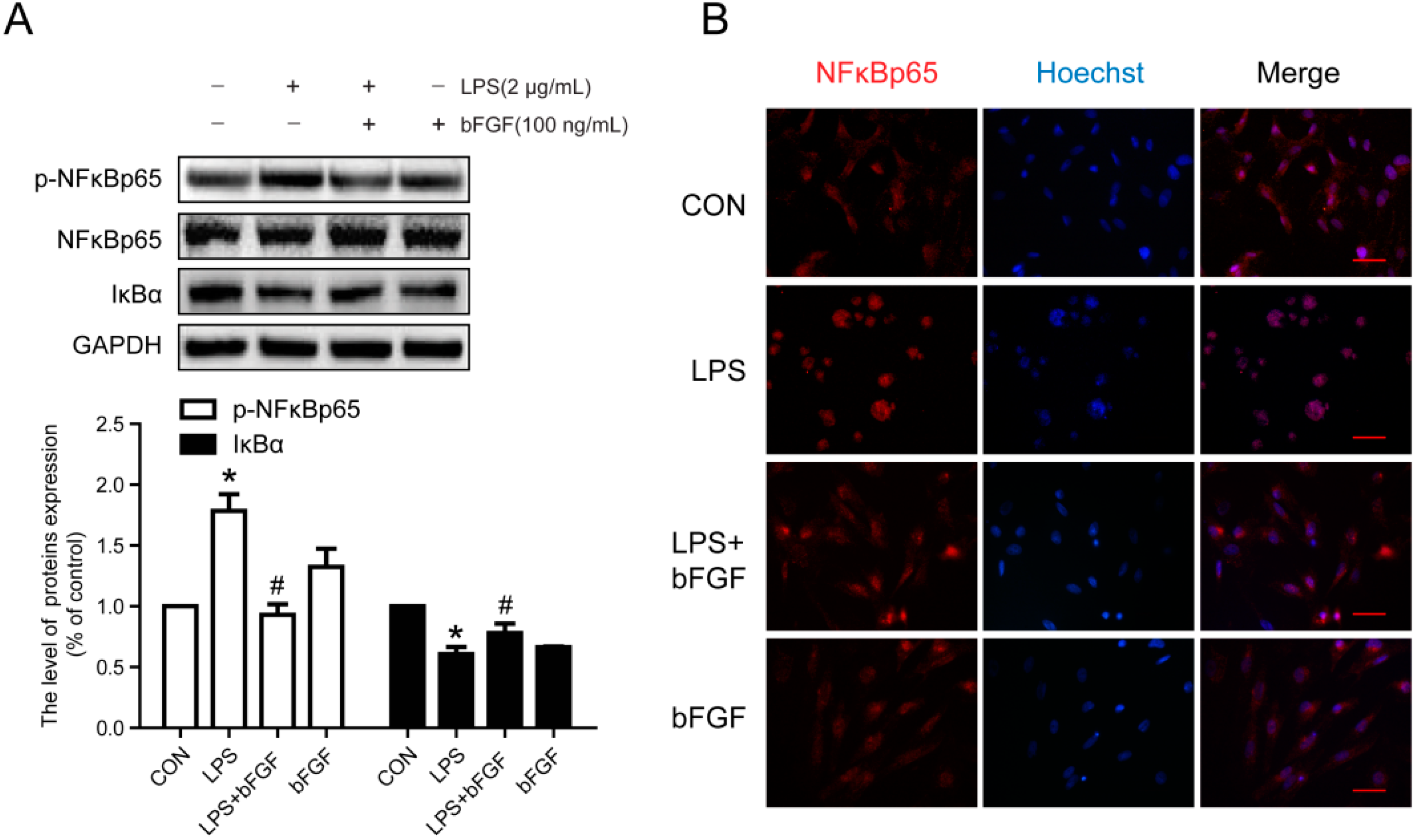
2.7. Exogenous bFGF Inhibits the Activation of NFκB in LPS-Induced Astrocytes

3. Discussion
4. Materials and Methods
4.1. Primary Astrocyte Cultures
4.2. Cell Treatment
4.3. Western Blot Analysis
4.4. Immunofluorescence Staining
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CNS | Central nervous system |
| bFGF | Basic fibroblast growth factor |
| GFAP | Glial fibrillary acidic protein |
| LPS | Lipopolysaccharide |
| NFκB | Nuclear factor κB |
| IκB | Inhibitor of κB |
| TLR4 | Toll-like receptor 4 |
References
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed]
- Gordon, G.R.; Mulligan, S.J.; MacVicar, B.A. Astrocyte control of the cerebrovasculature. Glia 2007, 55, 1214–1221. [Google Scholar] [CrossRef] [PubMed]
- Kawano, H.; Kimura-Kuroda, J.; Komuta, Y.; Yoshioka, N.; Li, H.P.; Kawamura, K.; Li, Y.; Raisman, G. Role of the lesion scar in the response to damage and repair of the central nervous system. Cell Tissue Res. 2012, 349, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Hawthorne, A.L.; Hu, H.; Kundu, B.; Steinmetz, M.P.; Wylie, C.J.; Deneris, E.S.; Silver, J. The unusual response of serotonergic neurons after cns injury: Lack of axonal dieback and enhanced sprouting within the inhibitory environment of the glial scar. J. Neurosci. 2011, 31, 5605–5616. [Google Scholar] [CrossRef] [PubMed]
- Dharmarajan, S.; Gurel, Z.; Wang, S.; Sorenson, C.M.; Sheibani, N.; Belecky-Adams, T.L. Bone morphogenetic protein 7 regulates reactive gliosis in retinal astrocytes and muller glia. Mol. Vis. 2014, 20, 1085–1108. [Google Scholar] [PubMed]
- Kelso, M.L.; Liput, D.J.; Eaves, D.W.; Nixon, K. Upregulated vimentin suggests new areas of neurodegeneration in a model of an alcohol use disorder. Neuroscience 2011, 197, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Lee, S.H.; Cha, J.H.; Seo, J.H.; Ahn, B.J.; Kim, K.W. Meteorin is upregulated in reactive astrocytes and functions as a negative feedback effector in reactive gliosis. Mol. Med. Rep. 2015, 12, 1817–1823. [Google Scholar] [CrossRef] [PubMed]
- Seo, T.B.; Chang, I.A.; Lee, J.H.; Namgung, U. Beneficial function of cell division cycle 2 activity in astrocytes on axonal regeneration after spinal cord injury. J. Neurotrauma 2013, 30, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Renault-Mihara, F.; Okada, S.; Shibata, S.; Nakamura, M.; Toyama, Y.; Okano, H. Spinal cord injury: Emerging beneficial role of reactive astrocytes’ migration. Int. J. Biochem. Cell Biol. 2008, 40, 1649–1653. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, J.E.; Imura, T.; Song, B.; Qi, J.; Ao, Y.; Nguyen, T.K.; Korsak, R.A.; Takeda, K.; Akira, S.; Sofroniew, M.V. STAT3 is a critical regulator of astrogliosis and scar formation after spinal cord injury. J. Neurosci. 2008, 28, 7231–7243. [Google Scholar] [CrossRef] [PubMed]
- Pekny, M.; Wilhelmsson, U.; Pekna, M. The dual role of astrocyte activation and reactive gliosis. Neurosci. Lett. 2014, 565, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, Q.; Yu, Z.; Zhang, L.; Tian, D.; Zhu, S.; Bu, B.; Xie, M.; Wang, W. Inhibiting cell cycle progression reduces reactive astrogliosis initiated by scratch injury in vitro and by cerebral ischemia in vivo. Glia 2007, 55, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wang, Z.; Gou, L.; Xu, H. A cortical astrocyte subpopulation inhibits axon growth in vitro and in vivo. Mol. Med. Rep. 2015, 12, 2598–2606. [Google Scholar] [CrossRef] [PubMed]
- Barcia, C.; Ros, C.M.; Annese, V.; Gomez, A.; Ros-Bernal, F.; Aguado-Yera, D.; Martinez-Pagan, M.E.; de Pablos, V.; Fernandez-Villalba, E.; Herrero, M.T. IFN-γ signaling, with the synergistic contribution of TNF-α, mediates cell specific microglial and astroglial activation in experimental models of parkinson’s disease. Cell Death Dis. 2011, 2, e142. [Google Scholar] [CrossRef] [PubMed]
- Daginakatte, G.C.; Gadzinski, A.; Emnett, R.J.; Stark, J.L.; Gonzales, E.R.; Yan, P.; Lee, J.M.; Cross, A.H.; Gutmann, D.H. Expression profiling identifies a molecular signature of reactive astrocytes stimulated by cyclic amp or proinflammatory cytokines. Exp. Neurol. 2008, 210, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Huang, Y.; Tian, C.; Zhao, Y.; Zheng, J. FOXO3a inhibits TNF-α- and IL-1β-induced astrocyte proliferation:Implication for reactive astrogliosis. Glia 2011, 59, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Sticozzi, C.; Belmonte, G.; Meini, A.; Carbotti, P.; Grasso, G.; Palmi, M. IL-1β induces GFAP expression in vitro and in vivo and protects neurons from traumatic injury-associated apoptosis in rat brain striatum via NFκB/Ca2+-calmodulin/ERK mitogen-activated protein kinase signaling pathway. Neuroscience 2013, 252, 367–383. [Google Scholar] [CrossRef] [PubMed]
- Olajide, O.A.; Bhatia, H.S.; de Oliveira, A.C.; Wright, C.W.; Fiebich, B.L. Inhibition of neuroinflammation in LPS-activated microglia by cryptolepine. Evid. Complement. Altern. Med. 2013, 459723. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R.; Preston-Hurlburt, P.; Janeway, C.A., Jr. A human homologue of the drosophila toll protein signals activation of adaptive immunity. Nature 1997, 388, 394–397. [Google Scholar] [PubMed]
- Wang, Y.; Li, C.; Cheng, K.; Zhang, R.; Narsinh, K.; Li, S.; Li, X.; Qin, X.; Zhang, R.; Li, C.; et al. Activation of liver X receptor improves viability of adipose-derived mesenchymal stem cells to attenuate myocardial ischemia injury through TLR4/NF-κB and Keap-1/Nrf-2 signaling pathways. Antioxid. Redox Signal. 2014, 21, 2543–2557. [Google Scholar] [CrossRef] [PubMed]
- Proia, P.; Schiera, G.; Mineo, M.; Ingrassia, A.M.; Santoro, G.; Savettieri, G.; di Liegro, I. Astrocytes shed extracellular vesicles that contain fibroblast growth factor-2 and vascular endothelial growth factor. Int. J. Mol. Med. 2008, 21, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, F.; Sano, Y.; Abe, M.A.; Maeda, T.; Ohtsuki, S.; Terasaki, T.; Kanda, T. Peripheral nerve pericytes modify the blood-nerve barrier function and tight junctional molecules through the secretion of various soluble factors. J. Cell. Physiol. 2011, 226, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Wu, D.; Zhang, R.; Shuang, W.; Sun, J.; Hao, H.; An, Q.; Liu, Q. Bone marrow-derived mesenchymal stem cells expressing the Shh transgene promotes functional recovery after spinal cord injury in rats. Neurosci. Lett. 2014, 573, 46–51. [Google Scholar] [CrossRef] [PubMed]
- DeLeo, J.A.; Colburn, R.W.; Rickman, A.J. Cytokine and growth factor immunohistochemical spinal profiles in two animal models of mononeuropathy. Brain Res. 1997, 759, 50–57. [Google Scholar] [CrossRef]
- Xiang, Y.; Liu, H.; Yan, T.; Zhuang, Z.; Jin, D.; Peng, Y. Functional electrical stimulation-facilitated proliferation and regeneration of neural precursor cells in the brains of rats with cerebral infarction. Neural Regen. Res. 2014, 9, 243–251. [Google Scholar] [PubMed]
- Rotschafer, J.H.; Hu, S.; Little, M.; Erickson, M.; Low, W.C.; Cheeran, M.C. Modulation of neural stem/progenitor cell proliferation during experimental herpes simplex encephalitis is mediated by differential FGF-2 expression in the adult brain. Neurobiol. Dis. 2013, 58, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pinilla, F.; Cummings, B.J.; Cotman, C.W. Induction of basic fibroblast growth factor in Alzheimer’s disease pathology. Neuroreport 1990, 1, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Goldshmit, Y.; Frisca, F.; Pinto, A.R.; Pebay, A.; Tang, J.K.; Siegel, A.L.; Kaslin, J.; Currie, P.D. FGF2 improves functional recovery-decreasing gliosis and increasing radial glia and neural progenitor cells after spinal cord injury. Brain Behav. 2014, 4, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Reilly, J.F.; Maher, P.A.; Kumari, V.G. Regulation of astrocyte GFAP expression by TGF-β1 and FGF-2. Glia 1998, 22, 202–210. [Google Scholar] [CrossRef]
- Kang, W.; Balordi, F.; Su, N.; Chen, L.; Fishell, G.; Hebert, J.M. Astrocyte activation is suppressed in both normal and injured brain by fgf signaling. Proc. Nat. Acad. Sci. USA 2014, 111, E2987–E2995. [Google Scholar] [CrossRef] [PubMed]
- Fahmy, G.H.; Moftah, M.Z. FGF-2 in astroglial cells during vertebrate spinal cord recovery. Front. Cell. Neurosci. 2010, 4, 129. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, W.; Zhou, C.; Lu, F.; Gao, T.; Liu, Y.; Cao, J.; Zhang, Y.; Zhang, Y.; Zhou, C. Ketamine inhibits lipopolysaccharide-induced astrocytes activation by suppressing TLR4/NF-κB pathway. Cell. Physiol. Biochem. 2012, 30, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, L.; Fu, H.; Wu, Y.; Wang, T. Ulinastatin suppresses lipopolysaccharide induced neuro-inflammation through the downregulation of nuclear factor-κB in SD rat hippocampal astrocyte. Biochem. Biophys. Res. Commun. 2015, 458, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Zhang, X.; Wang, Z.G.; Shi, H.X.; Wu, F.Z.; Lin, B.B.; Xu, X.L.; Wang, X.J.; Fu, X.B.; Li, Z.Y.; et al. Exogenous basic fibroblast growth factor inhibits er stress-induced apoptosis and improves recovery from spinal cord injury. CNS Neurosci. Ther. 2013, 19, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Ma, Y.F.; Gan, J.X.; Jiang, G.Y.; Xu, S.X.; Tao, X.L.; Hong, A.; Li, J.K. Basic fibroblast growth factor alleviates brain injury following global ischemia reperfusion in rabbits. J. Zhejiang Univ. Sci. B 2005, 6, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pinilla, F.; Vu, L.; Cotman, C.W. Regulation of astrocyte proliferation by FGF-2 and heparan sulfate in vivo. J. Neurosci. 1995, 15, 2021–2029. [Google Scholar] [PubMed]
- Neary, J.T.; Shi, Y.F.; Kang, Y.; Tran, M.D. Opposing effects of P2X7 and P2Y purine/pyrimidine-preferring receptors on proliferation of astrocytes induced by fibroblast growth factor-2: Implications for CNS development, injury, and repair. J. Neurosci. Res. 2008, 86, 3096–3105. [Google Scholar] [CrossRef] [PubMed]
- Lau, L.T.; Yu, A.C. Astrocytes produce and release interleukin-1, interleukin-6, tumor necrosis factor α and interferon-γ following traumatic and metabolic injury. J. Neurotrauma 2001, 18, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.R.; Kwon, M.J.; Lee, H.G.; Joe, E.H.; Lee, J.H.; Kim, S.S.; Suh-Kim, H.; Kim, B.G. Hepatocyte growth factor reduces astrocytic scar formation and promotes axonal growth beyond glial scars after spinal cord injury. Exp. Neurol. 2012, 233, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Fellner, L.; Irschick, R.; Schanda, K.; Reindl, M.; Klimaschewski, L.; Poewe, W.; Wenning, G.K.; Stefanova, N. Toll-like receptor 4 is required for α-synuclein dependent activation of microglia and astroglia. Glia 2013, 61, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.W.; Li, J.J.; Wang, L.; Zhang, J.P.; Wu, J.J.; Mao, X.Q.; Shi, G.F.; Wang, Q.; Wang, F.; Zou, J. Epidermal growth factor receptor inhibitor ameliorates excessive astrogliosis and improves the regeneration microenvironment and functional recovery in adult rats following spinal cord injury. J. Neuroinflamm. 2014, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Liu, Y.; Tu, J.; Wan, J.; Zhang, J.; Wu, B.; Chen, S.; Zhou, J.; Mu, Y.; Wang, L. Activated astrocytes enhance the dopaminergic differentiation of stem cells and promote brain repair through bFGF. Nat. Commun. 2014, 5, 5627. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, J.R.; Herrmann, J.E.; Woo, M.J.; Tansey, K.E.; Doan, N.B.; Sofroniew, M.V. Reactive astrocytes protect tissue and preserve function after spinal cord injury. J. Neurosci. 2004, 24, 2143–2155. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ye, H.; Satkunendrarajah, K.; Yao, G.S.; Bayon, Y.; Fehlings, M.G. A self-assembling peptide reduces glial scarring, attenuates post-traumatic inflammation and promotes neurological recovery following spinal cord injury. Acta Biomater. 2013, 9, 8075–8088. [Google Scholar] [CrossRef] [PubMed]
- Na, J.I.; Na, J.Y.; Choi, W.Y.; Lee, M.C.; Park, M.S.; Choi, K.H.; Lee, J.K.; Kim, K.T.; Park, J.T.; Kim, H.S. The HIF-1 inhibitor YC-1 decreases reactive astrocyte formation in a rodent ischemia model. Am. J. Transl. Res. 2015, 7, 751–760. [Google Scholar] [PubMed]
- Tang, X.; Davies, J.E.; Davies, S.J. Changes in distribution, cell associations, and protein expression levels of NG2, neurocan, phosphacan, brevican, versican V2, and tenascin-C during acute to chronic maturation of spinal cord scar tissue. J. Neurosci. Res. 2003, 71, 427–444. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.W.; Basu, A.; Druckman, C.; Cicchese, M.; Krady, J.K.; Levison, S.W. Astrogliosis is delayed in type 1 interleukin-1 receptor-null mice following a penetrating brain injury. J. Neuroinflamm. 2006, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Aranguez, I.; Torres, C.; Rubio, N. The receptor for tumor necrosis factor on murine astrocytes: Characterization, intracellular degradation, and regulation by cytokines and theiler’s murine encephalomyelitis virus. Glia 1995, 13, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Qiu, X.; Zhang, Y.; Fu, K.; Zhao, X.; Wu, J.; Hu, Y.; Zhu, W.; Guo, H. Basic fibroblast growth factor ameliorates endothelial dysfunction in radiation-induced bladder injury. Biomed Res. Int. 2015, 2015, 967680. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.C.; Park, J.H.; Ahn, J.H.; Kim, I.H.; Lee, J.C.; Yoo, K.Y.; Choi, J.H.; Hwang, I.K.; Cho, J.H.; Kwon, Y.G.; et al. Effects of high-fat diet on neuronal damage, gliosis, inflammatory process and oxidative stress in the hippocampus induced by transient cerebral ischemia. Neurochem. Res. 2014, 39, 2465–2478. [Google Scholar] [CrossRef] [PubMed]
- Alberdi, E.; Wyssenbach, A.; Alberdi, M.; Sanchez-Gomez, M.V.; Cavaliere, F.; Rodriguez, J.J.; Verkhratsky, A.; Matute, C. Ca2+-dependent endoplasmic reticulum stress correlates with astrogliosis in oligomeric amyloid β-treated astrocytes and in a model of Alzheimer’s disease. Aging Cell 2013, 12, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Kocer, G.; Naziroglu, M.; Celik, O.; Onal, L.; Ozcelik, D.; Kocer, M.; Sonmez, T.T. Basic fibroblast growth factor attenuates bisphosphonate-induced oxidative injury but decreases zinc and copper levels in oral epithelium of rat. Biol. Trace Element Res. 2013, 153, 251–256. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, L.; Yang, Y.; Zhang, X.; Cai, P.; Li, R.; Chen, D.; Wei, X.; Zhang, X.; Xu, H.; Xiao, J.; et al. The Role of bFGF in the Excessive Activation of Astrocytes Is Related to the Inhibition of TLR4/NFκB Signals. Int. J. Mol. Sci. 2016, 17, 37. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010037
Ye L, Yang Y, Zhang X, Cai P, Li R, Chen D, Wei X, Zhang X, Xu H, Xiao J, et al. The Role of bFGF in the Excessive Activation of Astrocytes Is Related to the Inhibition of TLR4/NFκB Signals. International Journal of Molecular Sciences. 2016; 17(1):37. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010037
Chicago/Turabian StyleYe, Libing, Ying Yang, Xie Zhang, Pingtao Cai, Rui Li, Daqing Chen, Xiaojie Wei, Xuesong Zhang, Huazi Xu, Jian Xiao, and et al. 2016. "The Role of bFGF in the Excessive Activation of Astrocytes Is Related to the Inhibition of TLR4/NFκB Signals" International Journal of Molecular Sciences 17, no. 1: 37. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010037