
Role of Endogenous and Exogenous Tocopherols in the Lipid Stability of Marine Oil Systems: A Review

 and
and
Abstract
:
1. Introduction
2. Lipid Oxidation in Marine Oil Systems
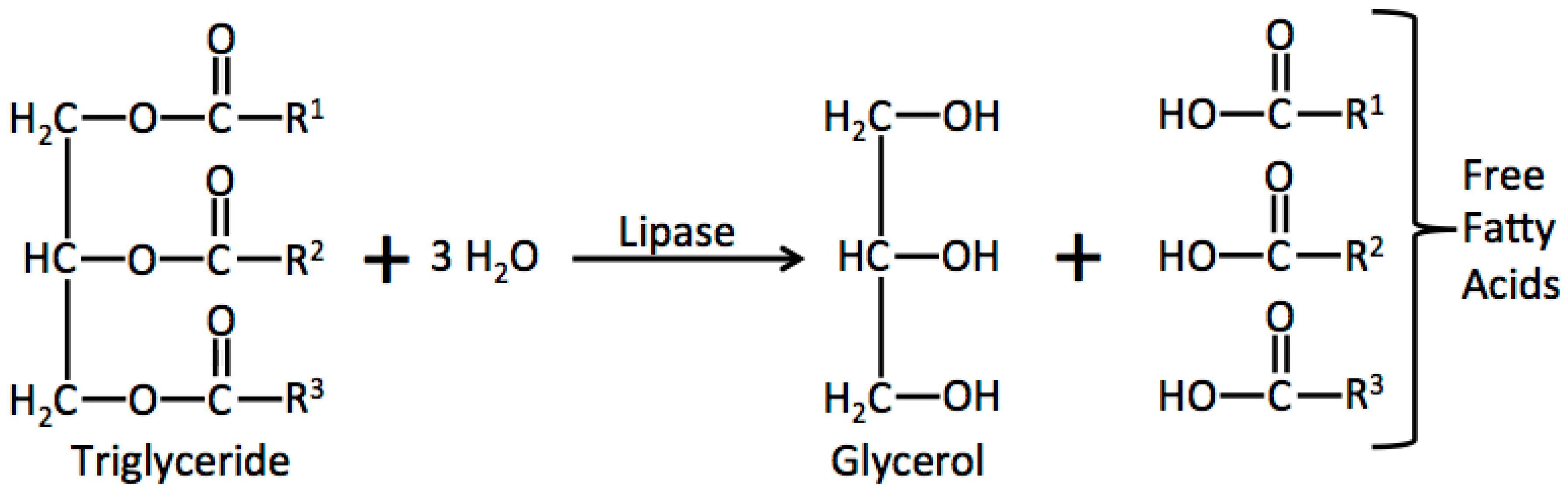
2.1. Lipid Hydrolysis
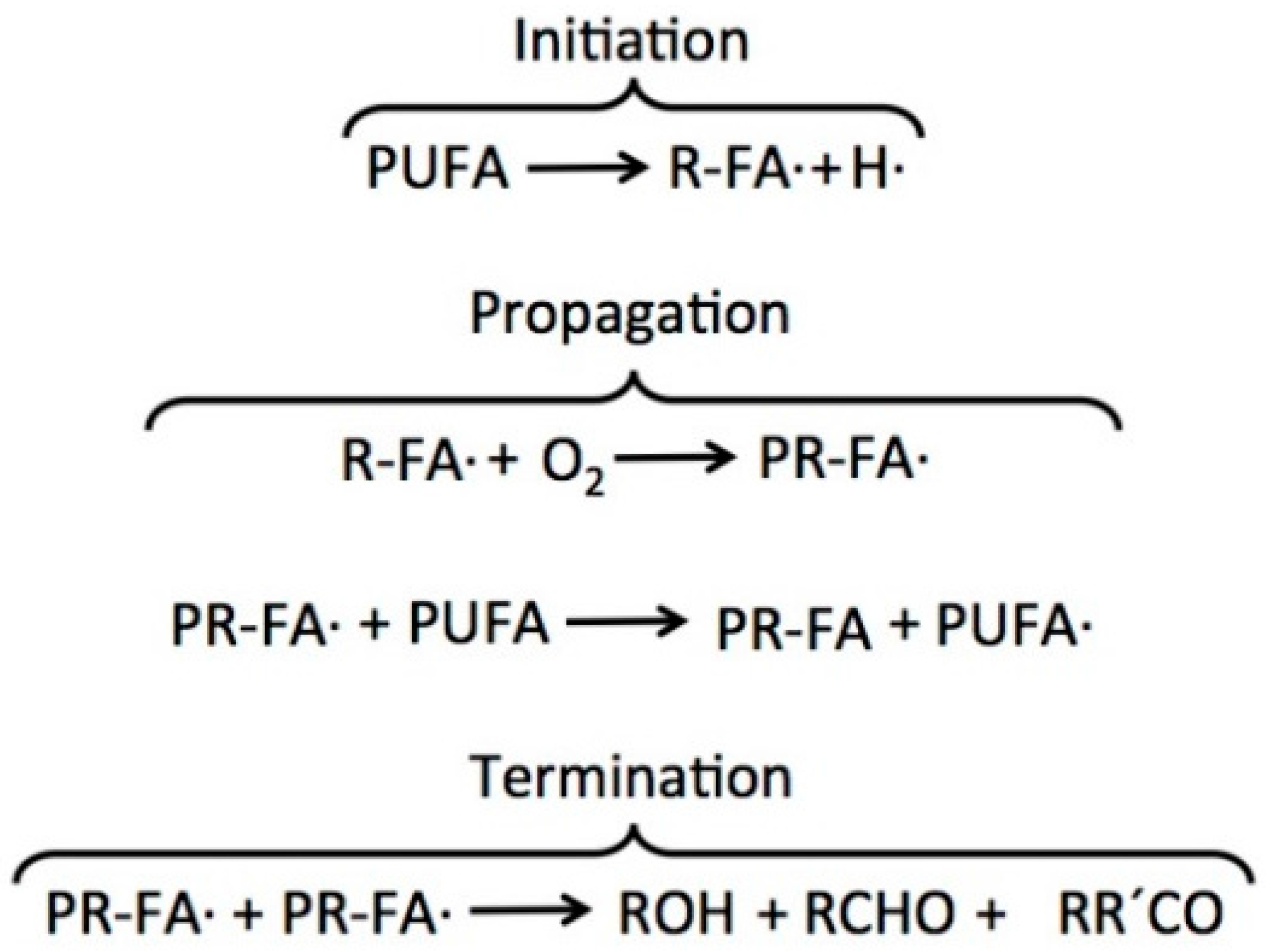
2.2. PUFAs Oxidation
2.3. Lipid Oxidation Consequences
3. Role of Antioxidants
4. Effect of Antioxidants on Fish Muscle during Storage
5. Tocopherols as Antioxidants
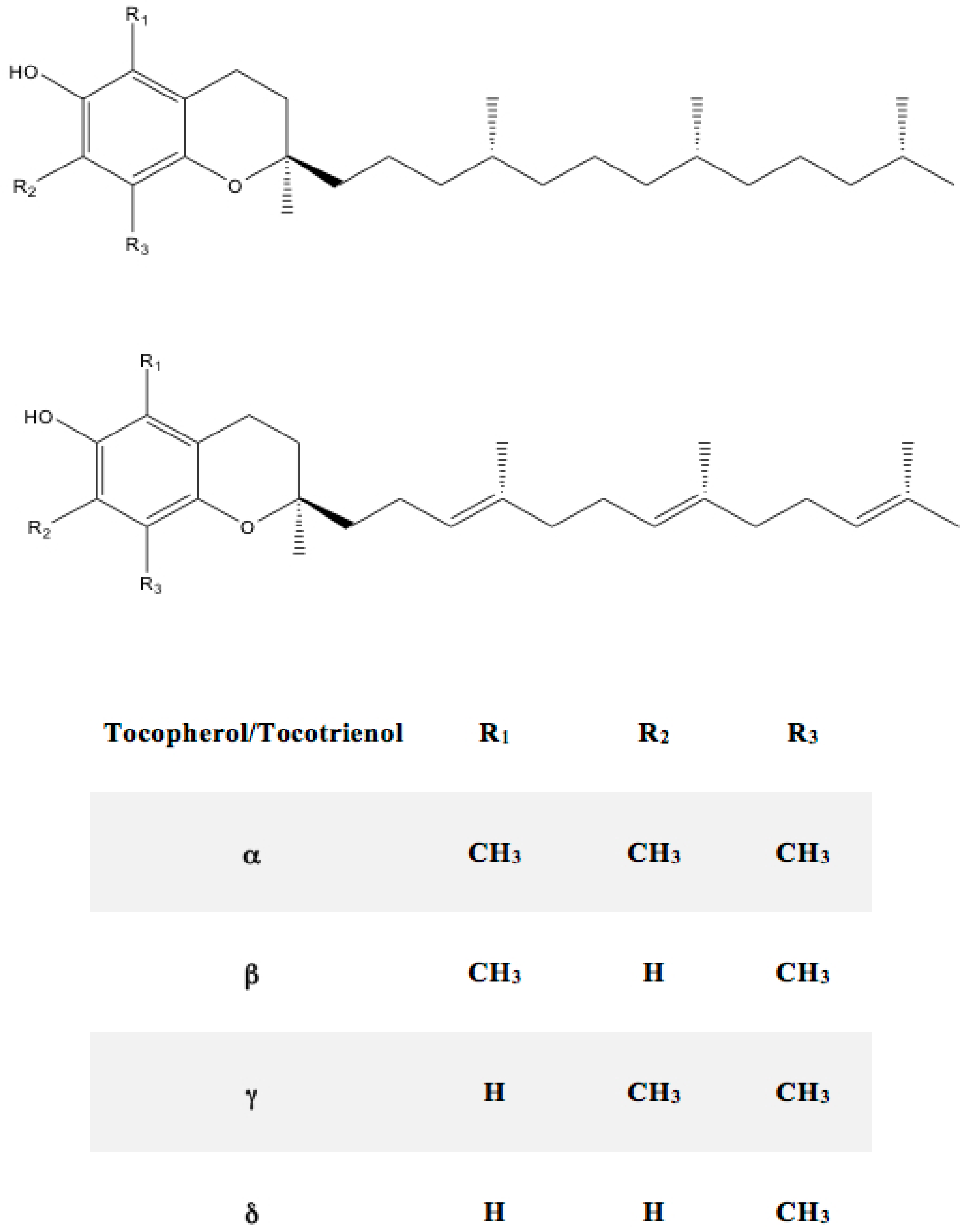
5.1. Chemistry of Tocopherols
5.2. Tocopherols’ Antioxidant Activity
5.3. Effects of Tocopherols in Living Organisms
5.4. Vitamin E Content in Fish
5.5. Tocopherols Bioavailability in Fish
5.6. The Role of Tocopherols in Cells
5.7. Endogenous Applications of Tocopherols
5.8. Exogenous Application of Tocopherols
5.9. Antioxidant Synergistic Effects of Tocopherols
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Soto-Valdez, H.; Ezquerra-Brauer, J.M.; Marquez-Rios, E.; Torres-Arreola, W. Effect of previous chilling storage on quality loss in frozen (−20 °C) sierra (Scomberomorus sierra) muscle packed with a low-density polyethylene film containing butylated hydroxytoluene. Food Sci. Technol.-Braz. 2015, 35, 202–206. [Google Scholar]
- Aubourg, S.R.; Rodriguez, A.; Gallardo, J.M. Rancidity development during frozen storage of mackerel (Scomber scombrus): Effect of catching season and commercial presentation. Eur. J. Lipid Sci. Technol. 2005, 107, 316–323. [Google Scholar] [CrossRef]
- Torres-Arreola, W.; Soto-Valdez, H.; Peralta, E.; Cárdenas-López, J.L.; Ezquerra-Brauer, J.M. Effect of a low-density polyethylene film containing butylated hydroxytoluene on lipid oxidation and protein quality of Sierra fish (Scomberomorus sierra) muscle during frozen storage. J. Agric. Food Chem. 2007, 55, 6140–6146. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Kerry, J.P.; Sheehan, D.; Buckley, D.J.; Morrissey, P.A. Antioxidative effect of added tea catechins on susceptibility of cooked red meat, poultry and fish patties to lipid oxidation. Food Res. Int. 2001, 34, 651–657. [Google Scholar] [CrossRef]
- Aubourg, S.P.; Torres-Arreola, W.; Trigo, M.; Ezquerra-Brauer, J.M. Partial characterization of jumbo squid skin pigment extract and its antioxidant potential in a marine oil system. Eur. J. Lipid Sci. Technol. 2016, 118, 1293–1304. [Google Scholar] [CrossRef]
- Huang, C.H.; Weng, Y.M. Inhibition of lipid oxidation in fish muscle by antioxidant incorporated polyethylene film. J. Food Process. Preserv. 1998, 22, 199–209. [Google Scholar] [CrossRef]
- Hraš, A.R.; Hadolin, M.; Knez, Ž.; Bauman, D. Comparison of antioxidative and synergistic effects of rosemary extract with α-tocopherol, ascorbyl palmitate and citric acid in sunflower oil. Food Chem. 2000, 71, 229–233. [Google Scholar] [CrossRef]
- Kerry, J.; Buckley, D.; Morrissey, P.; O’Sullivan, K.; Lynch, P. Endogenous and exogenous α-tocopherol supplementation: Effects on lipid stability (TBARS) and warmed-over flavour (WOF) in porcine M. longissimus dorsi roasts held in aerobic and vacuum packs. Food Res. Int. 1998, 31, 211–216. [Google Scholar] [CrossRef]
- Elisia, I.; Kitts, D.D. Modulation of NF-κB and Nrf2 control of inflammatory responses in FHs 74 Int cell line is tocopherol isoform-specific. Am. J. Physiol.-Gastrointest. Liver Physiol. 2013, 305, G940–G949. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.C. Compositional Parameters and Their Relationship to Oxidative Stability of Channel Catfish. J. Agric. Food Chem. 1993, 41, 1213–1218. [Google Scholar] [CrossRef]
- O’Keefe, T.M.; Noble, R.L. Storage stability of channel catfish (Ictalurus punctatus) in relation to dietary level of α-tocopherol. J. Fish. Res. Board Can. 1978, 35, 457–460. [Google Scholar] [CrossRef]
- Erickson, M.C. Ability of Chemical Measurements to Differentiate Oxidative Stabilities of Frozen Minced Muscle-Tissue from Farm-Raised Striped Bass and Hybrid Striped Bass. Food Chem. 1993, 48, 381–385. [Google Scholar] [CrossRef]
- Iglesias, J.; Pazos, M.; Andersen, M.L.; Skibsted, L.H.; Medina, I. Caffeic acid as antioxidant in fish muscle: Mechanism of synergism with endogenous ascorbic acid and α-tocopherol. J. Agric. Food Chem. 2008, 57, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Zuta, P.; Simpson, B.; Zhao, X.; Leclerc, L. The effect of α-tocopherol on the oxidation of mackerel oil. Food Chem. 2007, 100, 800–807. [Google Scholar] [CrossRef]
- Kul, E.; Ackman, R.G. Protection of α-tocopherol in nonpurified and purified fish oil. J. Am. Oil Chem. Soc. 2001, 78, 197–203. [Google Scholar] [CrossRef]
- Gamez-Meza, N.; Higuera-Ciapara, I.; de La Barca, A.C.; Vazquez-Moreno, L.; Noriega-Rodriguez, J.; Angulo-Guerrero, O. Seasonal variation in the fatty acid composition and quality of sardine oil from Sardinops sagax caeruleus of the Gulf of California. Lipids 1999, 34, 639–642. [Google Scholar] [CrossRef] [PubMed]
- Psota, T.L.; Gebauer, S.K.; Kris-Etherton, P. Dietary ω-3 fatty acid intake and cardiovascular risk. Am. J. Cardiol. 2006, 98, 3i–18i. [Google Scholar] [CrossRef] [PubMed]
- Trautwein, E. n-3 Fatty acids—Physiological and technical aspects for their use in food. Eur. J. Lipid Sci. Technol. 2001, 103, 45–55. [Google Scholar] [CrossRef]
- Horn, A.F.; Nielsen, N.S.; Jacobsen, C. Additions of caffeic acid, ascorbyl palmitate or γ-tocopherol to fish oil-enriched energy bars affect lipid oxidation differently. Food Chem. 2009, 112, 412–420. [Google Scholar] [CrossRef]
- Valencia-Perez, A.Z.; Soto-Valdez, H.; Ezquerra-Brauer, J.M.; Márquez-Ríos, E.; Torres-Arreola, W. Quality changes during frozen storage of blue shrimp (Litopenaeus stylirostris) with antioxidant, α-tocopherol, under different conditions. Food Sci. Technol. 2015, 35, 368–374. [Google Scholar] [CrossRef]
- Ruiz-Lopez, N.; Stubhaug, I.; Ipharraguerre, I.; Rimbach, G.; Menoyo, D. Positional Distribution of Fatty Acids in Triacylglycerols and Phospholipids from Fillets of Atlantic Salmon (Salmo Salar) Fed Vegetable and Fish Oil Blends. Mar. Drugs 2015, 13, 4255–4269. [Google Scholar] [CrossRef] [PubMed]
- Gámez-Meza, N.; Noriega-Rodríguez, J.A.; Leyva-Carrillo, L.; Ortega-García, J.; Bringas-Alvarado, L.; García, H.S.; Medina-Juárez, L.A. Antioxidant activity comparison of Thompson grape pomace extract, rosemary and tocopherols in soybean oil. J. Food Process. Preserv. 2009, 33, 110–120. [Google Scholar] [CrossRef]
- Murillo, E.; Rao, K.; Durant, A.A. The lipid content and fatty acid composition of four eastern central Pacific native fish species. J. Food Compos. Anal. 2014, 33, 1–5. [Google Scholar] [CrossRef]
- Kong, F.; Oliveira, A.; Tang, J.; Rasco, B.; Crapo, C. Salt effect on heat-induced physical and chemical changes of salmon fillet (O. gorbuscha). Food Chem. 2008, 106, 957–966. [Google Scholar] [CrossRef]
- Klinkesorn, U.; Sophanodora, P.; Chinachoti, P.; McClements, D.J.; Decker, E.A. Increasing the oxidative stability of liquid and dried tuna oil-in-water emulsions with electrostatic layer-by-layer deposition technology. J. Agric. Food Chem. 2005, 53, 4561–4566. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Aguilar, R.; Lugo-Sánchez, M.; Robles-Burgueño, M. Postmortem biochemical and functional characteristic of Monterey sardine muscle stored at 0 °C. J. Food Sci. 2000, 65, 40–47. [Google Scholar] [CrossRef]
- Sikorski, Z.E.; Kotakowski, E. Endogenous enzyme activity and seafood quality: Influence of chilling, freezing, and other environmental factors. In Endogenous Enzyme Activity and Seafood Quality: Influence of Chilling, Freezing, and Other Environmental Factors; Marcel-Dekker: New York, NY, USA, 2000; pp. 451–488. [Google Scholar]
- Rodríguez, A.; Losada, V.; Larraín, M.A.; Quitral, V.; Vinagre, J.; Aubourg, S.P. Development of lipid changes related to quality loss during the frozen storage of farmed coho salmon (Oncorhynchus kisutch). J. Am. Oil Chem. Soc. 2007, 84, 727–734. [Google Scholar] [CrossRef]
- Aubourg, S.P.; Sotelo, C.G.; Perez-Martin, R. Assessment of quality changes in frozen sardine (Sardina pilchardus) by fluorescence detection. J. Am. Oil Chem. Soc. 1998, 75, 575–580. [Google Scholar] [CrossRef] [Green Version]
- Medina, I.; Aubourg, S.P.; Martín, R.P. Composition of phospholipids of white muscle of six tuna species. Lipids 1995, 30, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Losada, V.; Barros-Velázquez, J.; Aubourg, S.P. Rancidity development in frozen pelagic fish: Influence of slurry ice as preliminary chilling treatment. LWT-Food Sci. Technol. 2007, 40, 991–999. [Google Scholar] [CrossRef] [Green Version]
- Miyashita, K.; Takagi, T. Study on the oxidative rate and prooxidant activity of free fatty acids. J. Am. Oil Chem. Soc. 1986, 63, 1380–1384. [Google Scholar] [CrossRef]
- Erickson, M.C. Lipid oxidation: Flavor and nutritional quality deterioration in frozen foods. In Quality in Frozen Food; Springer: New York, NY, USA, 1997; pp. 141–173. [Google Scholar]
- Frankel, E. Lipid Oxidation. Prog. Lipid Res. 1998, 1, 1–22. [Google Scholar]
- Buettner, G.R. The pecking order of free radicals and antioxidants: Lipid peroxidation, α-tocopherol, and ascorbate. Arch. Biochem. Biophys. 1993, 300, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Bjerkeng, B.; Johnsen, G. Frozen storage quality of rainbow trout (Oncorhynchus mykiss) as affected by oxygen, illumination, and fillet pigment. J. Food Sci. 1995, 60, 284–288. [Google Scholar] [CrossRef]
- Evans, H.M.; Bishop, K.S. On the existence of a hitherto unrecognized dietary factor essential for reproduction. Science 1922, 56, 650–651. [Google Scholar] [CrossRef] [PubMed]
- Kamal-Eldin, A.; Appelqvist, L.-Å. The chemistry and antioxidant properties of tocopherols and tocotrienols. Lipids 1996, 31, 671–701. [Google Scholar] [CrossRef] [PubMed]
- Rosa, C.; Blake, J.E.; Mazzaro, L.; Hoekstra, P.; Ylitalo, G.M.; O’Hara, T.M. Vitamin A and E tissue distribution with comparisons to organochlorine concentrations in the serum, blubber and liver of the bowhead whale (Balaena mysticetus). Comp. Biochem. Physiol. B-Biochem. Mol. Biol. 2007, 148, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.-S. Tocopherols and tocotrienols in plants and their products: A review on methods of extraction, chromatographic separation, and detection. Food Res. Int. 2016, 82, 59–70. [Google Scholar] [CrossRef]
- Oliva-Teles, A. Nutrition and health of aquaculture fish. J. Fish Dis. 2012, 35, 83–108. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.L. An update on vitamin E, tocopherol and tocotrienol—Perspectives. Molecules 2010, 15, 2103–2113. [Google Scholar] [CrossRef] [PubMed]
- Raiola, A.; Tenore, G.C.; Barone, A.; Frusciante, L.; Rigano, M.M. Vitamin E Content and Composition in Tomato Fruits: Beneficial Roles and Bio-Fortification. Int. J. Mol. Sci. 2015, 16, 29250–29264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, C.Y.; Kim, Y.-H.; Kim, H.S.; Ke, Q.; Kim, G.-W.; Park, S.-C.; Lee, H.-S.; Jeong, J.C.; Kwak, S.-S. Molecular characterization of tocopherol biosynthetic genes in sweetpotato that respond to stress and activate the tocopherol production in tobacco. Plant Physiol. Biochem. 2016, 106, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Packer, L.; Weber, S.U.; Rimbach, G. Molecular aspects of α-tocotrienol antioxidant action and cell signalling. J. Nutr. 2001, 131, 369S–373S. [Google Scholar] [PubMed]
- Yamauchi, R.; Yagi, Y.; Kato, K. Oxidation of α-tocopherol during the peroxidation of dilinoleoylphosphatidylcholine in liposomes. Biosci. Biotechnol. Biochem. 1996, 60, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Kontush, A.; Finckh, B.; Karten, B.; Kohlschütter, A.; Beisiegel, U. Antioxidant and prooxidant activity of α-tocopherol in human plasma and low density lipoprotein. J. Lipid Res. 1996, 37, 1436–1448. [Google Scholar] [PubMed]
- Brewer, M. Natural antioxidants: Sources, compounds, mechanisms of action, and potential applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Steiner, M. Vitamin E: more than an antioxidant. Clin. Cardiol. 1993, 16, 16–18. [Google Scholar] [CrossRef]
- Al-Serwi, R.H.; Ghoneim, F.M. The impact of vitamin E against acrylamide induced toxicity on skeletal muscles of adult male albino rat tongue: Light and electron microscopic study. J. Microsc. Ultrastruct. 2015, 3, 137–147. [Google Scholar] [CrossRef]
- Traber, M.G.; Stevens, J.F. Vitamins C and E: Beneficial effects from a mechanistic perspective. Free Radic. Biol. Med. 2011, 51, 1000–1013. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-Y.; Appel, L.J.; Croft, K.D.; Miller, E.R.; Mori, T.A.; Puddey, I.B. Effects of vitamin C and vitamin E on in vivo lipid peroxidation: Results of a randomized controlled trial. Am. J. Clin. Nutr. 2002, 76, 549–555. [Google Scholar] [PubMed]
- Obach, A.; Quentel, C.; Laurencin, F.B. Effects of α-tocopherol and dietary oxidized fish oil on the immune response of sea bass Dicentrarchus labrax. Dis. Aquat. Org. 1993, 15, 175–185. [Google Scholar] [CrossRef]
- Nascimento, T.S.R.; de Stéfani, M.V.; Malheiros, E.B.; Koberstein, T.C.R.D. High levels of dietary vitamin E improve the reproductive performance of female Oreochromis niloticus. Acta Sci. Biol. Sci. 2013, 36, 19–26. [Google Scholar]
- Amlashi, A.S.; Falahatkar, B.; Sharifi, S.D. Dietary vitamin E requirements and growth performance of young-of-the-year beluga, Huso huso (L.) (Chondrostei: Acipenseridae). Arch. Pol. Fish. 2012, 20, 299–306. [Google Scholar] [CrossRef]
- Ricciarelli, R.; Zingg, J.M.; Azzi, A. Vitamin E: Protective role of a Janus molecule. FASEB J. 2001, 15, 2314–2325. [Google Scholar] [CrossRef] [PubMed]
- Gagné, A.; Wei, S.Q.; Fraser, W.D.; Julien, P. Absorption, transport, and bioavailability of vitamin E and its role in pregnant women. J. Obstet. Gynaecol. Can. 2009, 31, 210–217. [Google Scholar] [CrossRef]
- Parazo, M.P.; Lall, S.P.; Castell, J.D.; Ackman, R.G. Distribution of α-and γ-tocopherols in Atlantic salmon (Salmo salar) tissues. Lipids 1998, 33, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Hamre, K. Metabolism, interactions, requirements and functions of vitamin E in fish. Aquac. Nutr. 2011, 17, 98–115. [Google Scholar] [CrossRef]
- Wang, X.; Quinn, P.J. The location and function of vitamin E in membranes (Review). Mol. Membr. Biol. 2000, 17, 143–156. [Google Scholar] [PubMed]
- Mashaii, N.; Mosaddegh, M.H.; Sarsangi, H.; Rajabipour, F.; Ghorghi, A.; Bitaraf, A.; Mozaffari-Khosravi, H. Proximate and Fatty Acid Composition in Muscle Tissues of Rainbow Trout, Oncorhynchus Mykiss, Cultured in Yazd Province of Iran. Walailak J. Sci. Technol. 2012, 9, 317–325. [Google Scholar]
- Jittinandana, S.; Kenney, P.B.; Slider, S.D.; Kamireddy, N.; Hankins, J. High dietary vitamin E affects storage stability of frozen-refrigerated trout fillets. J. Food Sci. 2006, 71, C91–C96. [Google Scholar] [CrossRef]
- Tocher, D.R.; Mourente, G.; van der Eecken, A.; Evjemo, J.O.; Diaz, E.; Bell, J.G.; Geurden, I.; Lavens, P.; Olsen, Y. Effects of dietary vitamin E on antioxidant defence mechanisms of juvenile turbot (Scophthalmus maximus L.), halibut (Hippoglossus hippoglossus L.) and sea bream (Sparus aurata L.). Aquac. Nutr. 2002, 8, 195–207. [Google Scholar] [CrossRef]
- Gao, J.; Koshio, S. Effect of dietary lipid oxidation with vitamin C and E supplementation on fillet quality of red sea bream, Pagrus major (Temminck & Schlegel) during storage. Aquac. Res. 2015, 46, 2382–2391. [Google Scholar]
- Hemre, G.; Juell, J.E.; Hamre, K.; Lie, Ø.; Strand, B.; Arnesen, P.; Holm, J.C. Cage feeding of Atlantic mackerel (Scomber scombrus): Effect on muscle lipid content, fatty acid composition, oxidation status and vitamin E concentration. Aquat. Living Resour. 1997, 10, 365–370. [Google Scholar] [CrossRef]
- Petillo, D.; Hultin, H.O.; Krzynowek, J.; Autio, W.R. Kinetics of antioxidant loss in mackerel light and dark muscle. J. Agric. Food Chem. 1998, 46, 4128–4137. [Google Scholar] [CrossRef]
- Chan, K.M.; Decker, E.A.; Feustman, C. Endogenous skeletal muscle antioxidants. Crit. Rev. Food Sci. Nutr. 1994, 34, 403–426. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, N.S.; Jacobsen, C. Retardation of lipid oxidation in fish oil-enriched fish pâté–combination effects. J. Food Biochem. 2013, 37, 88–97. [Google Scholar] [CrossRef]
- Belhaj, N.; Arab-Tehrany, E.; Linder, M. Oxidative kinetics of salmon oil in bulk and in nanoemulsion stabilized by marine lecithin. Process Biochem. 2010, 45, 187–195. [Google Scholar] [CrossRef]
- Drusch, S.; Serfert, Y.; Scampicchio, M.; Schmidt-Hansberg, B.; Schwarz, K. Impact of physicochemical characteristics on the oxidative stability of fish oil microencapsulated by spray-drying. J. Agric. Food Chem. 2007, 55, 11044–11051. [Google Scholar] [CrossRef] [PubMed]
- Let, M.B.; Jacobsen, C.; Meyer, A.S. Lipid oxidation in milk, yoghurt, and salad dressing enriched with neat fish oil or pre-emulsified fish oil. J. Agric. Food Chem. 2007, 55, 7802–7809. [Google Scholar] [CrossRef] [PubMed]
- Hogan, S.; O’riordan, E.; O’sullivan, M. Microencapsulation and oxidative stability of spray-dried fish oil emulsions. J. Microencapsul. 2003, 20, 675–688. [Google Scholar] [CrossRef] [PubMed]
- Coppes Petricorena, Z. Chemical Composition of Fish and Fishery Products. Chem. Comp. Fish Fishery Prod. 2015. [Google Scholar] [CrossRef]
- Pankyamma, V.; Basu, S.; Bhadran, S.S.; Chouksey, M.K.; Gudipati, V. Fish Oil-Fortified Extruded Snacks: Evaluation of Physical Properties and Oxidative Stability by Response Surface Methodology. J. Food Process. Eng. 2014, 37, 349–361. [Google Scholar] [CrossRef]
- Hamilton, R.; Kalu, C.; McNeill, G.; Padley, F.; Pierce, J. Effects of tocopherols, ascorbyl palmitate, and lecithin on autoxidation of fish oil. J. Am. Oil Chem. Soc. 1998, 75, 813–822. [Google Scholar] [CrossRef]
- Vicetti, R.; Ishitani, T.; Salas, A.; Ayala, M. Use of α-tocopherol combined with synergists and compared to other antioxidants on the oxidative stability of sardine skin lipids. J. Food Compos. Anal. 2005, 18, 131–137. [Google Scholar] [CrossRef]
- Yi, J.; Andersen, M.L.; Skibsted, L.H. Interactions between tocopherols, tocotrienols and carotenoids during autoxidation of mixed palm olein and fish oil. Food Chem. 2011, 127, 1792–1797. [Google Scholar] [CrossRef]
- Barrett, A.H.; Porter, W.L.; Marando, G.; Chinachoti, P. Effect of various antioxidants, antioxidant levels, and encapsulation on the stability of fish and flaxseed oils: Assessment by fluorometric analysis. J. Food Process. Preserv. 2011, 35, 349–358. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marine Source | PUFAs Content (g/100 g Total Fatty Acids) | Reference |
|---|---|---|
| Atlantic Salmon (Salmo salar) | ≈28 | [21] |
| Atlantic menhaden (Brevoortia tyrannus) | ≈26 | [22] |
| Atlantic Cod (Gadus morhua) | ≈22 | [5] |
| Monterey sardine (Sardinops sagax caerulea) | ≈25 | [22] |
| Pacific sierra (Scomberomorus sierra) | ≈28 | [23] |
| Pacific corvina (Cynoscion phoxocephalus) | ≈23 | [23] |
| Pink Salmon (Oncorhynchus gorbuscha) | ≈30 | [24] |
| Tuna fish (Thunnus) | ≈35 | [25] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suárez-Jiménez, G.M.; López-Saiz, C.M.; Ramírez-Guerra, H.E.; Ezquerra-Brauer, J.M.; Ruiz-Cruz, S.; Torres-Arreola, W. Role of Endogenous and Exogenous Tocopherols in the Lipid Stability of Marine Oil Systems: A Review. Int. J. Mol. Sci. 2016, 17, 1968. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17121968
Suárez-Jiménez GM, López-Saiz CM, Ramírez-Guerra HE, Ezquerra-Brauer JM, Ruiz-Cruz S, Torres-Arreola W. Role of Endogenous and Exogenous Tocopherols in the Lipid Stability of Marine Oil Systems: A Review. International Journal of Molecular Sciences. 2016; 17(12):1968. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17121968
Chicago/Turabian StyleSuárez-Jiménez, Guadalupe Miroslava, Carmen María López-Saiz, Hugo Enrique Ramírez-Guerra, Josafat Marina Ezquerra-Brauer, Saul Ruiz-Cruz, and Wilfrido Torres-Arreola. 2016. "Role of Endogenous and Exogenous Tocopherols in the Lipid Stability of Marine Oil Systems: A Review" International Journal of Molecular Sciences 17, no. 12: 1968. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17121968