
MicroRNA Expression Profiling in CCl4-Induced Liver Fibrosis of Mus musculus

Abstract
:
1. Introduction
2. Results
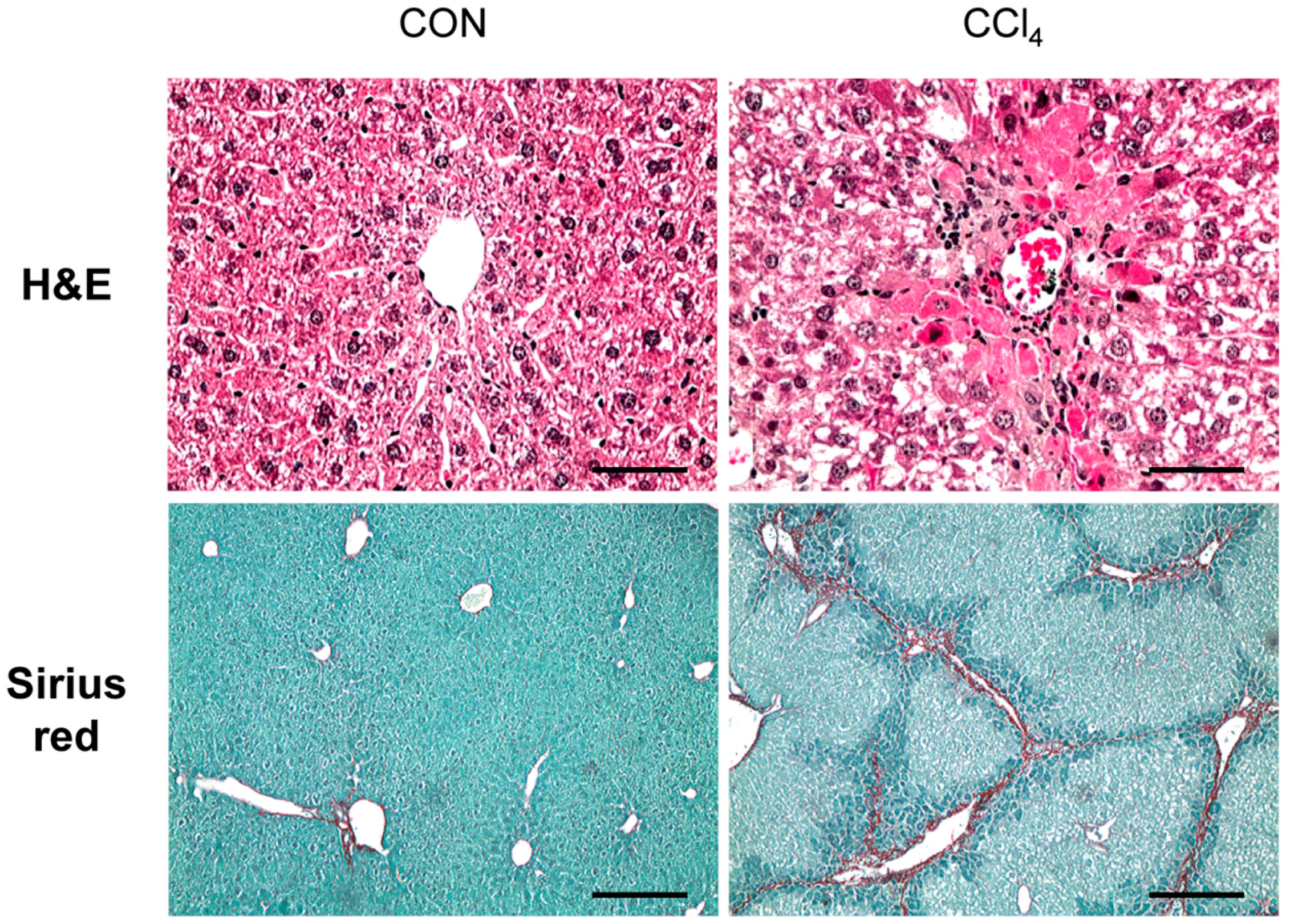
2.1. Carbon-Tetrachloride-Induced Liver Fibrosis in Mice
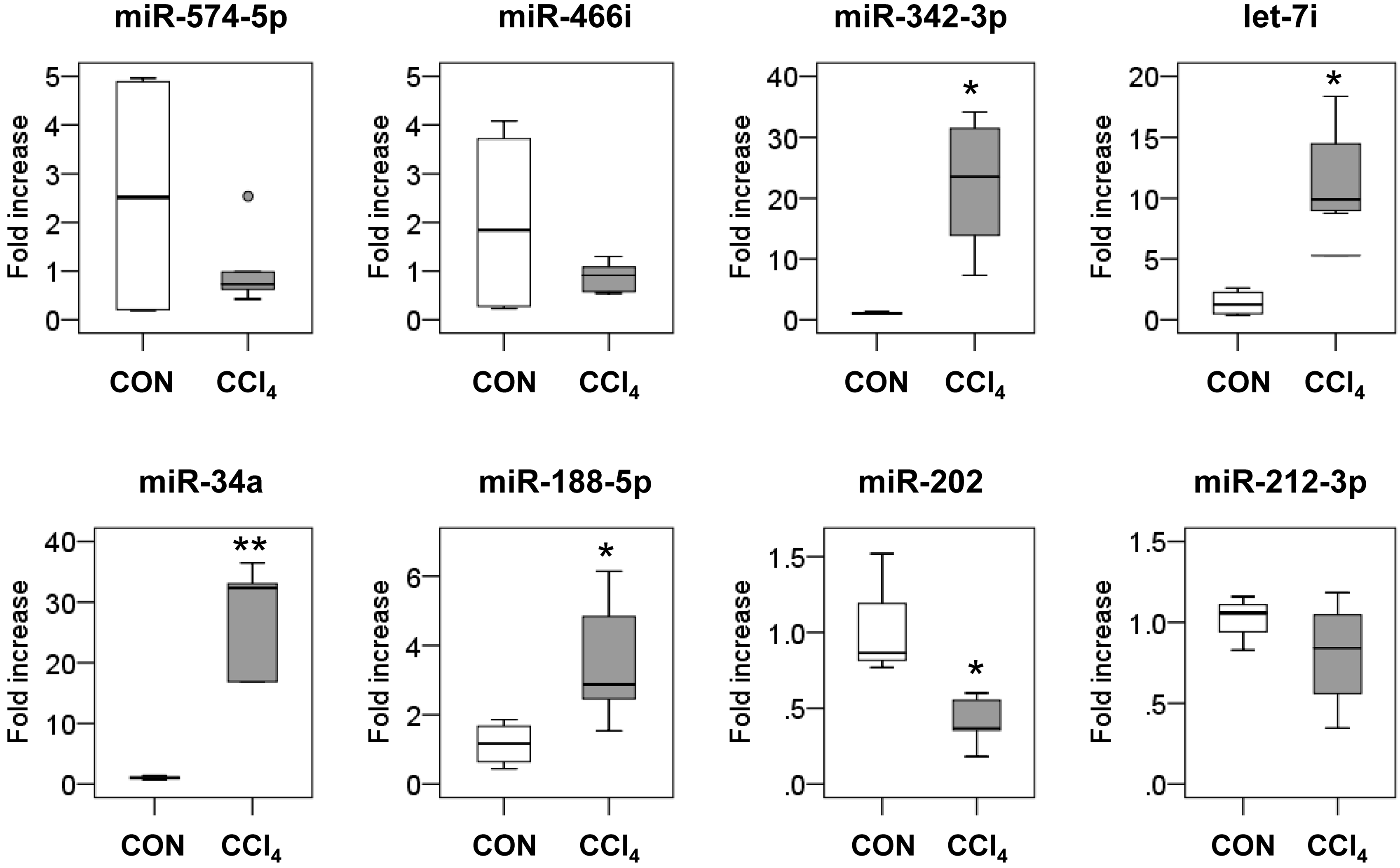
2.2. Dysregulated MicroRNAs’ (miRNAs) Expression in Fibrotic Livers
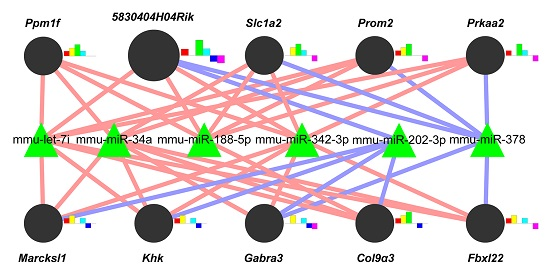
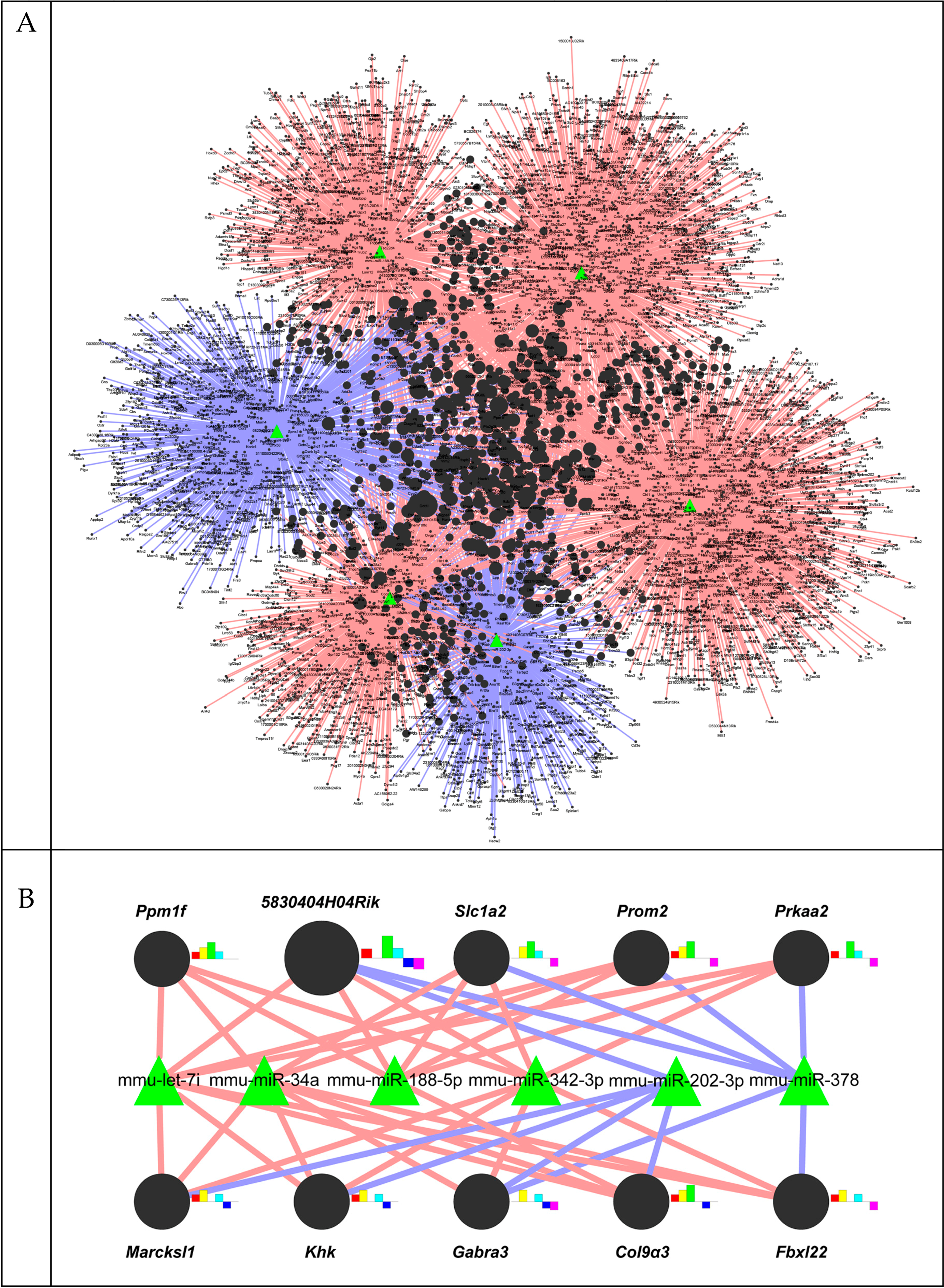
2.3. A Gene-Regulatory Network of miRNAs Altered in Liver Fibrosis
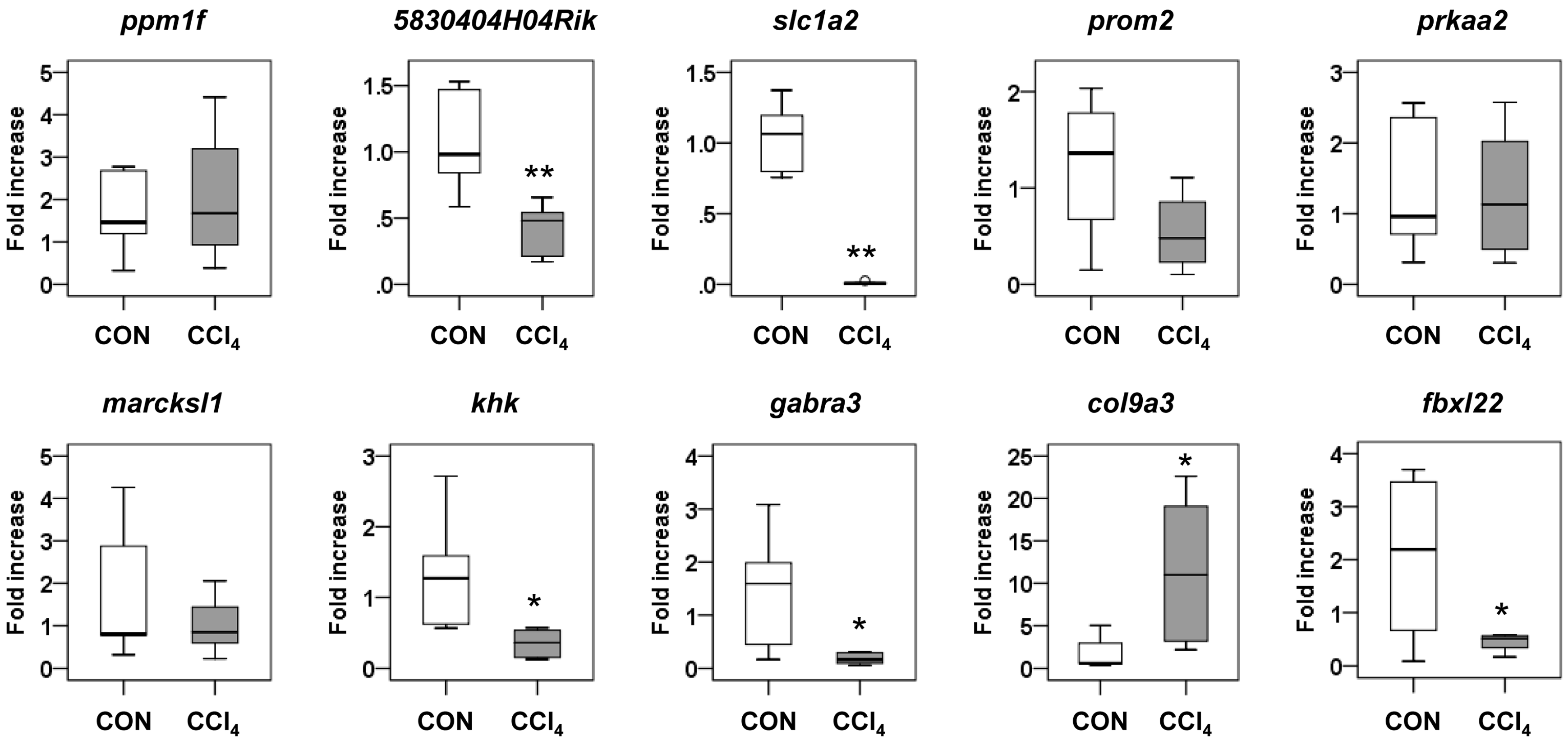
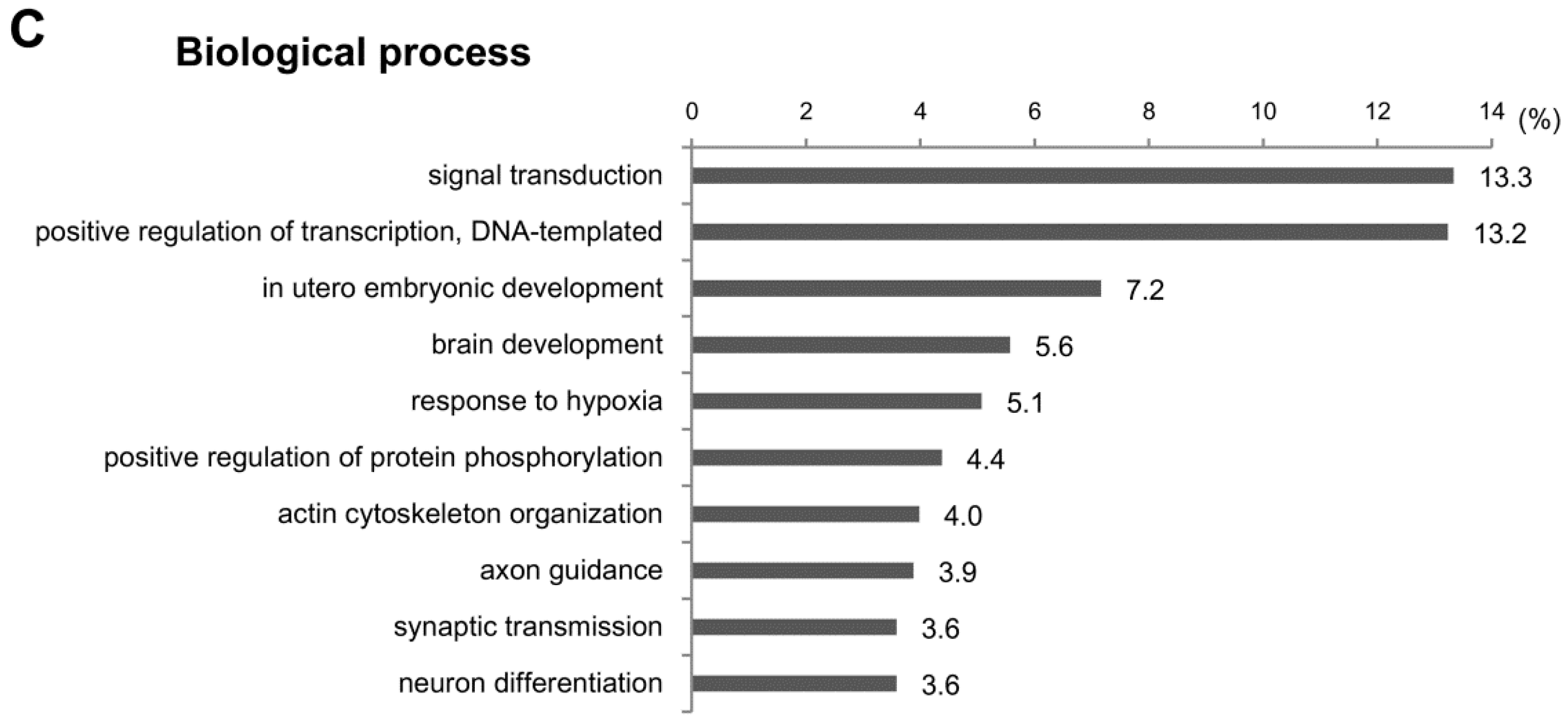
2.4. Functional Analysis of the Predicted Target Genes
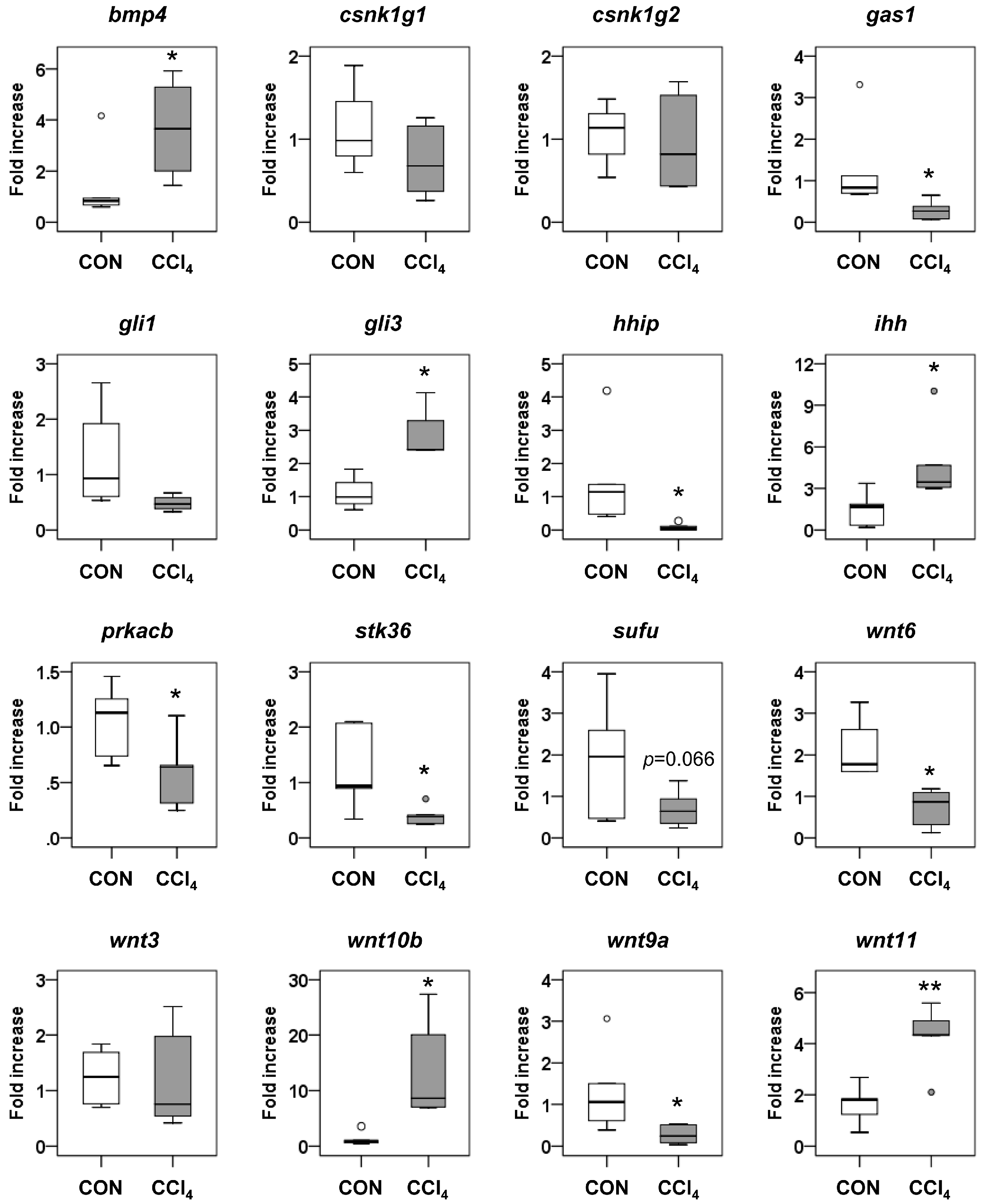
2.5. Interaction of miRNA Target Involved in Hedgehog Signaling
3. Discussion
4. Materials and Methods
4.1. Experimental Animal Model
4.2. Hematoxylin-Eosin and Sirius Red Staining
4.3. RNA Isolation and MicroRNA Microarray
4.4. Real-Time Quantitative Reverse Transcriptional Polymerase Chain Reaction Analysis
4.5. Statistical Analysis
4.6. MicroRNA Target Prediction
4.7. Gene Ontology and Kyoto Encyclopedia of Genes and Genomes Enrichment Analyses
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| HBV/HCV | hepatitis B/C virus |
| NASH | non-alcoholic steatohepatitis |
| miRNA | microRNAs |
| HSC | hepatic stellate cells |
| TGF | transforming growth factor |
| CCl4 | carbon-tetrachloride |
| GO | gene ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| H&E | hematoxylin-eosin stain |
| qRT-PCR | quantitative reverse transcriptional polymerase chain reaction |
| C2cd2 | C2 calcium-dependent domain containing 2 |
| Ppm1f | protein phosphatase 1F |
| Slcla2 | solute carrier family 1 |
| Prom2 | prominin 2 |
| Prkaa2 | protein kinase, AMP-activated, α 2 catalytic subunit |
| Marcksl1 | MARCKS-like 1 |
| Khk | ketohexokinase |
| Gabra3 | γ-aminobutyric acid (GABA) A receptor, subunit α 3 |
| Col9a3 | collagen, type IX, α 3 |
| TCA | tricarboxylic acid |
| Fbxl22 | F-box and leucine-rich repeat protein 22 |
| Hh | hedgehog |
| TNF | tumor necrosis factor |
| Bmp4 | bone morphogenetic protein 4 |
| Csnk1g1 | casein kinase 1 γ 1 |
| Gas1 | growth arrest specific 1 |
| Gli1 | GLI-Krüppel family member 1 |
| Hhip | Hh-interacting protein |
| Ihh | Indian hedgehog |
| cAMP | cyclic adenosine monophosphate |
| Prkacb | cAMP-dependent protein kinase catalytic subunit β |
| Stk36 | serine/threonine kinase 36 |
| Sufu | suppressor of fused homolog |
| MMTV | mouse mammary tumor virus |
| Wnt | wingless-type MMTV integration site family |
| MAPK | mitogen-activated protein kinase |
| mTOR | mechanistic target of rapamycin |
| HIF | hypoxia-inducible factor |
| FASN | fatty acid synthase |
| HMGCR | 3-hydroxy-3-methylglutaryl-CoA reductase |
| MMP9 | matrix metallopeptidase 9 |
| DMN | dimethylnitrosamine |
| MCD | methionine/choline-deficient |
| NBF | neutral buffered formalin |
| RNU1A | U1A small nuclear RNA |
| SEM | standard error of the mean |
| GO | gene ontology |
References
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Kim, V.N. MicroRNA biogenesis: Coordinated cropping and dicing. Nat. Rev. Mol. Cell Biol. 2005, 6, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-M.; Nguyen, D.T.; Lu, L.-F. Progress and challenge of microRNA research in immunity. Front. Genet. 2014, 5, 1–8. [Google Scholar]
- Shenoy, A.; Blelloch, R.H. Regulation of microRNA function in somatic stem cell proliferation and differentiation. Nat. Rev. Mol. Cell Biol. 2014, 15, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Bala, S.; Petrasek, J.; Mundkur, S.; Catalano, D.; Levin, I.; Ward, J.; Alao, H.; Kodys, K.; Szabo, G. Circulating microRNAs in exosomes indicate hepatocyte injury and inflammation in alcoholic, drug-induced, and inflammatory liver diseases. Hepatology 2012, 56, 1946–1957. [Google Scholar] [CrossRef] [PubMed]
- Szabo, G.; Bala, S. MicroRNAs in liver disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Tsitsiou, E.; Herrick, S.E.; Lindsay, M.A. MicroRNAs and the regulation of fibrosis. FEBS J. 2010, 277, 2015–2021. [Google Scholar] [CrossRef] [PubMed]
- Bowen, T.; Jenkins, R.H.; Fraser, D.J. MicroRNAs, transforming growth factor β-1, and tissue fibrosis. J. Pathol. 2013, 229, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Tang, N.; Wu, K.; Dai, W.; Ye, C.; Shi, J.; Zhang, J.; Ning, B.; Zeng, X.; Lin, Y. miR-21 simultaneously regulates ERK1 signaling in HSC activation and hepatocyte EMT in hepatic fibrosis. PLoS ONE 2014, 9, e108005. [Google Scholar] [CrossRef] [PubMed]
- Roderburg, C.; Urban, G.W.; Bettermann, K.; Vucur, M.; Zimmermann, H.; Schmidt, S.; Janssen, J.; Koppe, C.; Knolle, P.; Castoldi, M. Micro-RNA profiling reveals a role for miR-29 in human and murine liver fibrosis. Hepatology 2011, 53, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Lakner, A.M.; Steuerwald, N.M.; Walling, T.L.; Ghosh, S.; Li, T.; McKillop, I.H.; Russo, M.W.; Bonkovsky, H.L.; Schrum, L.W. Inhibitory effects of microRNA 19b in hepatic stellate cell-mediated fibrogenesis. Hepatology 2012, 56, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, W.; Xu, R.; Nie, Y.; Cao, X.; Meng, J.; Xu, X.; Hu, S.; Zheng, Z. MicroRNA-24 regulates cardiac fibrosis after myocardial infarction. J. Cell. Mol. Med. 2012, 16, 2150–2160. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.T.; Burk, R.F. In Carbon tetrachloride hepatotoxicity: An example of free radical-mediated injury. Semin. Liver Dis. 1990, 10, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Aisaki, K.; Ikawa, Y.; Wake, K. Evidence of hepatocyte apoptosis in rat liver after the administration of carbon tetrachloride. Am. J. Pathol. 1998, 153, 515–525. [Google Scholar] [CrossRef]
- Hyun, J.; Wang, S.; Kim, J.; Rao, K.M.; Park, S.Y.; Chung, I.; Ha, C.-S.; Kim, S.-W.; Yun, Y.H.; Jung, Y. MicroRNA-378 limits activation of hepatic stellate cells and liver fibrosis by suppressing GLI3 expression. Nat. Commun. 2016, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pawson, T.; Nash, P. Protein–protein interactions define specificity in signal transduction. Genes Dev. 2000, 14, 1027–1047. [Google Scholar] [PubMed]
- Chen, Y.; Choi, S.S.; Michelotti, G.A.; Chan, I.S.; Swiderska-Syn, M.; Karaca, G.F.; Xie, G.; Moylan, C.A.; Garibaldi, F.; Premont, R. Hedgehog controls hepatic stellate cell fate by regulating metabolism. Gastroenterology 2012, 143, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Sethi, J.K.; Hotamisligil, G.S. In The role of TNFα in adipocyte metabolism, Semin. Cell Dev. Biol. 1999, 10, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, Y.; Mao, H.; Fleig, S.; Omenetti, A.; Brown, K.D.; Sicklick, J.K.; Li, Y.X.; Diehl, A.M. Sonic hedgehog is an autocrine viability factor for myofibroblastic hepatic stellate cells. J. Hepatol. 2008, 48, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Tada, M.; Kanai, F.; Tanaka, Y.; Tateishi, K.; Ohta, M.; Asaoka, Y.; Seto, M.; Muroyama, R.; Fukai, K.; Imazeki, F. Down-regulation of hedgehog-interacting protein through genetic and epigenetic alterations in human hepatocellular carcinoma. Clin. Cancer Res. 2008, 14, 3768–3776. [Google Scholar] [CrossRef] [PubMed]
- Chuang, P.-T.; Kawcak, T.N.; McMahon, A.P. Feedback control of mammalian hedgehog signaling by the hedgehog-binding protein, HIP1, modulates FGF signaling during branching morphogenesis of the lung. Genes Dev. 2003, 17, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Gantier, M.P.; McCoy, C.E.; Rusinova, I.; Saulep, D.; Wang, D.; Xu, D.; Irving, A.T.; Behlke, M.A.; Hertzog, P.J.; Mackay, F. Analysis of microRNA turnover in mammalian cells following DICER1 ablation. Nucleic Acids Res. 2011, 39, 5692–5703. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Esau, C.; Davis, S.; Murray, S.F.; Yu, X.X.; Pandey, S.K.; Pear, M.; Watts, L.; Booten, S.L.; Graham, M.; McKay, R. MiR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab. 2006, 3, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Kota, J.; Chivukula, R.R.; O'Donnell, K.A.; Wentzel, E.A.; Montgomery, C.L.; Hwang, H.-W.; Chang, T.-C.; Vivekanandan, P.; Torbenson, M.; Clark, K.R. Therapeutic microRNA delivery suppresses tumorigenesis in a murine liver cancer model. Cell 2009, 137, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Dong, B.; Tian, Y.; Lefebvre, P.; Meng, Z.; Wang, X.; Pattou, F.; Han, W.; Lou, F.; Jove, R. MicroRNA-26a regulates insulin sensitivity and metabolism of glucose and lipids. J. Clin. Investig. 2015, 125, 2497–2509. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pan, Y.; Kan, M.; Xiao, X.; Wang, Y.; Guan, F.; Zhang, X.; Chen, L. Hepatoprotective effects of berberine on liver fibrosis via activation of amp-activated protein kinase. Life Sci. 2014, 98, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Adachi, M.; Osawa, Y.; Uchinami, H.; Kitamura, T.; Accili, D.; Brenner, D.A. The forkhead transcription factor FoxO1 regulates proliferation and transdifferentiation of hepatic stellate cells. Gastroenterology 2007, 132, 1434–1446. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Du, G.; Xu, Y.; Li, X.; Fan, W.; Chen, J.; Liu, C.; Chen, G.; Liu, C.; Zern, M.A. Inhibition of notch signaling pathway prevents cholestatic liver fibrosis by decreasing the differentiation of hepatic progenitor cells into cholangiocytes. Lab. Investig. 2016, 96, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Xue, R.; Tang, W.; Wu, W.; Hu, T.; Liu, X.; Peng, X.; Gu, J.; Chen, S.; Zhang, S. Toll like receptor 2 knock-out attenuates carbon tetrachloride (CCl4)-induced liver fibrosis by downregulating MAPK and NF-κB signaling pathways. FEBS Lett. 2014, 588, 2095–2100. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Yuko, K.; Sato, Y.; Ikeda, H.; Nakanuma, Y. Significance of oestrogen-related receptor γ on biliary epithelial cells in the pathogenesis of primary biliary cirrhosis. J. Clin. Pathol. 2014, 67, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Kodama, T.; Takehara, T.; Hikita, H.; Shimizu, S.; Shigekawa, M.; Tsunematsu, H.; Li, W.; Miyagi, T.; Hosui, A.; Tatsumi, T. Increases in p53 expression induce CTGF synthesis by mouse and human hepatocytes and result in liver fibrosis in mice. J. Clin. Investig. 2011, 121, 3343–3356. [Google Scholar] [CrossRef] [PubMed]
- Philips, G.M.; Chan, I.S.; Swiderska, M.; Schroder, V.T.; Guy, C.; Karaca, G.F.; Moylan, C.; Venkatraman, T.; Feuerlein, S.; Syn, W.-K. Hedgehog signaling antagonist promotes regression of both liver fibrosis and hepatocellular carcinoma in a murine model of primary liver cancer. PLoS ONE 2011, 6, e23943. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, X.; Xu, W.; Wang, S.; Hu, Z.; Zhang, Q.; Deng, X.; Wang, J.; Zhang, J.; Guo, C. Antifibrotic effects of luteolin on hepatic stellate cells and liver fibrosis by targeting AKT/mTOR/p70S6K and TGFβ/Smad signalling pathways. Liver Int. 2015, 35, 1222–1233. [Google Scholar] [CrossRef] [PubMed]
- Lok, A. Hepatitis B: Liver fibrosis and hepatocellular carcinoma. Gastroenterol. Clin. Biol. 2009, 33, 911–915. [Google Scholar] [CrossRef] [PubMed]
- Son, G.; Hines, I.N.; Lindquist, J.; Schrum, L.W.; Rippe, R.A. Inhibition of phosphatidylinositol 3-kinase signaling in hepatic stellate cells blocks the progression of hepatic fibrosis. Hepatology 2009, 50, 1512–1523. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, H.; Lv, X.; Wang, Q.; Zhao, H.; Yang, F.; Yang, Y.; Li, J. Involvement of camp-PKA pathway in adenosine A 1 and A 2A receptor-mediated regulation of acetaldehyde-induced activation of HSCs. Biochimie 2015, 115, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Gressner, A.M.; Weiskirchen, R.; Breitkopf, K.; Dooley, S. Roles of TGF-β in hepatic fibrosis. Front. Biosci. 2002, 7, d793–d807. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.-O.; Welch, T.P.; Gonzalez, F.J.; Copple, B.L. Reduced liver fibrosis in hypoxia-inducible factor-1α-deficient mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G582–G592. [Google Scholar] [CrossRef] [PubMed]
- Marchesini, G.; Bugianesi, E.; Forlani, G.; Cerrelli, F.; Lenzi, M.; Manini, R.; Natale, S.; Vanni, E.; Villanova, N.; Melchionda, N. Nonalcoholic fatty liver, steatohepatitis, and the metabolic syndrome. Hepatology 2003, 37, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Zhong, Y.; Dang, L.; Zhu, M.; Yu, H.; Chen, W.; Cui, J.; Bian, H.; Li, Z. Alteration of protein glycosylation in human hepatic stellate cells activated with transforming growth factor-β1. J. Proteom. 2012, 75, 4114–4123. [Google Scholar] [CrossRef] [PubMed]
- Blomme, B.; Van Steenkiste, C.; Callewaert, N.; van Vlierberghe, H. Alteration of protein glycosylation in liver diseases. J. Hepatol. 2009, 50, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Tsukada, S.; Parsons, C.J.; Rippe, R.A. Mechanisms of liver fibrosis. Clin. Chim. Acta 2006, 364, 33–60. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zhu, M.; Qin, Y.; Zhong, Y.; Yan, H.; Wang, Q.; Bian, H.; Li, Z. Analysis of glycan-related genes expression and glycan profiles in mice with liver fibrosis. J. Proteome Res. 2012, 11, 5277–5285. [Google Scholar] [CrossRef] [PubMed]
- Lian, N.; Jiang, Y.; Zhang, F.; Jin, H.; Lu, C.; Wu, X.; Lu, Y.; Zheng, S. Curcumin regulates cell fate and metabolism by inhibiting hedgehog signaling in hepatic stellate cells. Lab. Investig. 2015, 95, 790–803. [Google Scholar] [CrossRef] [PubMed]
- Bishop, B.; Aricescu, A.R.; Harlos, K.; O'Callaghan, C.A.; Jones, E.Y.; Siebold, C. Structural insights into hedgehog ligand sequestration by the human hedgehog-interacting protein hhip. Nat. Struct. Mol. Biol. 2009, 16, 698–703. [Google Scholar] [CrossRef] [PubMed]
- Chuang, P.-T.; McMahon, A.P. Vertebrate hedgehog signalling modulated by induction of a hedgehog-binding protein. Nature 1999, 397, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Omenetti, A.; Choi, S.; Michelotti, G.; Diehl, A.M. Hedgehog signaling in the liver. J. Hepatol. 2011, 54, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.J.; Johnson, R.L.; Burke, A.C.; Nelson, C.E.; Morgan, B.A.; Tabin, C. Sonic hedgehog is an endodermal signal inducing Bmp-4 and Hox genes during induction and regionalization of the chick hindgut. Development 1995, 121, 3163–3174. [Google Scholar] [PubMed]
- Astorga, J.; Carlsson, P. Hedgehog induction of murine vasculogenesis is mediated by Foxf1 and Bmp4. Development 2007, 134, 3753–3761. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Shen, H.; Sun, Y.; Li, P.; Burczynski, F.; Namaka, M.; Gong, Y. Bone morphogenetic protein 4 mediates bile duct ligation induced liver fibrosis through activation of SMAD1 and ERK1/2 in rat hepatic stellate cells. J. Cell. Physiol. 2006, 207, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.H.; She, H.; Han, Y.-P.; Wang, J.; Xiong, S.; Asahina, K.; Tsukamoto, H. Wnt antagonism inhibits hepatic stellate cell activation and liver fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G39–G49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Cai, Y.; Soofi, A.; Dressler, G.R. Activation of Wnt11 by transforming growth factor-β drives mesenchymal gene expression through non-canonical wnt protein signaling in renal epithelial cells. J. Biol. Chem. 2012, 287, 21290–21302. [Google Scholar] [CrossRef] [PubMed]
- Beaton, H.; Andrews, D.; Parsons, M.; Murphy, M.; Gaffney, A.; Kavanagh, D.; McKay, G.J.; Maxwell, A.P.; Taylor, C.T.; Cummins, E.P. Wnt6 regulates epithelial cell differentiation and is dysregulated in renal fibrosis. Am. J. Physiol. Ren. Physiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Valinezhad Orang, A.; Safaralizadeh, R.; Kazemzadeh-Bavili, M. Mechanisms of miRNA-mediated gene regulation from common downregulation to mRNA-specific upregulation. Int. J. Genom. 2014, 2014, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S. Posttranscriptional upregulation by microRNAs. Wiley Interdiscip. Rev. RNA 2012, 3, 311–330. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Yasuda, T.; Saigo, K.; Urashima, T.; Toyoda, H.; Okanoue, T.; Shimotohno, K. Comprehensive analysis of microRNA expression patterns in hepatocellular carcinoma and non-tumorous tissues. Oncogene 2005, 25, 2537–2545. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Chen, Y.P.; Kong, M.; Zheng, L.; Yang, Y.D.; Li, Y.M. Transition from hepatic steatosis to steatohepatitis: Unique microRNA patterns and potential downstream functions and pathways. J. Gastroenterol. Hepatol. 2012, 27, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.; Luo, D.; Rong, M.; Chen, G. Underexpression of miR-34a in hepatocellular carcinoma and its contribution towards enhancement of proliferating inhibitory effects of agents targeting c-MET. PLoS ONE 2013, 8, e61054. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Sun, Y.; Dong, R.; Yang, S.; Pan, C.; Xiang, D.; Miao, M.; Jiao, B. miR-34a is upregulated during liver regeneration in rats and is associated with the suppression of hepatocyte proliferation. PLoS ONE 2011, 6, e20238. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Glaser, S.S.; Francis, H.; Yang, F.; Han, Y.; Stokes, A.; Staloch, D.; McCarra, J.; Liu, J.; Venter, J. Epigenetic regulation of miR-34a expression in alcoholic liver injury. Am. J. Pathol. 2012, 181, 804–817. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Q.; Chen, C.; Xu, M.D.; Guo, J.; Li, Y.M.; Xia, Q.M.; Liu, H.M.; He, J.; Yu, H.Y.; Zhu, L. The rno-miR-34 family is upregulated and targets ACSL1 in dimethylnitrosamine-induced hepatic fibrosis in rats. FEBS J. 2011, 278, 1522–1532. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Niu, X.; Wang, Y.; Kong, L.; Wang, R.; Zhang, Y.; Zhao, S.; Nan, Y. miR-146a-5p suppresses activation and proliferation of hepatic stellate cells in nonalcoholic fibrosing steatohepatitis through directly targeting WNT1 and WNT5A. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, C.; Wang, M.; Su, L.; Qu, Y.; Li, J.; Yu, B.; Yan, M.; Yu, Y.; Liu, B. Decrease of miR-202–3p expression, a novel tumor suppressor, in gastric cancer. PLoS ONE 2013, 8, e69756. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Benz, F.; Vargas Cardenas, D.; Vucur, M.; Gautheron, J.; Schneider, A.; Hellerbrand, C.; Pottier, N.; Alder, J.; Tacke, F. miR-30c and miR-193 are a part of the TGF-β-dependent regulatory network controlling extracellular matrix genes in liver fibrosis. J. Dig. Dis. 2015, 16, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Mirbase. Available online: http://www.mirbase.org/ (accessed on 1 November 2014).
- Kim, J.; Wang, S.; Hyun, J.; Choi, S.S.; Cha, H.; Ock, M.; Jung, Y. Hepatic stellate cells express thymosin β4 in chronically damaged liver. PLoS ONE 2015, 10, e0122758. [Google Scholar]
- Bioconductor. Available online: http://www.bioconductor.org/ (accessed on 1 November 2014).
- R project. Available online: https://www.r-project.org/ (accessed on 1 November 2014).
- Dweep, H.; Sticht, C.; Pandey, P.; Gretz, N. Mirwalk–database: Prediction of possible miRNA binding sites by “walking” the genes of three genomes. J. Biomed. Inform. 2011, 44, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Cytoscape. Available online: http://cytoscape.org/ (accessed on 1 November 2014).
- Uniprot. Available online: http://www.uniprot.org/ (accessed on 1 November 2014).
- GNU Wget 1.18 Manual. Available online: https://www.gnu.org/software/wget/ (accessed on 1 November 2014).
- Qureshi, R.; Sacan, A. Weighted set enrichment of gene expression data. BMC Syst. Biol. 2013, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe ID | Log2 Change (CCl4/Corn Oil) | Fold Change (CCl4/Corn Oil) | Adjust p-Value |
|---|---|---|---|
| Upregulated miRNAs | |||
| mmu-miR-574-5p | 3.625210638 | 12.33949 | 0.000259147 |
| mmu-miR-466i-5p | 3.084363793 | 8.481761 | 0.000830411 |
| mmu-miR-342-3p | 1.180557713 | 2.266644 | 0.003273323 |
| mmu-let-7i-5p | 1.136496066 | 2.198464 | 0.016247731 |
| mmu-miR-34a-5p | 1.871455739 | 3.659016 | 0.032876548 |
| mmu-miR-188-5p | 2.700779126 | 6.501529 | 0.042779303 |
| mmu-miR-5119 | 1.048052536 | 2.067737 | 0.042779303 |
| Downregulated miRNAs | |||
| mmu-miR-378a-3p | −1.338779005 | 0.395355 | 0.00162313 |
| mmu-miR-202-3p | −1.079114551 | 0.473319 | 0.003273323 |
| mmu-miR-378b | −1.360233472 | 0.389519 | 0.003273323 |
| mmu-miR-378d | −1.424843795 | 0.37246 | 0.003273323 |
| mmu-miR-212-3p | −1.132709203 | 0.456058 | 0.018962762 |
| KEGG ID | Pathway | p-Value | KEGG ID | Pathway | p-Value |
|---|---|---|---|---|---|
| mmu01100 | Metabolic pathways | 8.71 × 10−8 | mmu04020 | Calcium signaling pathway | 3.75 × 10−3 |
| mmu04152 | AMPK signaling pathway | 6.75 × 10−6 | mmu00071 | Fatty acid degradation | 4.23 × 10−3 |
| mmu04920 | Adipocytokine signaling pathway | 4.78 × 10−5 | mmu04022 | cGMP-PKG signaling pathway | 4.62 × 10−3 |
| mmu05200 | Pathways in cancer | 1.90 × 10−4 | mmu04915 | Estrogen signaling pathway | 6.78 × 10−3 |
| mmu00514 | Other types of O-glycan biosynthesis | 3.13 × 10−4 | mmu00601 | Glycosphingolipid biosynthesis: lacto and neolacto series | 7.17 × 10−3 |
| mmu00600 | Sphingolipid metabolism | 4.13 × 10−4 | mmu00561 | Glycerolipid metabolism | 7.21 × 10−3 |
| mmu00533 | Glycosaminoglycan biosynthesis: keratan sulfate | 7.12 × 10−4 | mmu04115 | p53 signaling pathway | 9.55 × 10−3 |
| mmu04910 | Insulin signaling pathway | 9.09 × 10−4 | mmu04666 | Fc γ R-mediated phagocytosis | 9.57 × 10−3 |
| mmu00310 | Lysine degradation | 1.04 × 10−3 | mmu04014 | Ras signaling pathway | 1.00 × 10−2 |
| mmu04068 | FoxO signaling pathway | 1.08 × 10−3 | mmu04725 | Cholinergic synapse | 1.01 × 10−2 |
| mmu04974 | Protein digestion and absorption | 1.34 × 10−3 | mmu04340 | Hedgehog signaling pathway | 1.02 × 10−2 |
| mmu04210 | Apoptosis | 1.53 × 10−3 | mmu04668 | TNF signaling pathway | 1.03 × 10−2 |
| mmu00510 | N-Glycan biosynthesis | 1.62 × 10−3 | mmu04660 | T cell receptor signaling pathway | 1.05 × 10−2 |
| mmu04330 | Notch signaling pathway | 1.62 × 10−3 | mmu04150 | mTOR signaling pathway | 1.12 × 10−2 |
| mmu04080 | Neuroactive ligand-receptor interaction | 1.83 × 10−3 | mmu04142 | Lysosome | 1.25 × 10−2 |
| mmu04611 | Platelet activation | 2.06 × 10−3 | mmu04550 | Signaling pathways regulating pluripotency of stem cells | 1.26 × 10−2 |
| mmu00061 | Fatty acid biosynthesis | 2.21 × 10−3 | mmu01212 | Fatty acid metabolism | 1.27 × 10−2 |
| mmu04913 | Ovarian steroidogenesis | 2.56 × 10−3 | mmu05340 | Primary immunodeficiency | 1.28 × 10−2 |
| mmu04360 | Axon guidance | 2.84 × 10−3 | mmu00280 | Valine, leucine and isoleucine degradation | 1.31 × 10−2 |
| mmu04010 | MAPK signaling pathway | 3.34 × 10−3 | mmu00010 | Glycolysis/gluconeogenesis | 1.36 × 10−2 |
| RefSeq | Gene Name | Targeting miRNA | Other Involved Pathways |
|---|---|---|---|
| NM_007554 | Bmp4 | mmu-miR-378 | TGF-β signaling pathway, signaling pathways regulating pluripotency of stem cells, pathways in cancer, basal cell carcinoma |
| NM_173185 | Csnk1g1 | mmu-miR-202-3p | - |
| NM_134002 | Csnk1g2 | mmu-miR-378 | - |
| NM_008086 | Gas1 | mmu-miR-188-5p | - |
| NM_010296 | Gli1 | mmu-miR-188-5p | cAMP signaling pathway, pathways in cancer, basal cell carcinoma |
| NM_008130 | Gli3 | mmu-miR-378 | |
| NM_020259 | Hhip | mmu-miR-342-3p | |
| NM_010544 | Ihh | mmu-miR-34a | - |
| NM_011100 | Prkacb | mmu-miR-34a | MAPK signaling pathway, Ras signaling pathway, calcium signaling pathway, cAMP signaling pathway, apoptosis, adrenergic signaling in cardiomyocytes, platelet activation, cholinergic synapse, insulin signaling pathway, ovarian steroidogenesis, estrogen signaling pathway, melanogenesis, thyroid hormone synthesis, vasopressin-regulated water reabsorption, salivary secretion, bile secretion, pathways in cancer, proteoglycans in cancer, dilated cardiomyopathy |
| NM_175031 | Stk36 | mmu-let-7i | Pathways in cancer, basal cell carcinoma |
| NM_015752 | Sufu | mmu-miR-34a | |
| NM_009526 | Wnt6 | mmu-miR-378 | Signaling pathways regulating pluripotency of stem cells, melanogenesis, pathways in cancer, proteoglycans in cancer, basal cell carcinoma |
| NM_009522 | Wnt3a | mmu-let-7i | |
| NM_009521 | Wnt3 | mmu-miR-342-3p | |
| NM_011718 | Wnt10b | ||
| NM_139298 | Wnt9a | mmu-let-7i, mmu-miR-342-3p | |
| NM_009519 | Wnt11 | mmu-miR-188-5p |
| Gene | Forward (5’ to 3’) | Reverse (5’ to 3’) |
|---|---|---|
| Ppm1f | GCTTCTTCAACTGCCTTTGG | CCATGACCATCAAACACAGC |
| 5830404H04Rik | GCTGGCTCATTTTCTTCAGG | CGGAAGAAAAGCACCATCAT |
| Slc1a2 | TCTGAGGAGGCCAATACCAC | TTCATCCCGTCCTTGAACTC |
| Prom2 | GTGTGACATGATGGCTGACC | ACCCTGGGGATATGGAAAAG |
| Pkaa2 | CGGCGCCTTTCCTTGAATAT | GGCCTGTTCCTCACGGTATTA |
| Marcksl1 | GGCAGCCAGAGCTCTAAGG | TCACGTGGCCATTCTCCT |
| Khk | AGCCTCATGGAAGAGAAGCA | GGAGGTCATCCAGGACAAAA |
| Gabra3 | ATGTGGCACTTTTATGTGACCA | CCCCAGGTTCTTGTCGTCTTG |
| Col9a3 | CAAGGATGGCATTGATGGAG | CAGACCATCTACACCAGGCAGT |
| Fbxl22 | ATGGGCAAGCAGGTTAAATG | CACGCGAACAGAACAGAAAA |
| Bmp4 | GAGGAGGAGGAAGAGCAGAG | TGGGATGTTCTCCAGATGTT |
| Csnk1g1 | CAGTGGTTTTCACCTTCTGG | AGGCCTTCACCTGCACTG |
| Csnk1g2 | TACTACTTCGGCCCTTGTGG | GGCTTCACGTCACGGTAGAT |
| Gas1 | TAAATTGCTTGTGACCACTG | GAGCATTACCGATGGATAGA |
| Gli1 | TGTGTGAGCAAGAAGGTTGC | ATGGCTTCTCATTGGAGTGG |
| Gli3 | GCAACCTCACTCTGCAACAA | CCTTGTGCCTCCATTTTGAT |
| Hhip | GGTCACATCTTGGGATTTGG | TCCATTGTGAGTCTGGGTCA |
| Ihh | CAATCCCGACATCATCTTCA | AGTTCAGACGGTCCTTGCAG |
| Prkacb | TCTTTCCTGCGTCATCAGTG | AAGGGAGCACTGGTCAGAGA |
| Stk36 | CCAGGAGAGTAGCAGCATCC | TGTCTGCTCGGAATCATCTG |
| Sufu | TCCAGGTTACCGCTATCGTC | TCCACTGTTGGGCTGAATGT |
| Wnt6 | TGTCAGTTCCAGTTCCGTTTCC | GCTGCGGTGATTGCAAACA |
| Wnt3a | GGCTCCTCTCGGATACCTCT | GGGCATGATCTCCACGTAGT |
| Wnt3 | CAAGCACAACAATGAAGCAGG | TCGGGACTCACGGTGTTTC |
| Wnt10b | ACGACATGGACTTCGGAGAGAAGT | CATTCTCGCCTGGATGTCCC |
| Wnt9a | GGCACAGGGTTACAAACAAC | GGACAGAGGCAACTGAGAAA |
| Wnt11 | ATGTGCGGACAACCTCAGCTA | CGCATCAGTTTATTGGCTTGG |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyun, J.; Park, J.; Wang, S.; Kim, J.; Lee, H.-H.; Seo, Y.-S.; Jung, Y. MicroRNA Expression Profiling in CCl4-Induced Liver Fibrosis of Mus musculus. Int. J. Mol. Sci. 2016, 17, 961. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17060961
Hyun J, Park J, Wang S, Kim J, Lee H-H, Seo Y-S, Jung Y. MicroRNA Expression Profiling in CCl4-Induced Liver Fibrosis of Mus musculus. International Journal of Molecular Sciences. 2016; 17(6):961. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17060961
Chicago/Turabian StyleHyun, Jeongeun, Jungwook Park, Sihyung Wang, Jieun Kim, Hyun-Hee Lee, Young-Su Seo, and Youngmi Jung. 2016. "MicroRNA Expression Profiling in CCl4-Induced Liver Fibrosis of Mus musculus" International Journal of Molecular Sciences 17, no. 6: 961. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17060961