
Salvianolic Acid A, as a Novel ETA Receptor Antagonist, Shows Inhibitory Effects on Tumor in Vitro

Abstract
:1. Introduction
2. Results
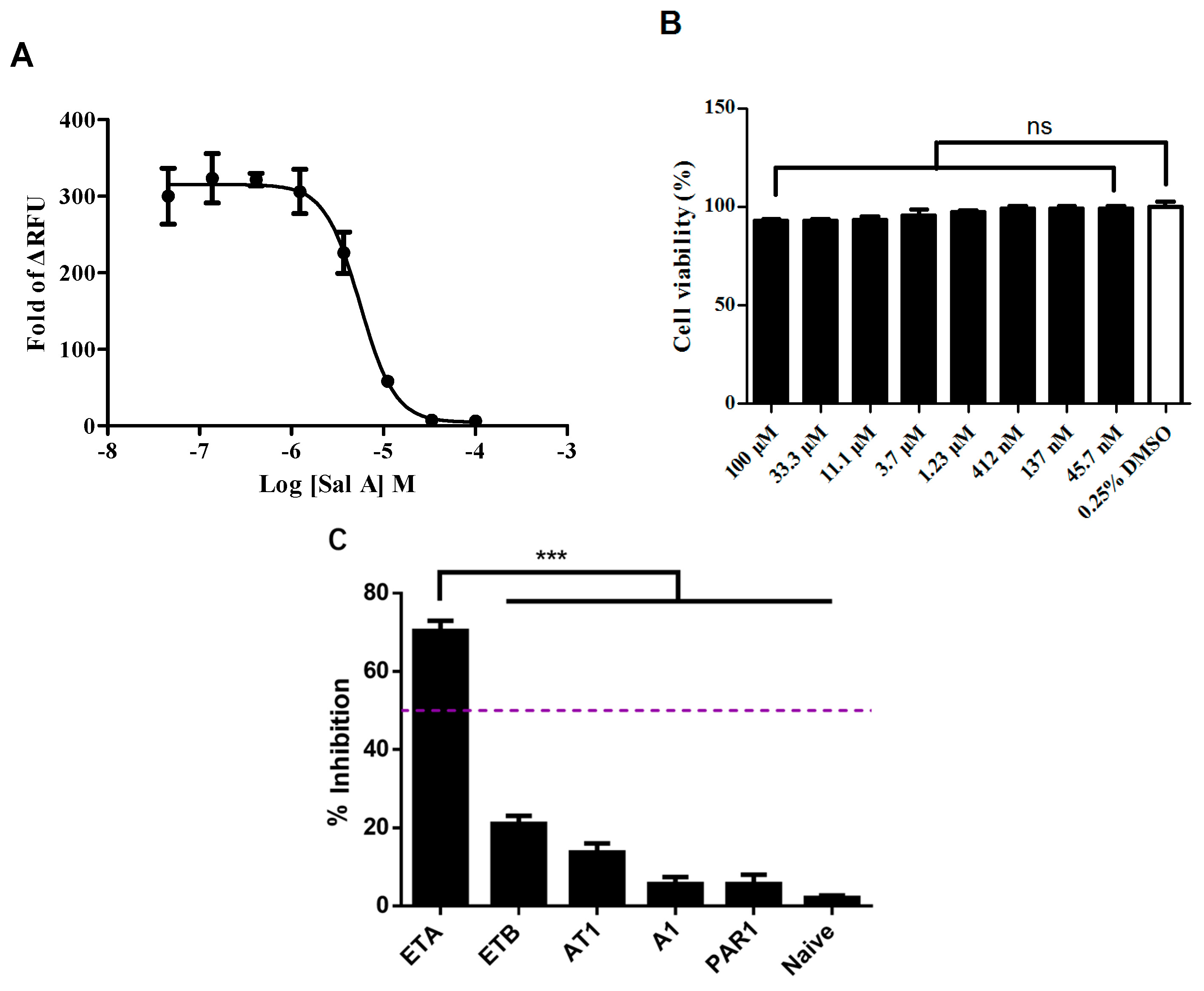
2.1. HEK293/Endothelin A Receptor (ETAR) Cell Line Validation
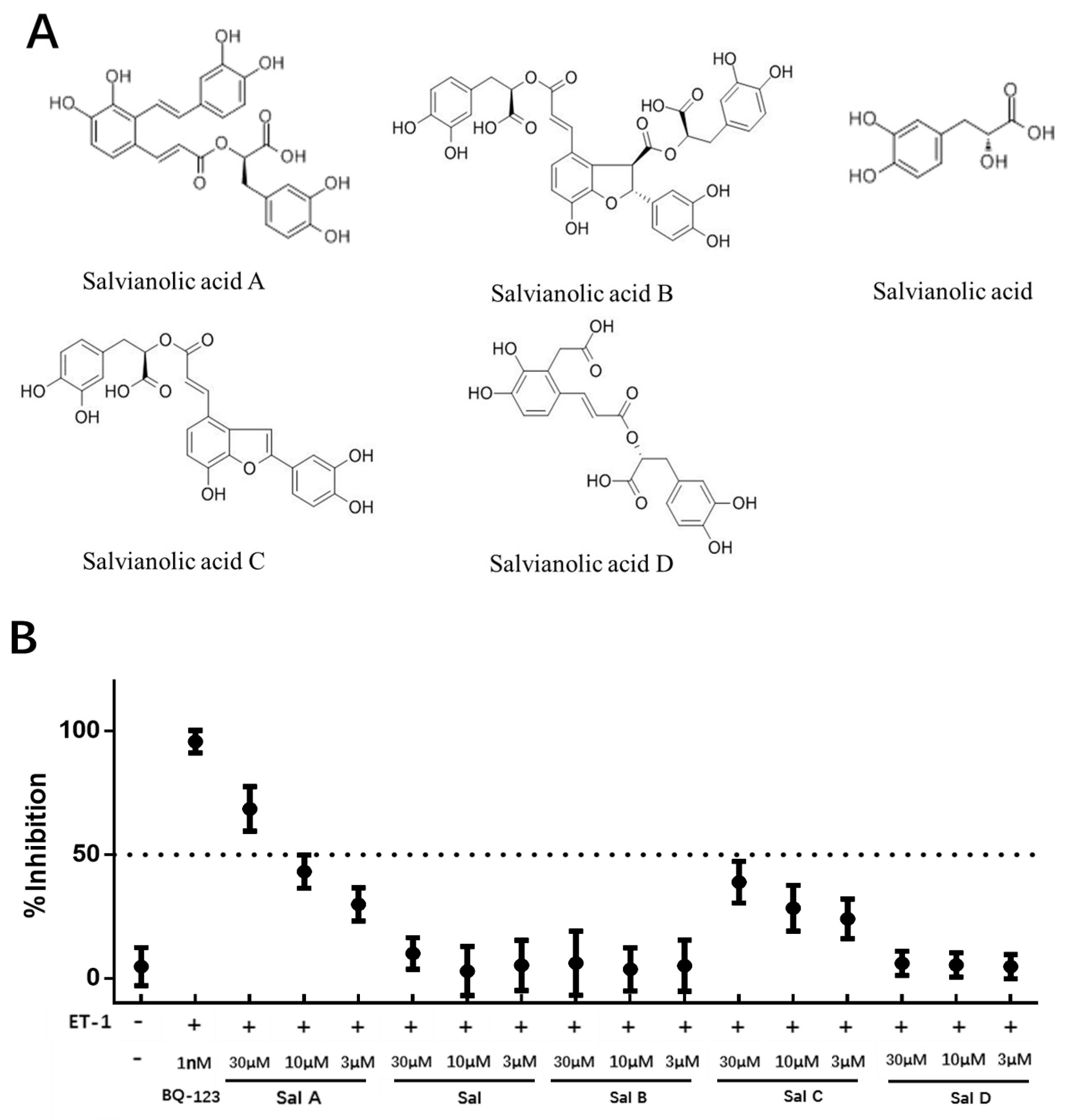
2.2. ETAR Antagonist Primary Screening
2.3. Salvianolic Acid A (Sal A) Dose-Dependently Blocked Exogenous ETAR without Inducing Cytotoxicity in the HEK293/ETAR Cell Line
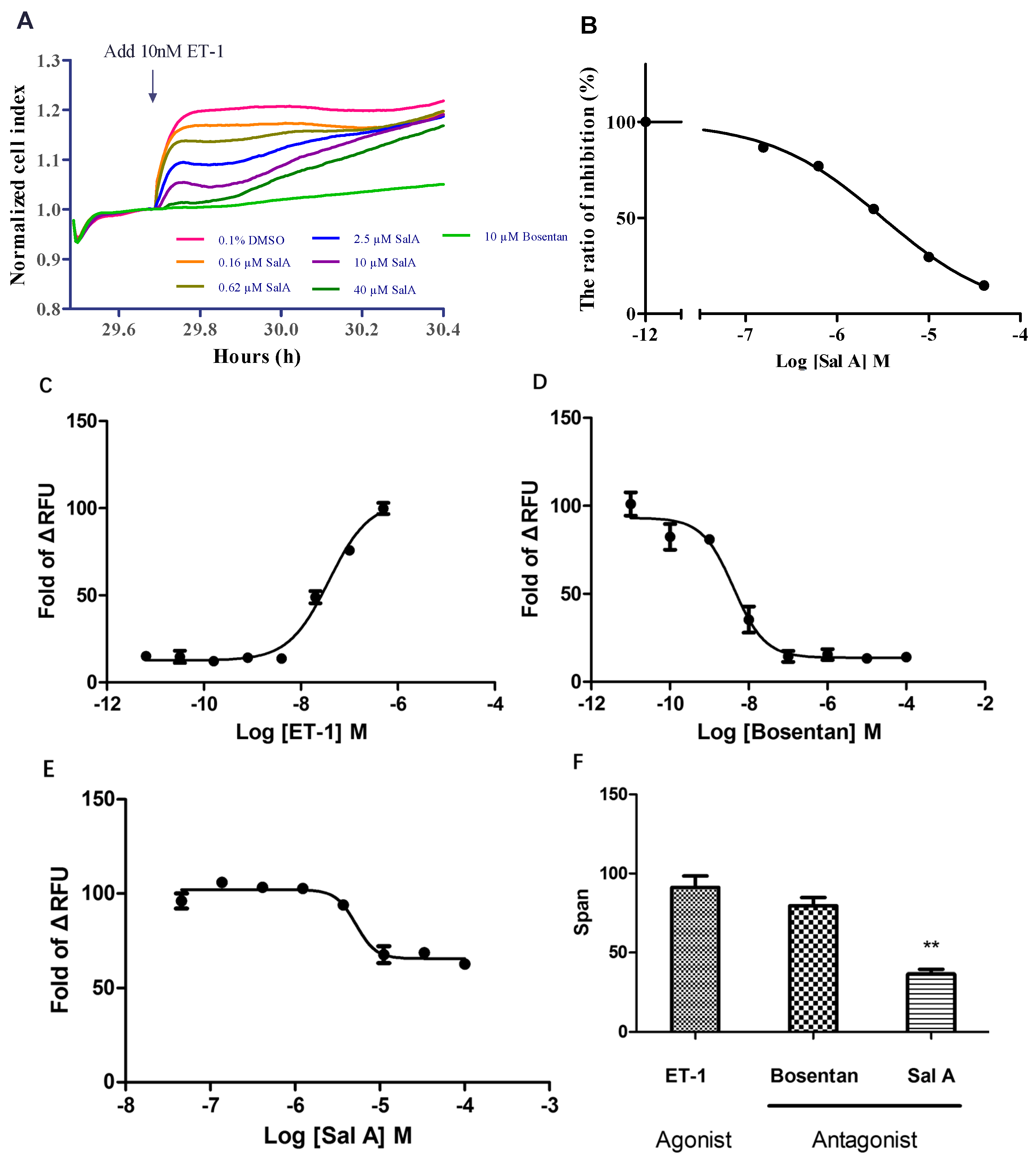
2.4. Sal A Inhibited Endogenously Expressed ETAR in HeLa Cells
2.5. Sal A Suppressed Proliferations in Multiple Cancer Cell Lines
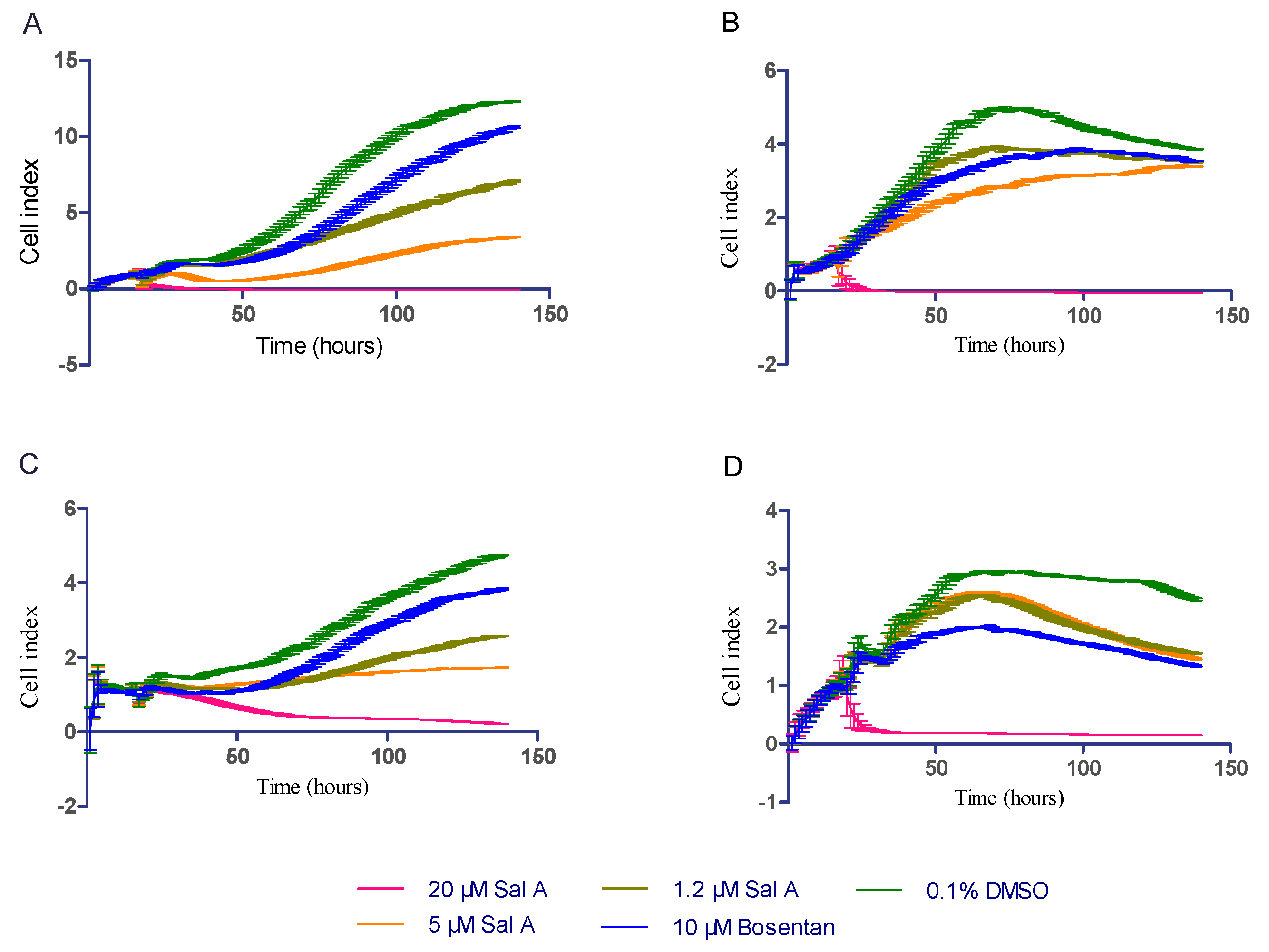
2.6. Sal A Extended Cell Lins Proliferation Cycle by Real-Time Cell Analysis (RTCA)
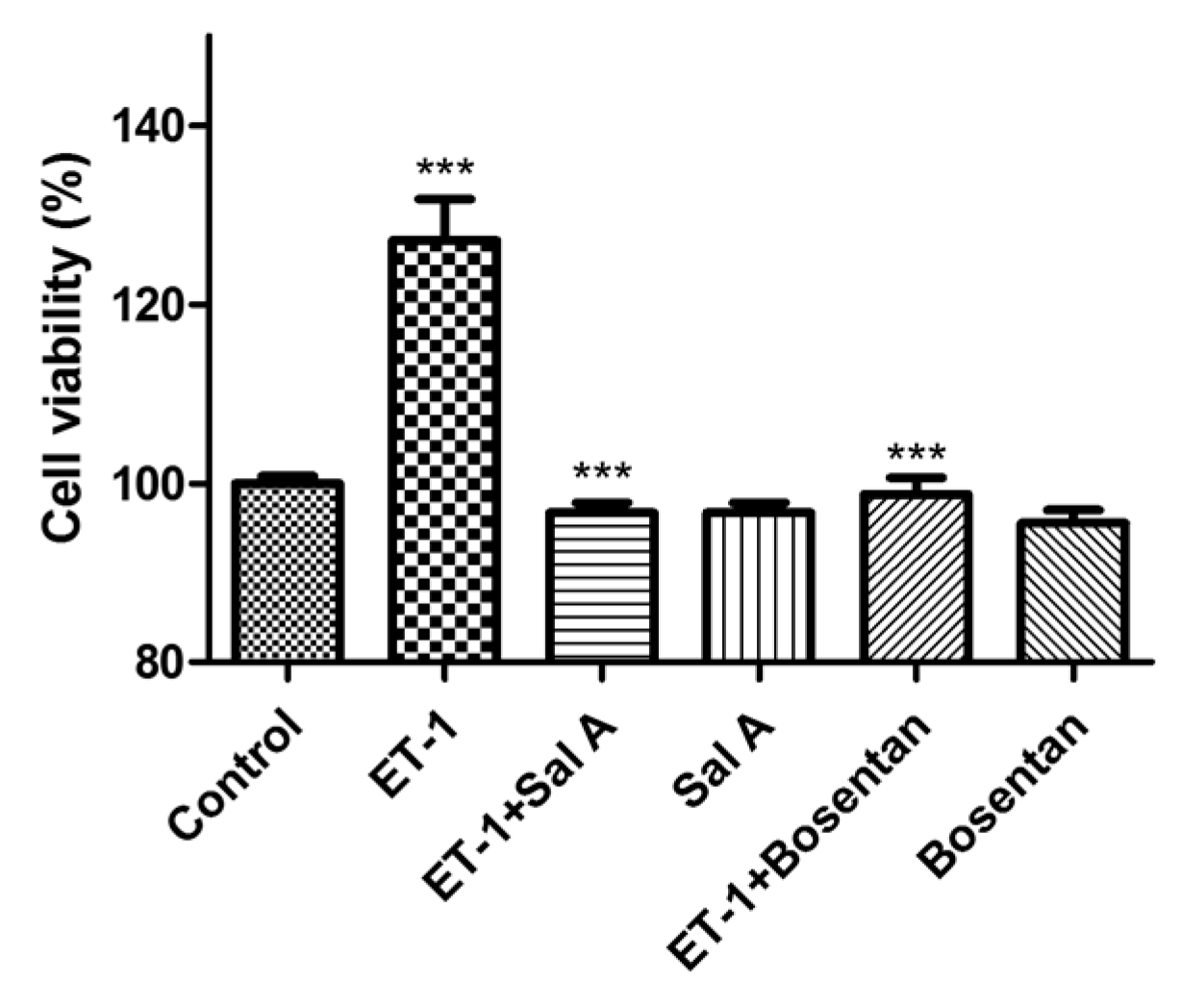
2.7. Sal A Inhibited the Proliferation Induced by Exogenous ET-1 in DU145 Cells
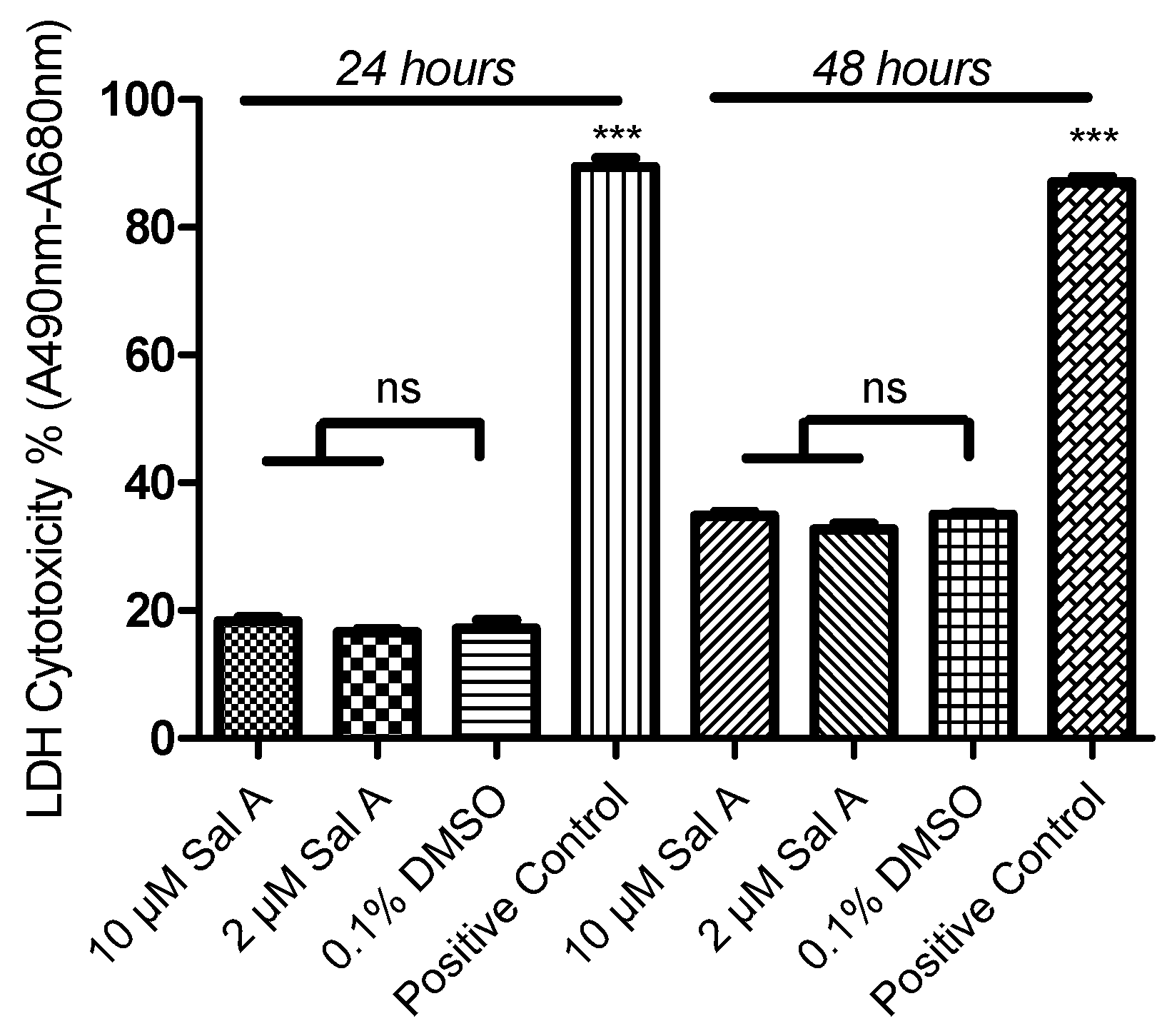
2.8. Effect of Sal A on Cell Cytotoxicity in HUVEC
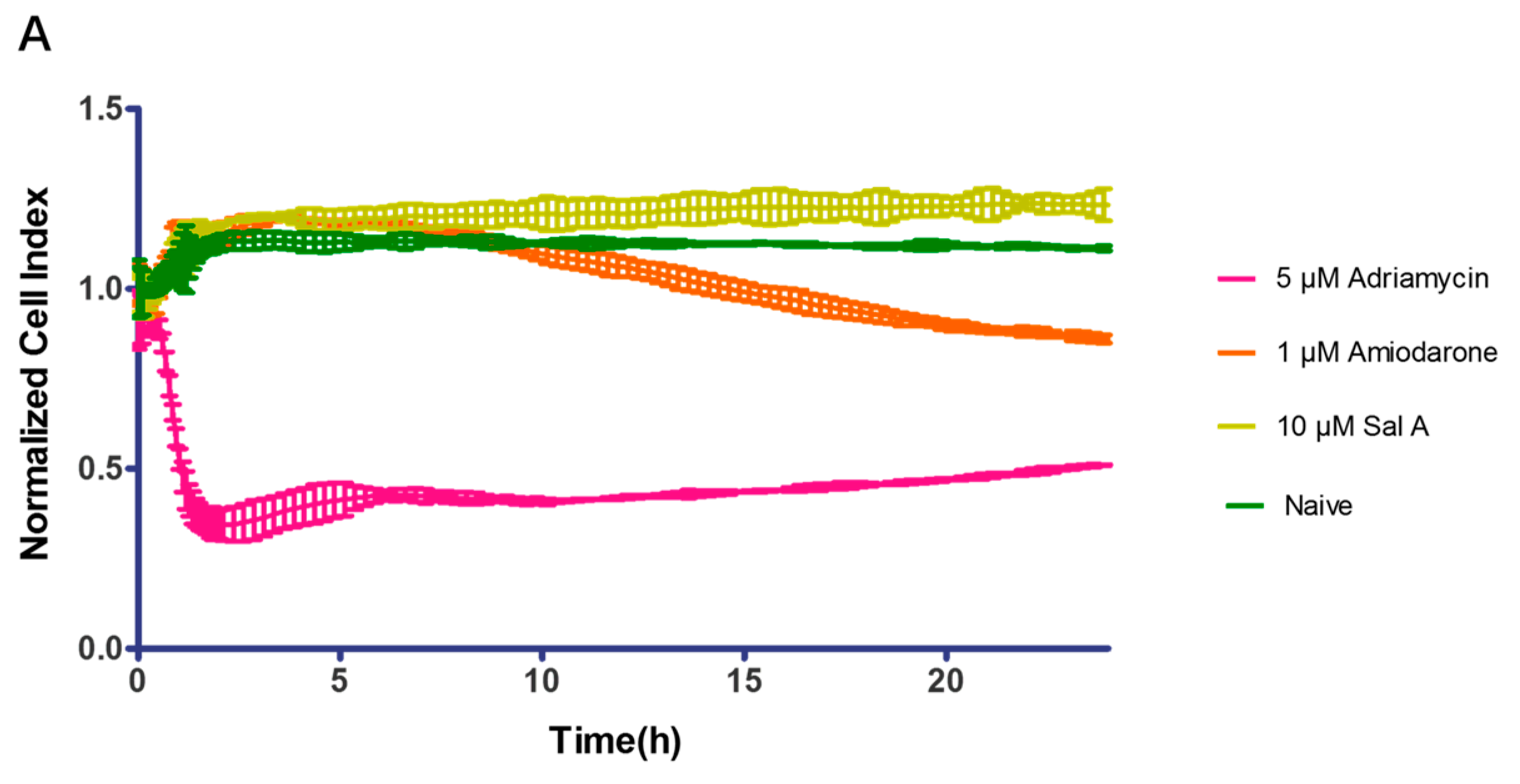
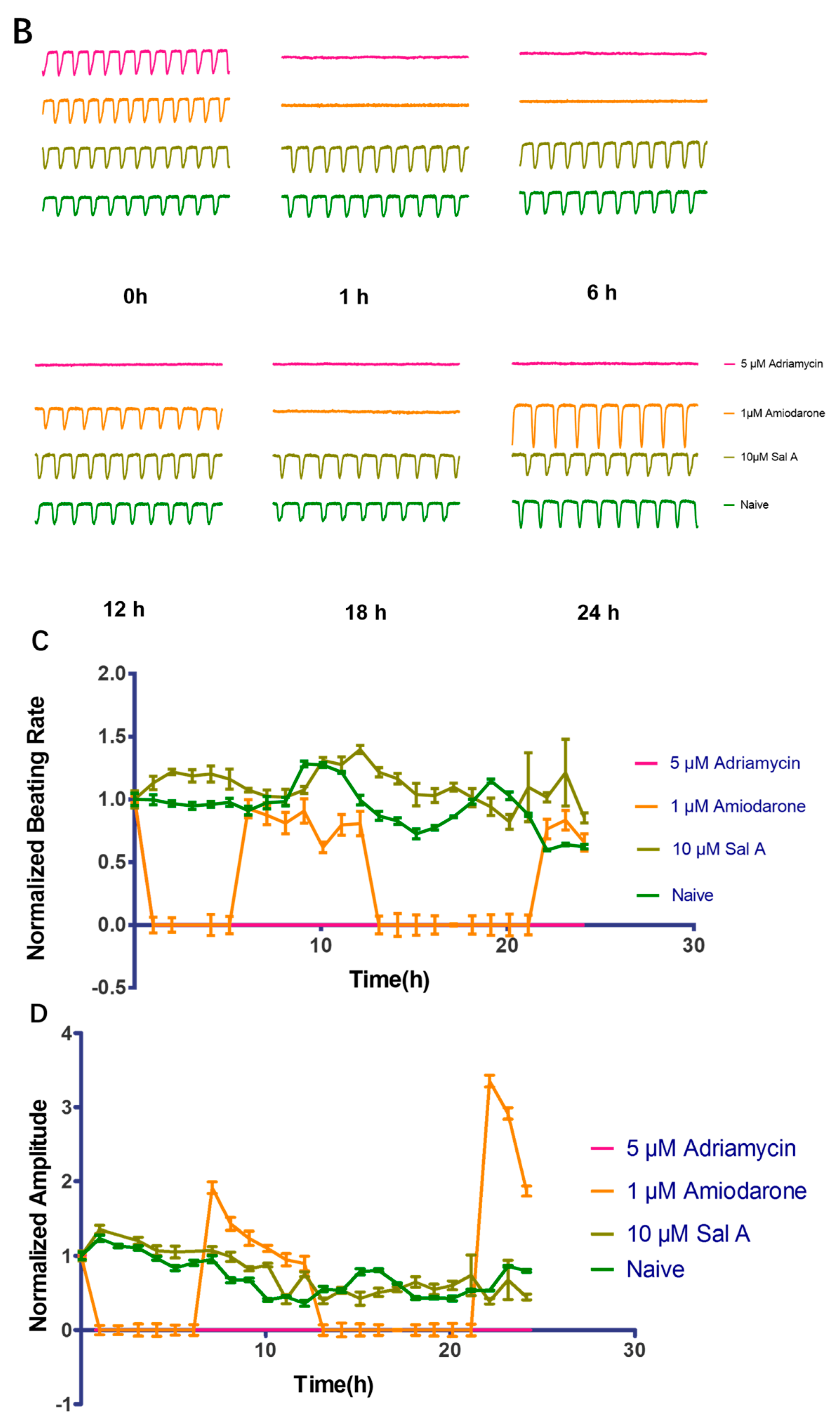
2.9. Effect of Sal A on Cardio-Toxicity in hiPS-CMs
3. Discussion
4. Materials and Methods
4.1. Reagents and Materials
4.2. Compund Preparation
4.3. Cell Culture
4.4. Antagonist Screening Assay
4.4.1. Ca2+ Influx Assay
4.4.2. Real-Time Cell Analysis Assay
4.5. Compound Cytotoxicity Assay
4.5.1. Cell Viability Assay
4.5.2. Lactate Dehydrogenase Leakage Assay
4.5.3. Cardio-Toxicity Assay
4.6. Cell Proliferation Assay
4.6.1. Cell Viability Assay
4.6.2. Real-Time Cell Analysis Assay
4.7. Data Analysis and Statistics
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Grant, K.; Loizidou, M.; Taylor, I. Endothelin-1: A multifunctional molecule in cancer. Br. J. Cancer 2003, 88, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Rosano, L.; Spinella, F.; Bagnato, A. Endothelin 1 in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer 2013, 13, 637–651. [Google Scholar] [CrossRef] [PubMed]
- Shichiri, M.; Hirata, Y.; Nakajima, T.; Ando, K.; Imai, T.; Yanagisawa, M.; Masaki, T.; Marumo, F. Endothelin-1 is an autocrine/paracrine growth factor for human cancer cell lines. J. Clin. Investig. 1991, 87, 1867–1871. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.; Bagnato, A.; Battistini, B.; Nisen, P. The endothelin axis: emerging role in cancer. Nat. Rev. Cancer 2003, 3, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.K.; Dayyani, F.; Gallick, G.E. Steps in prostate cancer progression that lead to bone metastasis. Int. J. Cancer 2011, 128, 2545–2561. [Google Scholar] [CrossRef] [PubMed]
- Nie, S.; Zhou, J.; Bai, F.; Jiang, B.; Chen, J.; Zhou, J. Role of endothelin A receptor in colon cancer metastasis: In vitro and in vivo evidence. Mol. Carcinog. 2014, 53, E85–E91. [Google Scholar] [CrossRef] [PubMed]
- Bagnato, A.; Cirilli, A.; Salani, D.; Simeone, P.; Muller, A.; Nicotra, M.R.; Natali, P.G.; Venuti, A. Growth inhibition of cervix carcinoma cells in vivo by endothelin A receptor blockade. Cancer Res. 2002, 62, 6381–6384. [Google Scholar] [PubMed]
- Banerjee, S.; Hussain, M.; Wang, Z.; Saliganan, A.; Che, M.; Bonfil, D.; Cher, M.; Sarkar, F.H. In vitro and in vivo molecular evidence for better therapeutic efficacy of ABT-627 and taxotere combination in prostate cancer. Cancer Res. 2007, 67, 3818–3826. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.U.; Dashwood, M.R.; Heetun, M.; Shiwen, X.; Farooqui, N.; Ramesh, B.; Welch, H.; Savage, F.J.; Ogunbiyi, O.; Abraham, D.J.; et al. Efficacy of the specific endothelin a receptor antagonist zibotentan (ZD4054) in colorectal cancer: A preclinical study. Mol. Cancer Ther. 2013, 12, 1556–1567. [Google Scholar] [CrossRef] [PubMed]
- Daub, H.; Weiss, F.U.; Wallasch, C.; Ullrich, A. Role of transactivation of the EGF receptor in signalling by G-protein-coupled receptors. Nature 1996, 379, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Grant, K.; Knowles, J.; Dawas, K.; Burnstock, G.; Taylor, I.; Loizidou, M. Mechanisms of endothelin 1-stimulated proliferation in colorectal cancer cell lines. Br. J. Surg. 2007, 94, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Akhavan, A.; McHugh, K.H.; Guruli, G.; Bies, R.R.; Zamboni, W.C.; Strychor, S.A.; Nelson, J.B.; Pflug, B.R. Endothelin receptor A blockade enhances taxane effects in prostate cancer. Neoplasia 2006, 8, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Rosano, L.; Di Castro, V.; Spinella, F.; Tortora, G.; Nicotra, M.R.; Natali, P.G.; Bagnato, A. Combined targeting of endothelin A receptor and epidermal growth factor receptor in ovarian cancer shows enhanced antitumor activity. Cancer Res. 2007, 67, 6351–6359. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Liu, X.; Li, W. Tetrandrine, a Chinese plant-derived alkaloid, is a potential candidate for cancer chemotherapy. Oncotarget 2016. [Google Scholar] [CrossRef] [PubMed]
- Apaya, M.K.; Chang, M.T.; Shyur, L.F. Phytomedicine polypharmacology: Cancer therapy through modulating the tumor microenvironment and oxylipin dynamics. Pharmacol. Ther. 2016, 162, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Moselhy, J.; Srinivasan, S.; Ankem, M.K.; Damodaran, C. Natural products that target cancer stem cells. Anticancer Res. 2015, 35, 5773–5788. [Google Scholar] [PubMed]
- Fan, H.Y.; Fu, F.H.; Yang, M.Y.; Xu, H.; Zhang, A.H.; Liu, K. Antiplatelet and antithrombotic activities of salvianolic acid A. Thromb. Res. 2010, 126, e17–e22. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.T.; Zhang, T.M.; Wang, B.E.; Wang, Y.W. Protective action of seven natural phenolic compounds against peroxidative damage to biomembranes. Biochem. Pharmacol. 1992, 43, 147–152. [Google Scholar] [CrossRef]
- Oh, K.S.; Oh, B.K.; Mun, J.; Seo, H.W.; Lee, B.H. Salvianolic acid A suppress lipopolysaccharide-induced NF-κB signaling pathway by targeting IKKβ. Int. Immunopharmacol. 2011, 11, 1901–1906. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Chen, S.; Yang, Q.; Cai, J.; Zhang, W.; You, H.; Xing, J.; Dong, Y. Salvianolic acid A reverses the paclitaxel resistance and inhibits the migration and invasion abilities of human breast cancer cells by inactivating transgelin 2. Cancer Biol. Ther. 2015, 16, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Chen, S.; Zhang, W.; Zheng, X.; Hu, S.; Pang, C.; Lu, J.; Xing, J.; Dong, Y. Salvianolic acid A reverses paclitaxel resistance in human breast cancer MCF-7 cells via targeting the expression of transgelin 2 and attenuating PI3 K/Akt pathway. Phytomed. Int. J. Phytother. Phytopharm. 2014, 21, 1725–1732. [Google Scholar] [CrossRef] [PubMed]
- Ihara, M.; Ishikawa, K.; Fukuroda, T.; Saeki, T.; Funabashi, K.; Fukami, T.; Suda, H.; Yano, M. In vitro biological profile of a highly potent novel endothelin (ET) antagonist BQ-123 selective for the ETA receptor. J. Cardiovasc. Pharmacol. 1992, 20, S11–S14. [Google Scholar] [CrossRef] [PubMed]
- Drimal, J.; Drimal, J., Jr.; Drimal, D. Enhanced endothelin ET(B) receptor down-regulation in human tumor cells. Eur. J. Pharmacol. 2000, 396, 19–22. [Google Scholar] [CrossRef]
- Ke, N.; Nguyen, K.; Irelan, J.; Abassi, Y.A. Multidimensional GPCR profiling and screening using impedance-based label-free and real-time assay. Methods Mol. Biol. 2015, 1272, 215–226. [Google Scholar] [PubMed]
- Yu, Y.; Sun, S.; Wang, S.; Zhang, Q.; Li, M.; Lan, F.; Li, S.; Liu, C. Liensinine- and neferine-induced cardiotoxicity in primary neonatal rat cardiomyocytes and human-induced pluripotent stem cell-derived cardiomyocytes. Int. J. Mol. Sci. 2016, 17, 186. [Google Scholar] [CrossRef] [PubMed]
- Clements, M.; Millar, V.; Williams, A.S.; Kalinka, S. Bridging functional and structural cardiotoxicity assays using human embryonic stem cell-derived cardiomyocytes for a more comprehensive risk assessment. Toxicol. Sci. 2015, 148, 241–260. [Google Scholar] [CrossRef] [PubMed]
- Nguemo, F.; Saric, T.; Pfannkuche, K.; Watzele, M.; Reppel, M.; Hescheler, J. In vitro model for assessing arrhythmogenic properties of drugs based on high-resolution impedance measurements. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2012, 29, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Xu, J.; Li, D.; Chen, J.; Shen, X.; Xu, F.; Teng, F.; Deng, Y.; Ma, H.; Zhang, L.; et al. Salvianolic acid A, a matrix metalloproteinase-9 inhibitor of Salvia miltiorrhiza, attenuates aortic aneurysm formation in apolipoprotein E-deficient mice. Phytomed. Int. J. Phytother. Phytopharm. 2014, 21, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Chouaid, C.; Nathan, F.; Pemberton, K.; Morris, T. A phase II, randomized, multicenter study to assess the efficacy, safety, and tolerability of zibotentan (ZD4054) in combination with pemetrexed in patients with advanced non-small cell lung cancer. Cancer Chemother. Pharmacol. 2011, 67, 1203–1208. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.B.; Love, W.; Chin, J.L.; Saad, F.; Schulman, C.C.; Sleep, D.J.; Qian, J.; Steinberg, J.; Carducci, M. Atrasentan Phase 3 Study Guides Phase 3, randomized, controlled trial of atrasentan in patients with nonmetastatic, hormone-refractory prostate cancer. Cancer 2008, 113, 2478–2487. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, V.; Breznan, D.; Thomson, E.; Karthikeyan, S.; Vincent, R. Effects of ambient air particles on the endothelin system in human pulmonary epithelial cells (A549). Cell Biol. Toxicol. 2005, 21, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Xiong, H.; Kim, T.H.; Ren, B.; Zhang, Z. Positive inter-regulation between β-catenin/T cell factor-4 signaling and endothelin-1 signaling potentiates proliferation and survival of prostate cancer cells. Mol. Pharmacol. 2006, 69, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Von Brandenstein, M.G.; Ngum Abety, A.; Depping, R.; Roth, T.; Koehler, M.; Dienes, H.P.; Fries, J.W. A p38-p65 transcription complex induced by endothelin-1 mediates signal transduction in cancer cells. Biochim. Biophys. Acta 2008, 1783, 1613–1622. [Google Scholar] [CrossRef] [PubMed]
- Tocci, P.; Caprara, V.; Cianfrocca, R.; Sestito, R.; Di Castro, V.; Bagnato, A.; Rosano, L. Endothelin-1/endothelin A receptor axis activates RhoA GTPase in epithelial ovarian cancer. Life Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Bi, L.; Chen, J.; Yuan, X.; Jiang, Z.; Chen, W. Salvianolic acid A positively regulates PTEN protein level and inhibits growth of A549 lung cancer cells. Biomed. Rep. 2013, 1, 213–217. [Google Scholar] [PubMed]
- Masarik, M.; Gumulec, J.; Hlavna, M.; Sztalmachova, M.; Babula, P.; Raudenska, M.; Pavkova-Goldbergova, M.; Cernei, N.; Sochor, J.; Zitka, O.; et al. Monitoring of the prostate tumour cells redox state and real-time proliferation by novel biophysical techniques and fluorescent staining. Integr. Biol. Quant. Biosci. Nano Macro 2012, 4, 672–684. [Google Scholar] [CrossRef] [PubMed]
- Kuzumaki, N.; Suzuki, A.; Narita, M.; Hosoya, T.; Nagasawa, A.; Imai, S.; Yamamizu, K.; Morita, H.; Suzuki, T.; Okada, Y.; et al. Multiple analyses of G-protein coupled receptor (GPCR) expression in the development of gefitinib-resistance in transforming non-small-cell lung cancer. PLoS ONE 2012, 7, e44368. [Google Scholar] [CrossRef] [PubMed]
- Rosano, L.; Cianfrocca, R.; Spinella, F.; di Castro, V.; Natali, P.G.; Bagnato, A. Combination therapy of zibotentan with cisplatinum and paclitaxel is an effective regimen for epithelial ovarian cancer. Can. J. Physiol. Pharmacol. 2010, 88, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Bloom, M.W.; Hamo, C.E.; Cardinale, D.; Ky, B.; Nohria, A.; Baer, L.; Skopicki, H.; Lenihan, D.J.; Gheorghiade, M.; Lyon, A.R.; et al. Cancer therapy-related cardiac dysfunction and heart failure: Part 1: Definitions, pathophysiology, risk factors, and imaging. Circ. Heart Fail. 2016, 9, e002661. [Google Scholar] [CrossRef] [PubMed]
- Hamo, C.E.; Bloom, M.W.; Cardinale, D.; Ky, B.; Nohria, A.; Baer, L.; Skopicki, H.; Lenihan, D.J.; Gheorghiade, M.; Lyon, A.R.; et al. Cancer therapy-related cardiac dysfunction and heart failure: Part 2: Prevention, treatment, guidelines, and future directions. Circul. Heart Fail. 2016, 9, e002843. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Yang, L.; Fu, F.; Xu, H.; Meng, Q.; Zhu, H.; Teng, L.; Yang, M.; Zhang, L.; Zhang, Z.; et al. Cardioprotective effects of salvianolic Acid a on myocardial ischemia-reperfusion injury in vivo and in vitro. Evid. Based Complement. Altern. Med. eCAM 2012, 2012, 508938. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.H.; Chang, H.J.; Cho, J.Y.; Chun, H.S. Cytoprotective effect of anthocyanins against doxorubicin-induced toxicity in H9c2 cardiomyocytes in relation to their antioxidant activities. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2007, 45, 1873–1881. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, S.; Guo, L.; Mussio, J.; Furniss, M.; Hamre, J., 3rd; Davis, M. Examining the protective role of ErbB2 modulation in human-induced pluripotent stem cell-derived cardiomyocytes. Toxicol. Sci. 2014, 141, 547–559. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Eldridge, S.; Furniss, M.; Mussio, J.; Davis, M. Use of human induced pluripotent stem cell-derived cardiomyocytes (hiPSC-CMs) to monitor compound effects on cardiac myocyte signaling pathways. Curr. Protoc. Chem. Biol. 2015, 7, 141–185. [Google Scholar] [PubMed]
- Guo, L.; Coyle, L.; Abrams, R.M.; Kemper, R.; Chiao, E.T.; Kolaja, K.L. Refining the human iPSC-cardiomyocyte arrhythmic risk assessment model. Toxicol. Sci. 2013, 136, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.; Lan, F.; Lee, A.S.; Gong, T.; Sanchez-Freire, V.; Wang, Y.; Diecke, S.; Sallam, K.; Knowles, J.W.; Wang, P.J.; et al. Drug screening using a library of human induced pluripotent stem cell-derived cardiomyocytes reveals disease-specific patterns of cardiotoxicity. Circulation 2013, 127, 1677–1691. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Inhibition (%) | IC50 Values (µM) | |||
|---|---|---|---|---|---|
| 100 µM | 25 µM | 6.25 µM | 1.56 µM | ||
| SKOV3 | 99.13(0.19) | 30.87(3.18) | 1.215(1.48) | 6.001(0.21) | 30.84 |
| HeLa | 96.30(0.49) | 75.65(3.81) | 8.751(1.25) | 3.419(0.15) | 15.85 |
| H1975 | 96.38(0.19) | 73.65(1.84) | 35.09(5.75) | 11.64(1.48) | 10.19 |
| DU145 | 96.48(0.04) | 77.25(2.18) | 32.94(2.26) | 18.15(4.08) | 9.512 |
| A549 | 99.13(0.11) | 58.43(2.39) | 43.40(1.55) | 38.35(2.56) | 6.461 |
| Cell Lines | Sal A | Sal A | Bosentan | 0.1% DMSO |
|---|---|---|---|---|
| 5 µM | 1.2 µM | 10 µM | ||
| HeLa | 33.5 ± 1.04 ** | 23.7 ± 0.48 ** | 26.3 ± 0.62 ** | 21.1 ± 0.34 |
| DU145 | 41.0 ± 1.35 ** | 35.8 ± 0.37 ** | 29.6 ± 0.26 ** | 25.6 ± 0.19 |
| H1975 | 94.1 ± 2.43 ** | 49.6 ± 1.61 ** | 29.4 ± 0.38 * | 27.1 ± 0.63 |
| A549 | 28.7 ± 1.10 | 28.9 ± 1.08 | 40.9 ± 2.33 ** | 29.7 ± 0.99 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Wang, S.; Yu, Y.; Sun, S.; Zhang, Y.; Zhang, Y.; Yang, W.; Li, S.; Qiao, Y. Salvianolic Acid A, as a Novel ETA Receptor Antagonist, Shows Inhibitory Effects on Tumor in Vitro. Int. J. Mol. Sci. 2016, 17, 1244. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17081244
Zhang Q, Wang S, Yu Y, Sun S, Zhang Y, Zhang Y, Yang W, Li S, Qiao Y. Salvianolic Acid A, as a Novel ETA Receptor Antagonist, Shows Inhibitory Effects on Tumor in Vitro. International Journal of Molecular Sciences. 2016; 17(8):1244. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17081244
Chicago/Turabian StyleZhang, Qiao, Shifeng Wang, Yangyang Yu, Shengnan Sun, Yuxin Zhang, Yanling Zhang, Wei Yang, Shiyou Li, and Yanjiang Qiao. 2016. "Salvianolic Acid A, as a Novel ETA Receptor Antagonist, Shows Inhibitory Effects on Tumor in Vitro" International Journal of Molecular Sciences 17, no. 8: 1244. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17081244