
The Effects of Female Sexual Hormones on the Expression of Aquaporin 5 in the Late-Pregnant Rat Uterus


Abstract
:1. Introduction
2. Results
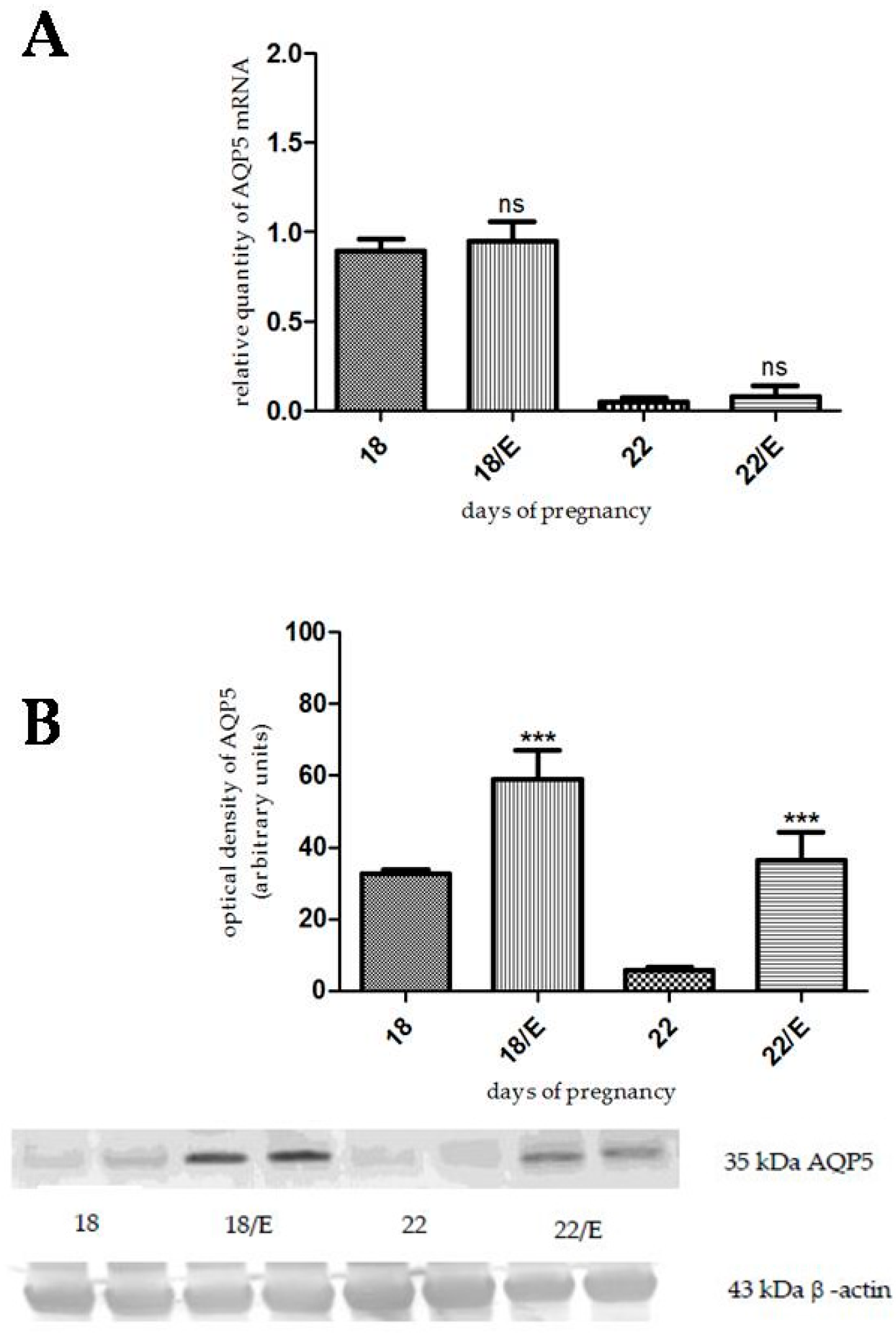
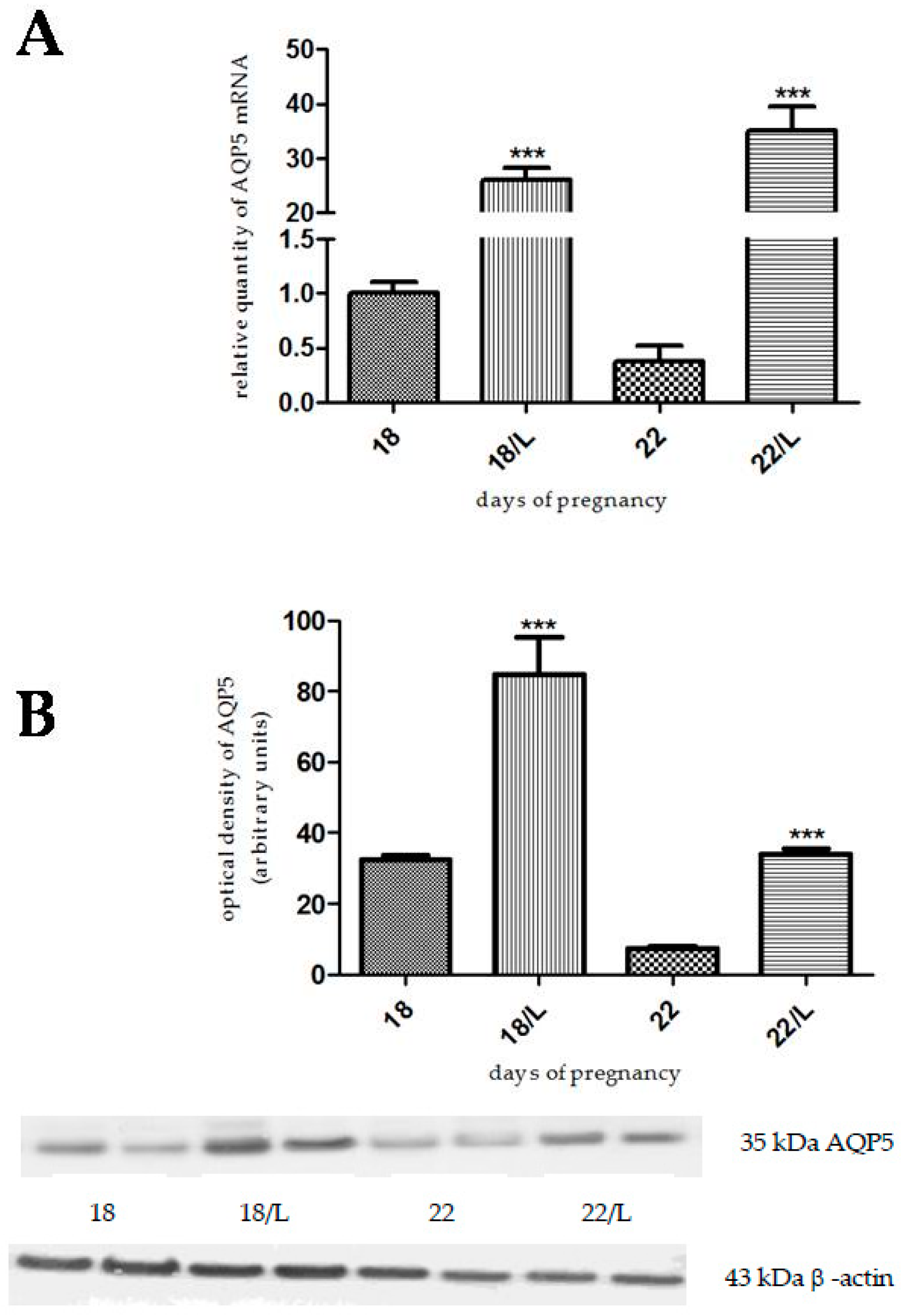
2.1. The Effects of Estrogen-Related Compounds on Aquaporin 5 (AQP5) Expression
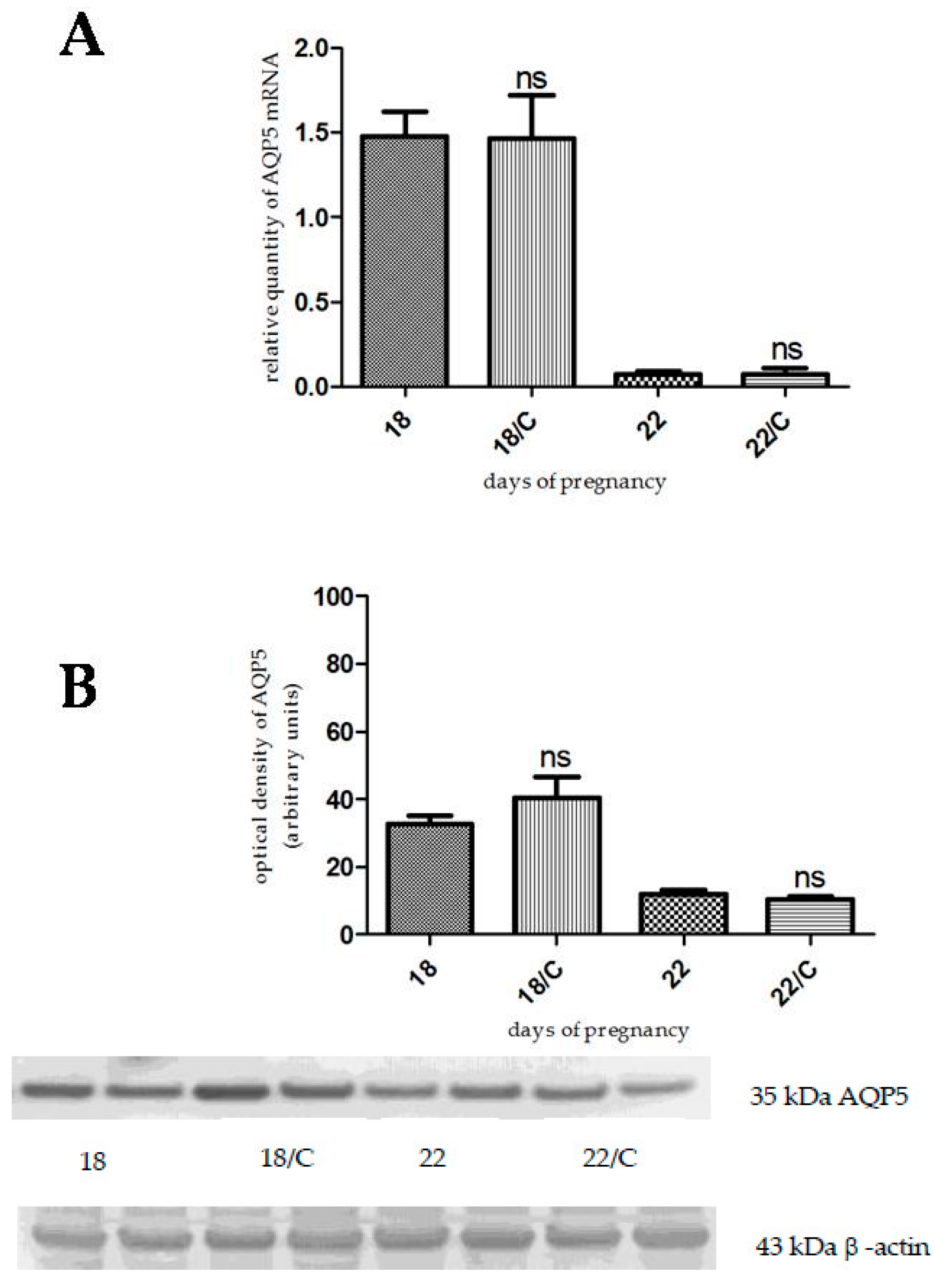
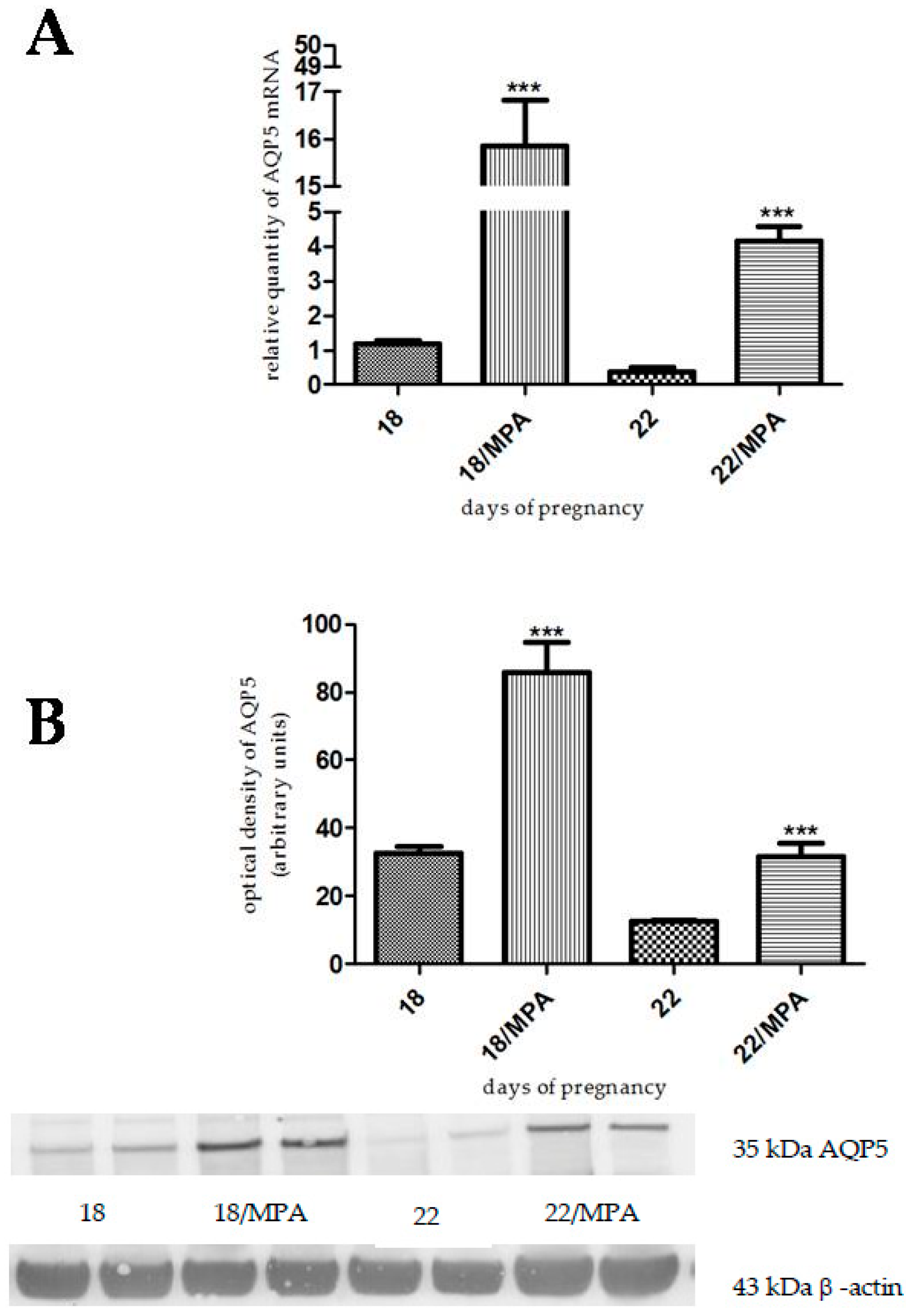
2.2. The Effects of Gestagen-Related Compounds on AQP5 Expression
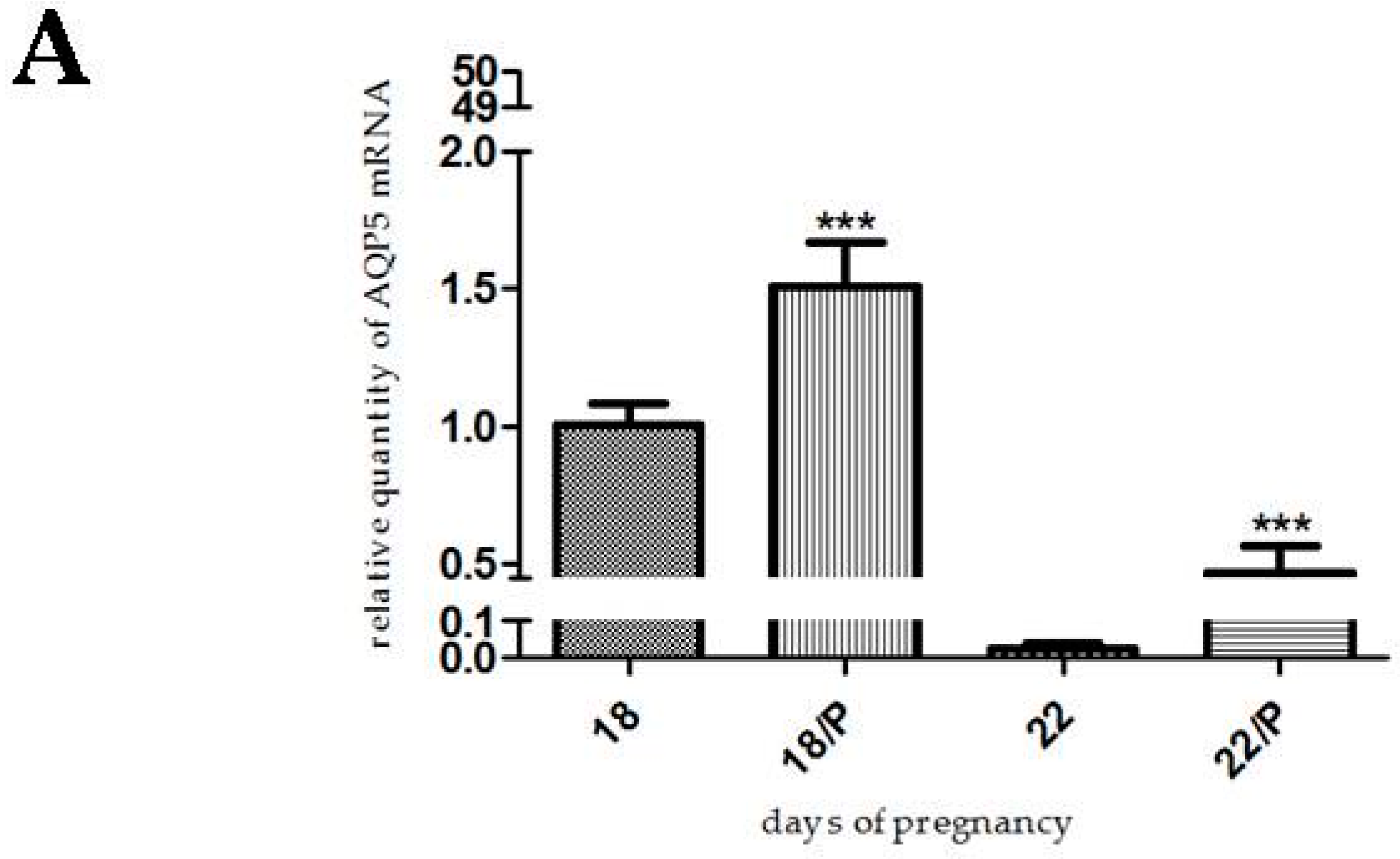
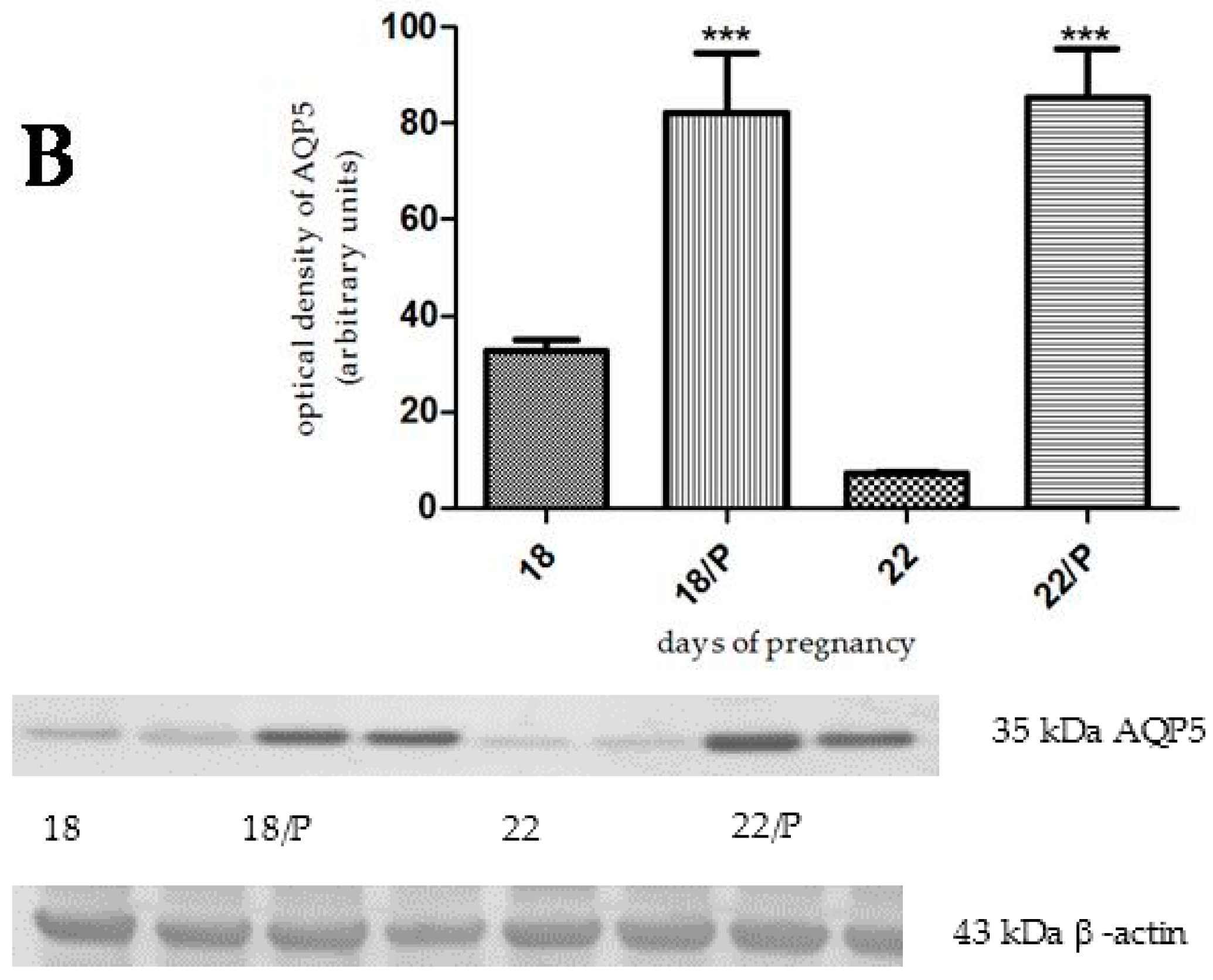
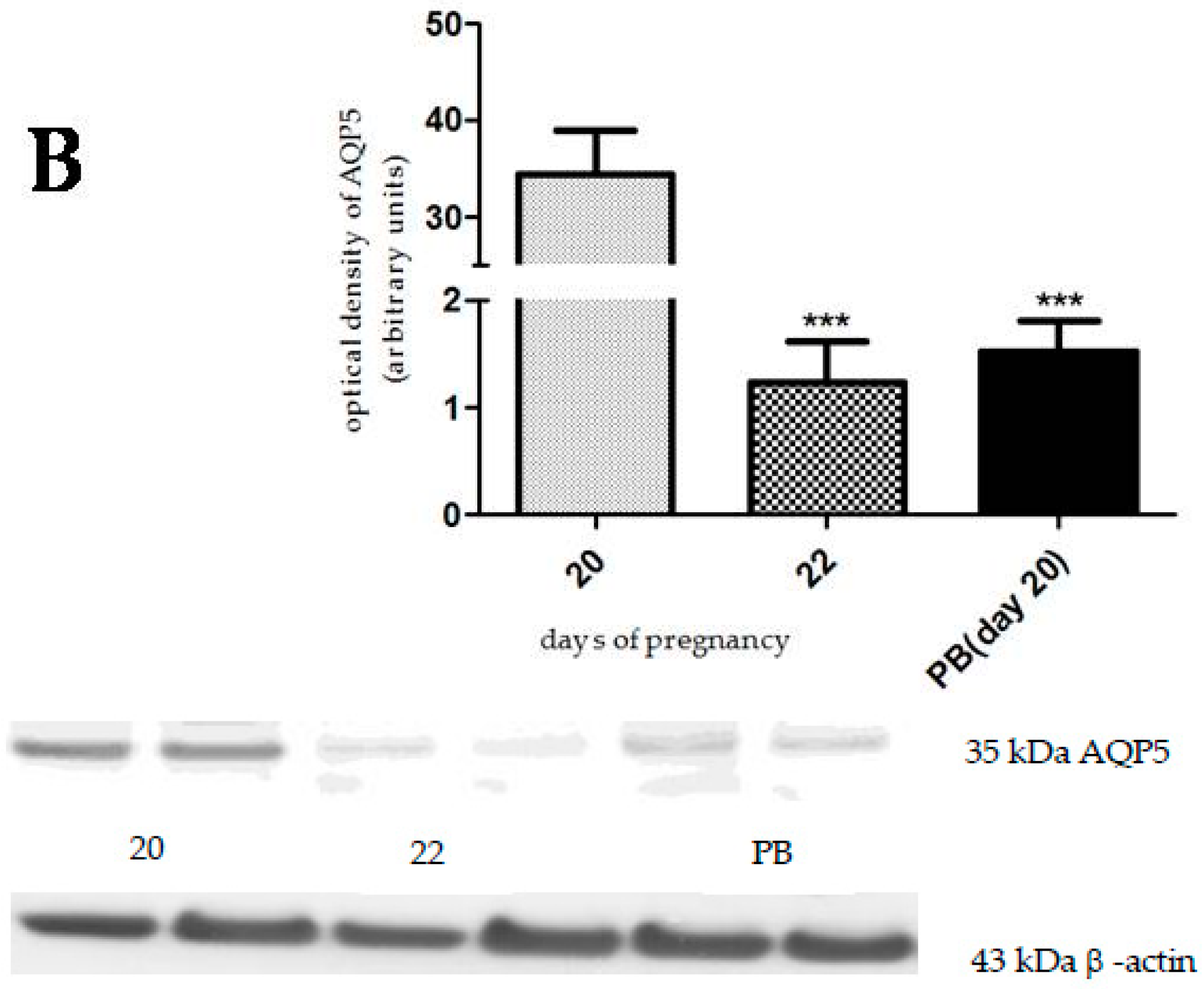
2.3. The Effect of Hormonally-Induced Preterm Delivery on AQP5 Expression
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.1.1. Housing and Handling of the Animals
4.1.2. Mating of the Animals
4.2. In Vivo Treatments of the Rats
4.3. RT-PCR Studies
4.3.1. Tissue Isolation
4.3.2. Total RNA Preparation
4.3.3. Real-Time Quantitative Reverse-Transcriptase PCR
4.4. Western Blot Analysis
4.5. Statistical Analysis
Author Contributions
Conflicts of Interest
Abbreviations
| AQP | Aquaporin |
References
- Denker, B.M.; Smith, B.L.; Kuhajda, F.P.; Agre, P. Identification, purification, and partial characterization of a novel Mr 28,000 integral membrane protein from erythrocytes and renal tubules. J. Biol. Chem. 1988, 263, 15634–15642. [Google Scholar] [PubMed]
- Zhang, D.; Tan, Y.-J.; Qu, F.; Sheng, J.-Z.; Huang, H.-F. Functions of water channels in male and female reproductive systems. Mol. Aspects Med. 2012, 33, 676–690. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Jiang, Z.; Bazer, F.W.; Johnson, G.A.; Burghardt, R.C.; Wu, G. Aquaporins in the female reproductive system of mammals. Front. Biosci. Landmark Ed. 2015, 20, 838–871. [Google Scholar] [PubMed]
- Hara-Chikuma, M.; Verkman, A.S. Physiological roles of glycerol-transporting aquaporins: The aquaglyceroporins. Cell. Mol. Life Sci. 2006, 63, 1386–1392. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Kondo, S.; Hara, S.; Morishita, Y. The evolutionary aspects of aquaporin family. AJP Regul. Integr. Comp. Physiol. 2011, 300, R566–R576. [Google Scholar] [CrossRef] [PubMed]
- Yakata, K.; Hiroaki, Y.; Ishibashi, K.; Sohara, E.; Sasaki, S.; Mitsuoka, K.; Fujiyoshi, Y. Aquaporin-11 containing a divergent NPA motif has normal water channel activity. Biochim. Biophys. Acta BBA Biomembr. 2007, 1768, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Rai, T.; Kuwahara, M.; Ko, S.B.H.; Uchida, S.; Sasaki, S.; Ishibashi, K. Identification of a novel aquaporin, AQP12, expressed in pancreatic acinar cells. Biochem. Biophys. Res. Commun. 2005, 330, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, E.M.; McConnell, N.A.; Hughes, F.M.; Huet-Hudson, Y.M. Estrogen regulation of aquaporins in the mouse uterus: Potential roles in uterine water movement. Biol. Reprod. 2003, 69, 1481–1487. [Google Scholar] [CrossRef] [PubMed]
- Sha, X.Y.; Xiong, Z.F.; Liu, H.S.; Zheng, Z.; Ma, T.H. Pregnant phenotype in aquaporin 8-deficient mice. Acta Pharmacol. Sin. 2011, 32, 840–844. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Liu, H.; Beall, M.; Ma, T.; Hao, R.; Ross, M.G. Role of aquaporin 1 in fetal fluid homeostasis. J. Matern. Fetal Neonatal Med. 2014, 27, 505–510. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Q.; Zhang, H.; Wang, H.; Ma, T.; Qiao, J.; Duan, E. Aquaporin-dependent excessive intrauterine fluid accumulation is a major contributor in hyper-estrogen induced aberrant embryo implantation. Cell Res. 2015, 25, 139–142. [Google Scholar]
- He, R.-H.; Sheng, J.-Z.; Luo, Q.; Jin, F.; Wang, B.; Qian, Y.-L.; Zhou, C.-Y.; Sheng, X.; Huang, H.-F. Aquaporin-2 expression in human endometrium correlates with serum ovarian steroid hormones. Life Sci. 2006, 79, 423–429. [Google Scholar]
- Zou, L.-B.; Zhang, R.-J.; Tan, Y.-J.; Ding, G.-L.; Shi, S.; Zhang, D.; He, R.-H.; Liu, A.-X.; Wang, T.-T.; Leung, P.C.K.; et al. Identification of estrogen response element in the aquaporin-2 gene that mediates estrogen-induced cell migration and invasion in human endometrial carcinoma. J. Clin. Endocrinol. Metab. 2011, 96, E1399–E1408. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Takahashi, E.; Miyagawa, S.; Watanabe, H.; Iguchi, T. Chromatin immunoprecipitation-mediated target identification proved aquaporin 5 is regulated directly by estrogen in the uterus. Genes Cells 2006, 11, 1133–1143. [Google Scholar] [CrossRef] [PubMed]
- Aralla, M.; Borromeo, V.; Groppetti, D.; Secchi, C.; Cremonesi, F.; Arrighi, S. A collaboration of aquaporins handles water transport in relation to the estrous cycle in the bitch uterus. Theriogenology 2009, 72, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Skowronska, A.; Młotkowska, P.; Wojciechowicz, B.; Okrasa, S.; Nielsen, S.; Skowronski, M.T. Progesterone, estradiol, arachidonic acid, oxytocin, forskolin and cAMP influence on aquaporin 1 and 5 expression in porcine uterine explants during the mid-luteal phase of the estrous cycle and luteolysis: An in vitro study. Reprod. Biol. Endocrinol. 2015, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Salleh, N.; Mokhtar, H.M.; Kassim, N.M.; Giribabu, N. Testosterone induces increase in aquaporin (AQP)-1, 5, and 7 expressions in the uteri of ovariectomized rats. J. Membr. Biol. 2015, 248, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, L.A.; Murphy, C.R. Redistribution of aquaporins 1 and 5 in the rat uterus is dependent on progesterone: A study with light and electron microscopy. Reproduction 2006, 131, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Richard, C.; Gao, J.; Brown, N.; Reese, J. Aquaporin water channel genes are differentially expressed and regulated by ovarian steroids during the periimplantation period in the mouse. Endocrinology 2003, 144, 1533–1541. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, L.A.; Murphy, C.R. Redistribution of aquaporins in uterine epithelial cells at the time of implantation in the rat. Acta Histochem. 2004, 106, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Yasui, M. Cellular and subcellular localization of aquaporins 1, 3, 8, and 9 in amniotic membranes during pregnancy in mice. Cell Tissue Res. 2010, 342, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Ducza, E.; Seres, A.B.; Hajagos-Tóth, J.; Falkay, G.; Gáspár, R. Oxytocin regulates the expression of aquaporin 5 in the late-pregnant rat uterus: Alterations in AQPs in the late-pregnant rat uterus. Mol. Reprod. Dev. 2014, 81, 524–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, L.A.; Murphy, C.R. Aquaporins are upregulated in glandular epithelium at the time of implantation in the rat. J. Mol. Histol. 2007, 38, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Boroditsky, R.S.; Reyes, F.I.; Winter, J.S.; Faiman, C. Maternal serum estrogen and progesterone concentrations preceding normal labor. Obstet. Gynecol. 1978, 51, 686–691. [Google Scholar] [PubMed]
- Tulchinsky, D.; Hobel, C.J.; Yeager, E.; Marshall, J.R. Plasma estrone, estradiol, estriol, progesterone, and 17-hydroxyprogesterone in human pregnancy. I. Normal pregnancy. Am. J. Obstet. Gynecol. 1972, 112, 1095–1100. [Google Scholar] [CrossRef]
- Shaikh, A.A. Estrone and estradiol levels in the ovarian venous blood from rats during the estrous cycle and pregnancy. Biol. Reprod. 1971, 5, 297–307. [Google Scholar] [PubMed]
- Morishige, W.K.; Pepe, G.J.; Rothchild, I. Serum luteinizing hormone, prolactin and progesterone levels during pregnancy in the rat. Endocrinology 1973, 92, 1527–1530. [Google Scholar] [CrossRef] [PubMed]
- Schindler, A.E.; Campagnoli, C.; Druckmann, R.; Huber, J.; Pasqualini, J.R.; Schweppe, K.W.; Thijssen, J.H. Classification and pharmacology of progestins. Maturitas 2003, 46, 7–16. [Google Scholar] [CrossRef]
- Philibert, D.; Bouchoux, F.; Degryse, M.; Lecaque, D.; Petit, F.; Gaillard, M. The pharmacological profile of a novel norpregnane progestin (trimegestone). Gynecol. Endocrinol. 1999, 13, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Maier, T.; Güell, M.; Serrano, L. Correlation of mRNA and protein in complex biological samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, M.; Smith, C.L. Molecular mechanisms of selective estrogen receptor modulator (SERM) action. J. Pharmacol. Exp. Ther. 2000, 295, 431–437. [Google Scholar] [PubMed]
- Dickey, R.P. Clomiphene citrate for women with unexplained infertility. In Unexplained Infertility; Schattman, G.L., Esteves, S.C., Agarwal, A., Eds.; Springer New York: New York, NY, USA, 2015; pp. 261–271. [Google Scholar]
- Telleria, C.M.; Deis, R.P. Reproductive function in rats after mifepristone-induced termination of pregnancy. Contraception 1996, 53, 185–190. [Google Scholar] [CrossRef]
- Thomas, J.; Fairclough, A.; Kavanagh, J.; Kelly, A.J. Vaginal prostaglandin (PGE2 and PGF2a) for induction of labour at term. In Cochrane Database of Systematic Reviews; Cochrane Pregnancy and Childbirth Group, Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2014. [Google Scholar]
- Bóta, J.; Hajagos-Tóth, J.; Ducza, E.; Samavati, R.; Borsodi, A.; Benyhe, S.; Gáspár, R. The effects of female sexual hormones on the expression and function of α1A- and α1D-adrenoceptor subtypes in the late-pregnant rat myometrium. Eur. J. Pharmacol. 2015, 769, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Hódi, Á.; Földesi, I.; Ducza, E.; Hajagos-Tóth, J.; Seres, A.B.; Klukovits, A.; Gáspár, R. Tocopherol inhibits the relaxing effect of terbutaline in the respiratory and reproductive tracts of the rat: The role of the oxidative stress index. Life Sci. 2014, 105, 48–55. [Google Scholar]
- Cyr, M. Modulation by estrogen-receptor directed drugs of 5-hydroxytryptamine-2A receptors in rat brain. Neuropsychopharmacology 2000, 23, 69–78. [Google Scholar] [CrossRef]
- Nutu, M.; Feng, Y.; Egecioglu, E.; Weijdegård, B.; Stener-Victorin, E.; Shao, R. Stromal cell-specific apoptotic and antiestrogenic mechanisms may explain uterine defects in humans after clomiphene citrate therapy. Am. J. Obstet. Gynecol. 2010, 203, 65.e1–65.e10. [Google Scholar] [CrossRef] [PubMed]
- Bharti, S.; Misro, M.M.; Rai, U. Clomiphene citrate potentiates the adverse effects of estrogen on rat testis and down-regulates the expression of steroidogenic enzyme genes. Fertil. Steril. 2013, 99, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Graham, B.M.; Milad, M.R. Blockade of estrogen by hormonal contraceptives impairs fear extinction in female rats and women. Biol. Psychiatry 2013, 73, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Brandon, J.M.; Raval, P.J. Interaction of estrogen and histamine during ovum implantation in the rat. Eur. J. Pharmacol. 1979, 57, 171–177. [Google Scholar] [CrossRef]
- Hajagos-Toth, J.; Bota, J.; Ducza, E.; Csanyi, A.; Tiszai, Z.; Borsodi, A.; Samavati, R.; Benyhe, S.; Gaspar, R. The effects of estrogen on the α2-adrenergic receptor subtypes in rat uterine function in late pregnancy in vitro. Croat. Med. J. 2016, 57, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Hajagos-Tóth, J.; Bóta, J.; Ducza, E.; Samavati, R.; Borsodi, A.; Benyhe, S.; Gáspár, R. The effects of progesterone on the α2-adrenergic receptor subtypes in late-pregnant uterine contractions in vitro. Reprod. Biol. Endocrinol. 2016, 14, 33. [Google Scholar] [CrossRef] [PubMed]
- Elovitz, M.A.; Mrinalini, C. Animal models of preterm birth. Trends Endocrinol. Metab. 2004, 15, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TaqMan Assays | Assay ID (ThermoFisher Scientific) | Accession Number | Assay Location | Amplicon Length | Annealing Temp. (°C) | Reaction Volume (µL) |
|---|---|---|---|---|---|---|
| AQP5 | Rn00562837_m1 | NM_012779.1 | 473 | 69 | 60 | 20 |
| β-Actin | Rn00667869_m1 | NM_031144.3 | 881 | 91 | 60 | 20 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Csányi, A.; Bóta, J.; Falkay, G.; Gáspár, R.; Ducza, E. The Effects of Female Sexual Hormones on the Expression of Aquaporin 5 in the Late-Pregnant Rat Uterus. Int. J. Mol. Sci. 2016, 17, 1300. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17081300
Csányi A, Bóta J, Falkay G, Gáspár R, Ducza E. The Effects of Female Sexual Hormones on the Expression of Aquaporin 5 in the Late-Pregnant Rat Uterus. International Journal of Molecular Sciences. 2016; 17(8):1300. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17081300
Chicago/Turabian StyleCsányi, Adrienn, Judit Bóta, George Falkay, Robert Gáspár, and Eszter Ducza. 2016. "The Effects of Female Sexual Hormones on the Expression of Aquaporin 5 in the Late-Pregnant Rat Uterus" International Journal of Molecular Sciences 17, no. 8: 1300. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17081300