
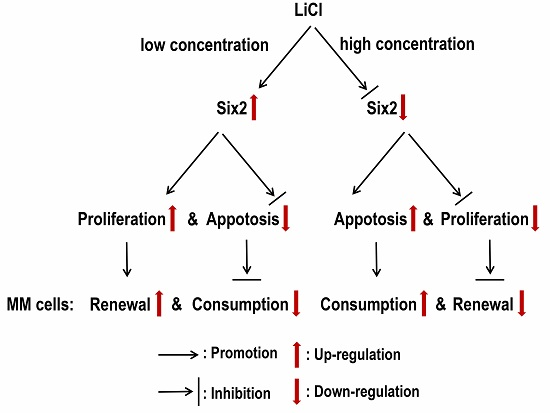
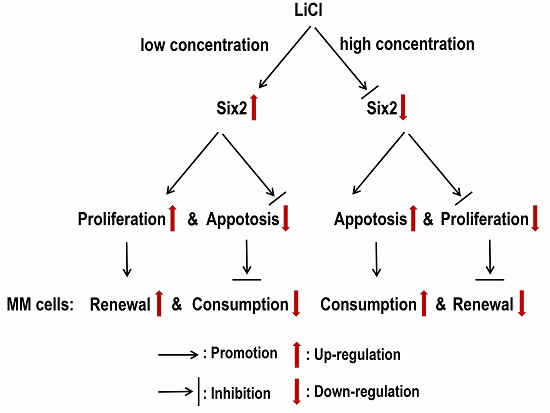
Six2 Is a Coordinator of LiCl-Induced Cell Proliferation and Apoptosis

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
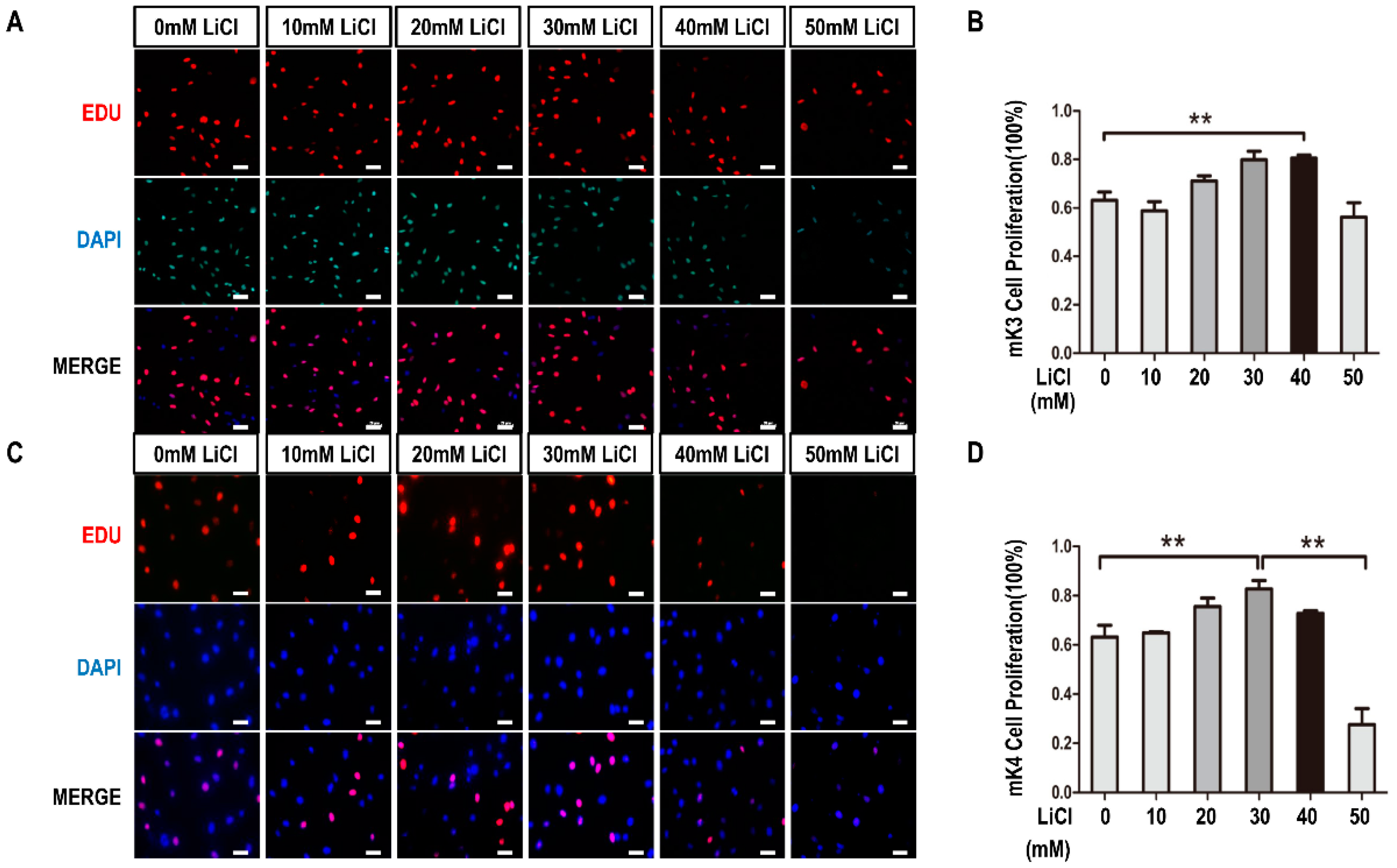
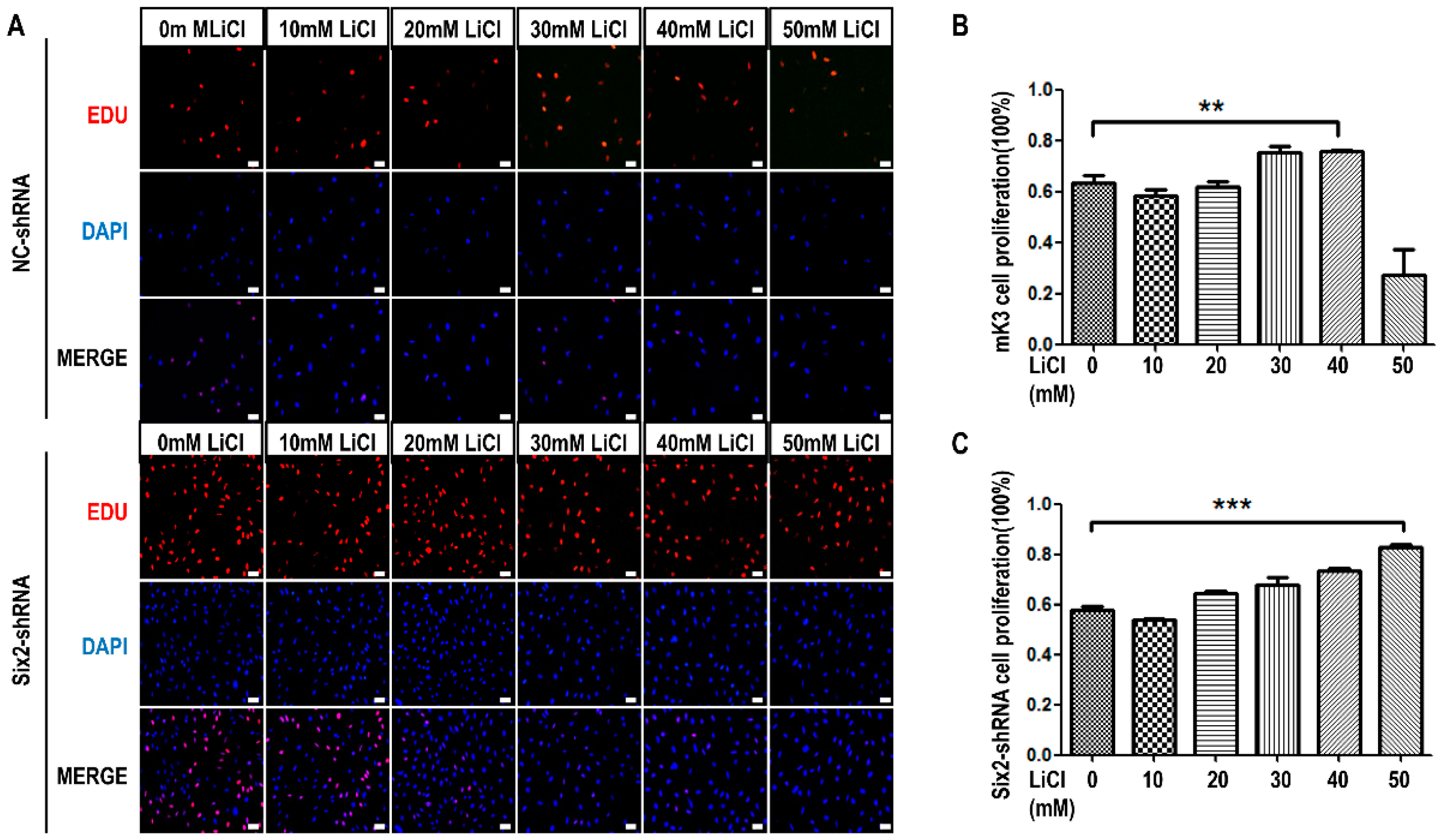
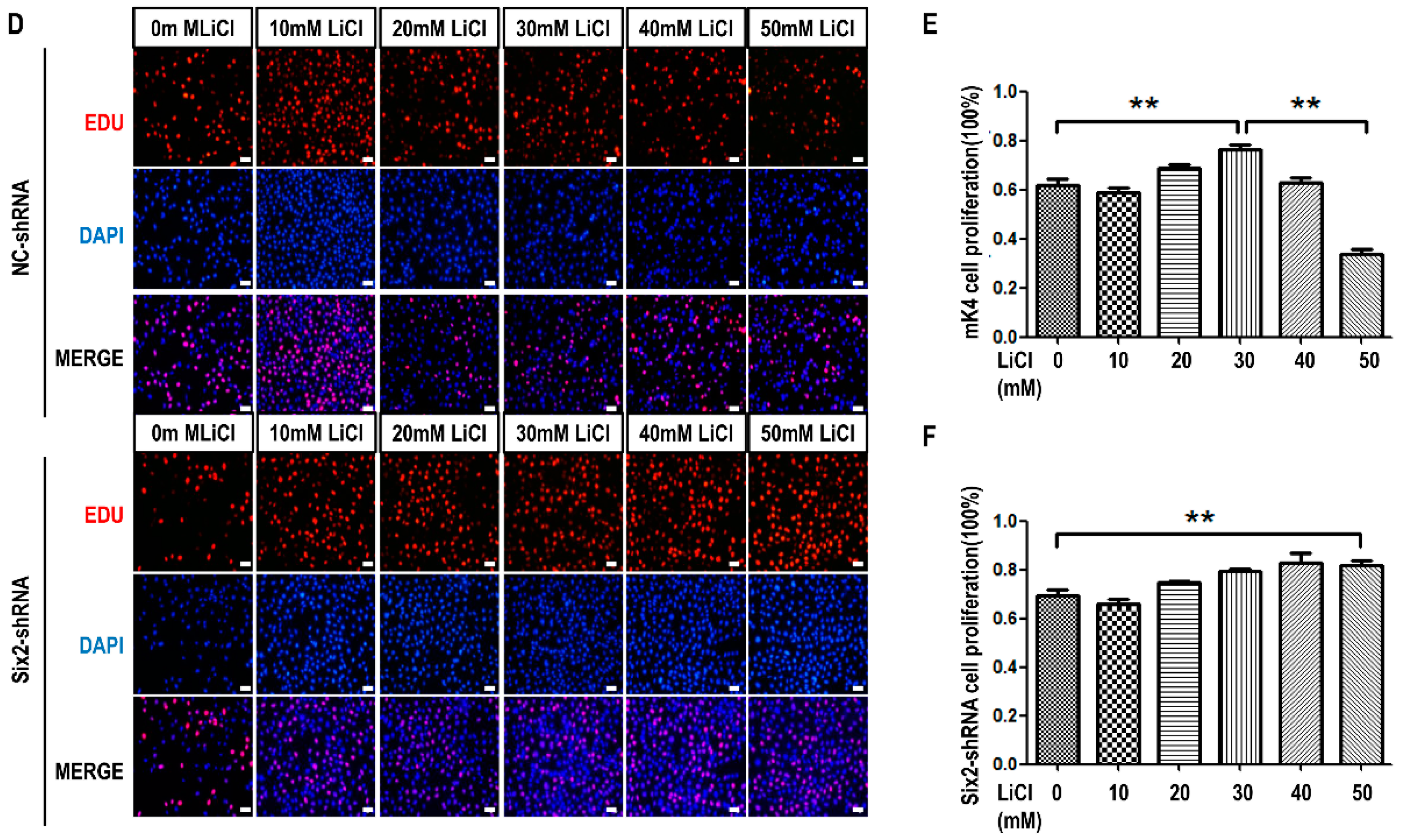
2.1. LiCl Promotes the Proliferation of Metanephric mesenchyme (MM) Cells at Low-Concentration and Inhibits It at High-Concentration
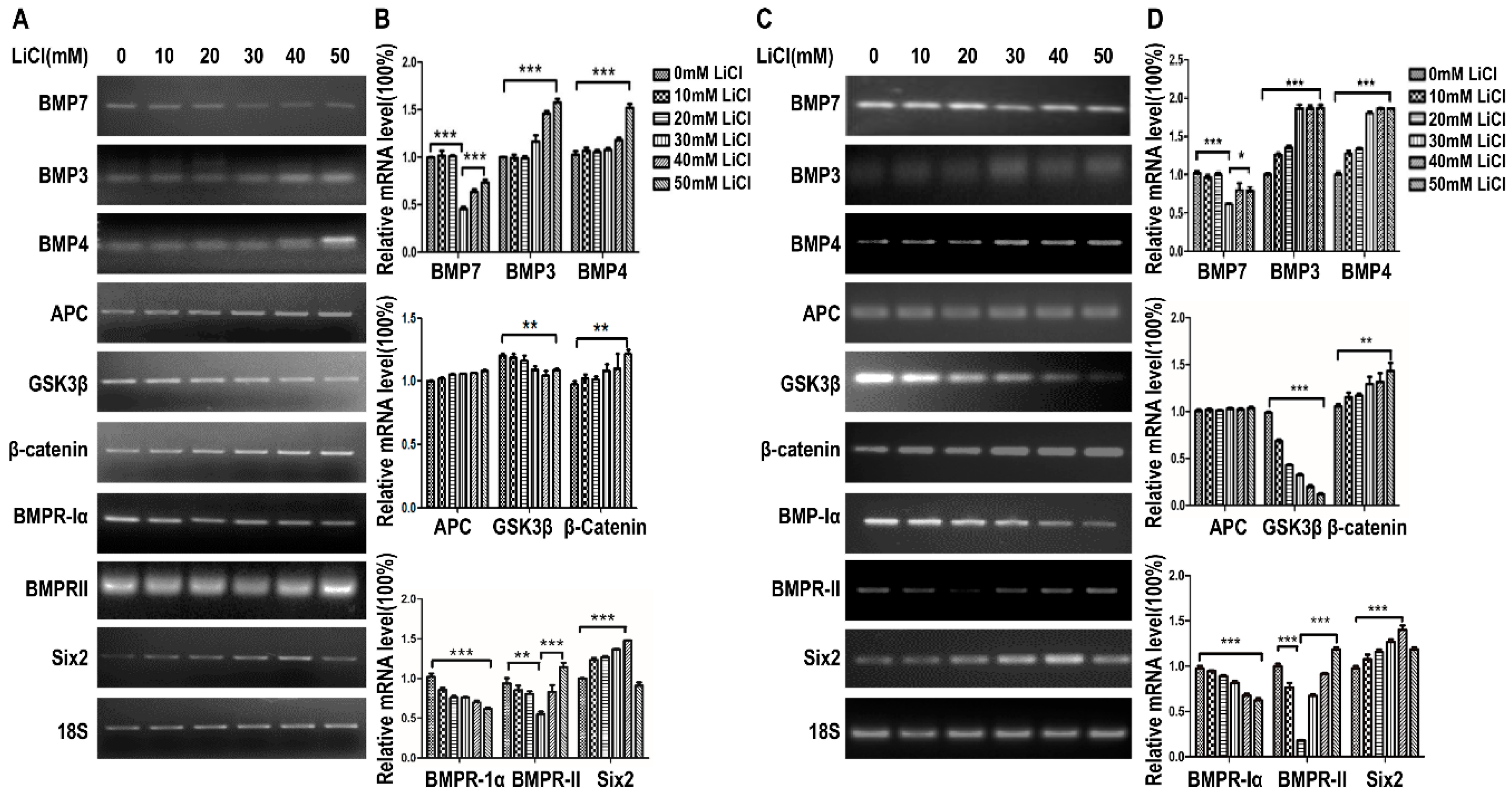
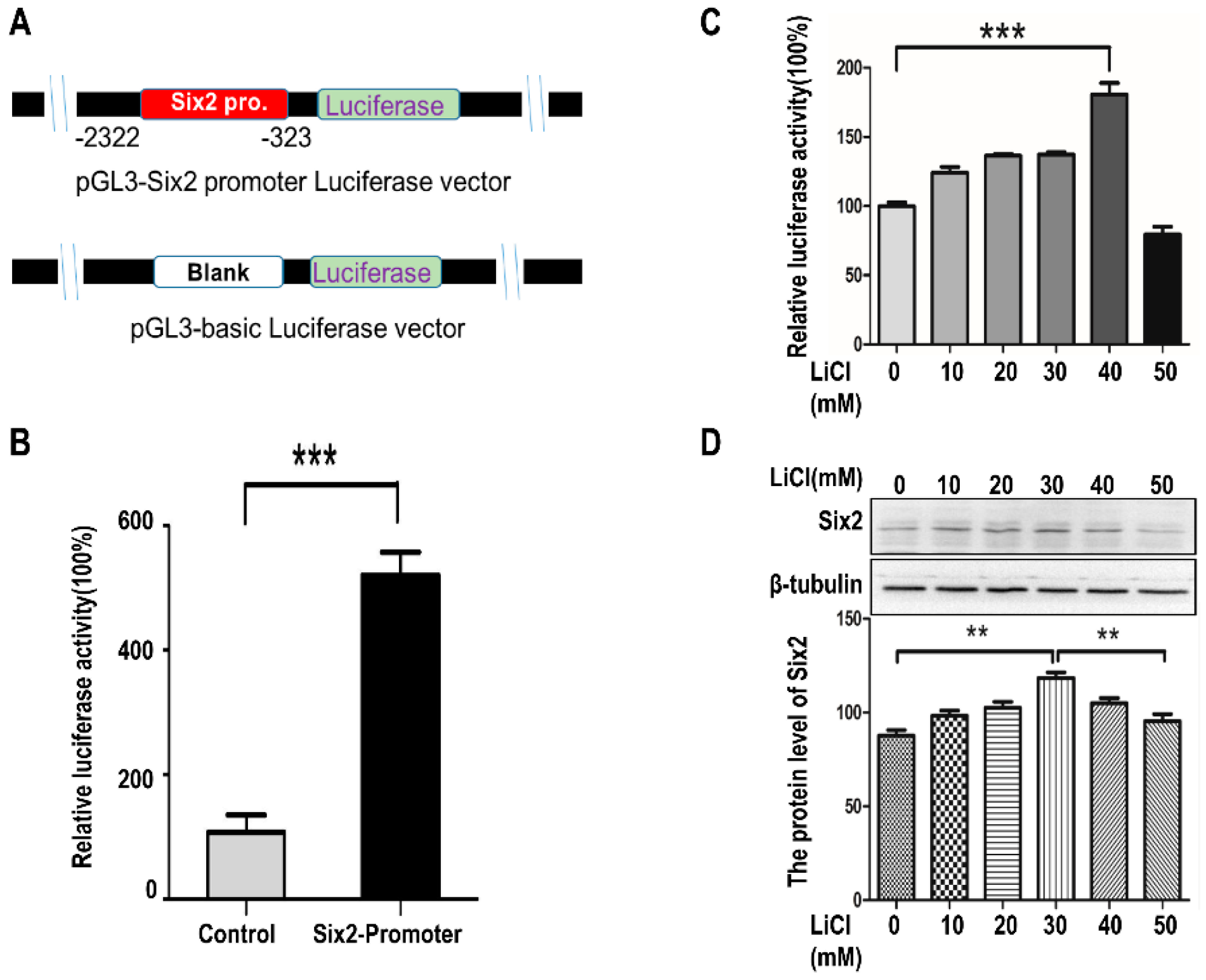
2.2. LiCl Up-Regulates the Expression of Six2 at Low-Concentration and Down-Regulates Six2 at High-Concentration
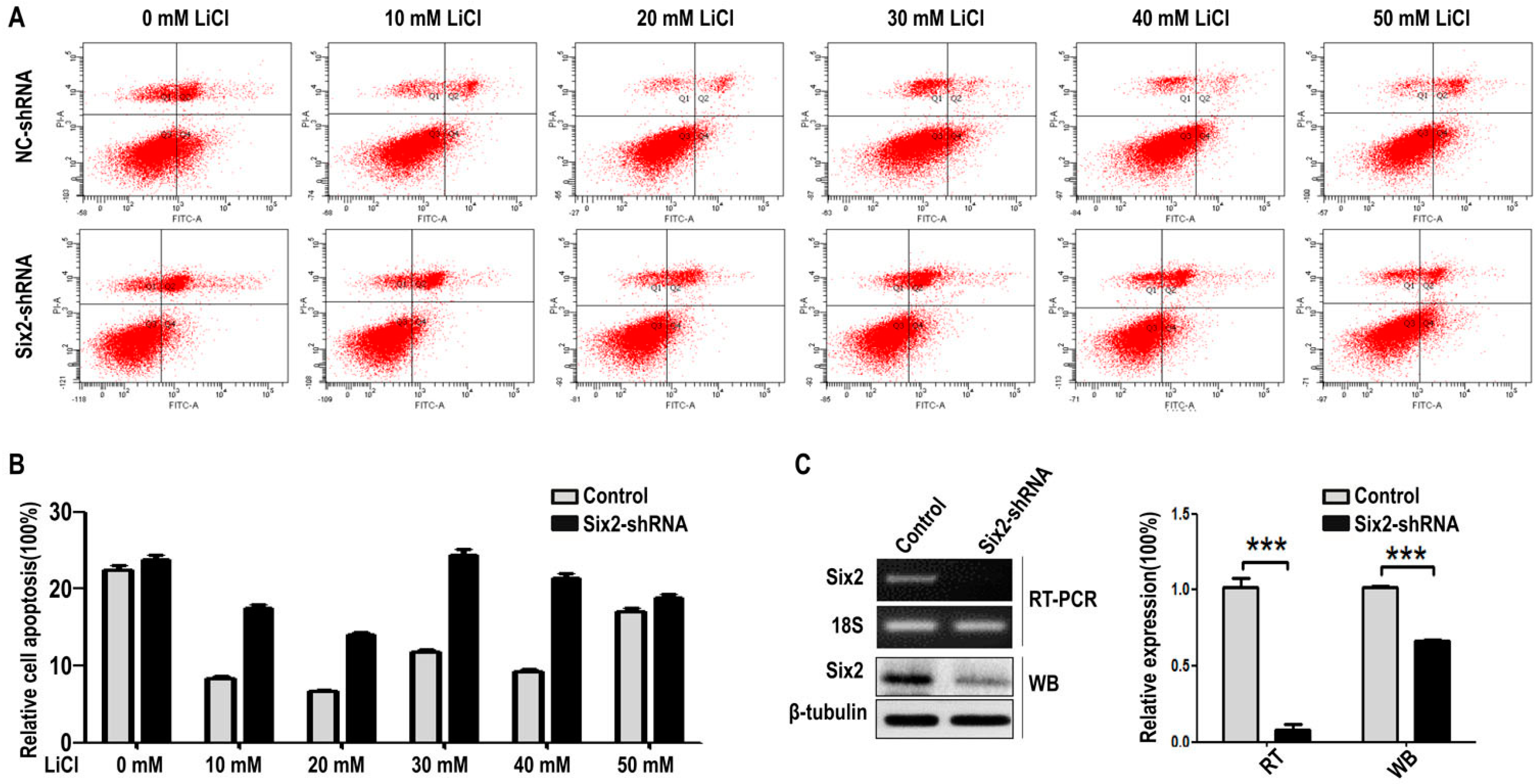
2.3. LiCl Induces the Proliferation and Apoptosis of MM Cells with the Coordination of Six2
3. Discussion
4. Materials and Methods
4.1. Plasmids Construction
4.2. Cell Culture
4.3. Transfection and Luciferase Assays
4.4. Reverse Transcription-PCR (RT-PCR)
4.5. Western Blot
4.6. 5-Ethynyl-20-deoxyuridine (EdU) Assays
4.7. Flow Cytometry Apoptosis Assays
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MM | metanephric mesenchymal |
| UB | ureteric bud |
| Six2 | Sine oculis homeobox homolog 2 |
| LiCl | lithium chloride |
| BMP | bone morphogenetic protein |
| FITC | Fluorescein isothiocyanate fitc |
| HEK | human embryonic kidney |
| MET | mesenchymal-epithelial-transition |
| DMEM | Dulbecco’s modified Eagle’s medium |
References
- Self, M.; Lagutin, O.V.; Bowling, B.; Hendrix, J.; Cai, Y.; Dressler, G.R.; Oliver, G. Six2 is required for suppression of nephrogenesis and progenitor renewal in the developing kidney. EMBO J. 2006, 25, 5214–5228. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Valerius, M.T.; Mugford, J.W.; Carroll, T.J.; Self, M.; Oliver, G.; McMahon, A.P. Six2 defines and regulates a multipotent self-renewing nephron progenitor population throughout mammalian kidney development. Cell Stem Cell 2008, 3, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.Y.; Sims-Lucas, S.; Bushnell, D.S.; Bodnar, A.J.; Kreidberg, J.A.; Ho, J. Dicer function is required in the metanephric mesenchyme for early kidney development. Am. J. Physiol. Ren. Physiol. 2014, 306, F764–F772. [Google Scholar] [CrossRef] [PubMed]
- Klein, G.; Langegger, M.; Goridis, C.; Ekblom, P. Neural cell adhesion molecules during embryonic induction and development of the kidney. Development 1988, 102, 749–761. [Google Scholar] [PubMed]
- Mugford, J.W.; Yu, J.; Kobayashi, A.; McMahon, A.P. High-resolution gene expression analysis of the developing mouse kidney defines novel cellular compartments within the nephron progenitor population. Dev. Biol. 2009, 333, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Ma, W.; O’Brien, L.; Chung, E.; Guo, J.J.; Cheng, J.G.; Valerius, M.T.; Mcmahon, J.; Wong, W.H.; Mcmahon, A. Six2 and Wnt regulate self-renewal and commitment of nephron progenitors through shared gene regulatory networks. Dev. Cell 2012, 23, 637–651. [Google Scholar] [CrossRef] [PubMed]
- lyu, Z.; Mao, Z.; Wang, H.; Fang, Y.; Chen, T.; Wan, Q.; Wang, M.; Wang, N.; Xiao, J.; Wei, H.; et al. MiR-181b targets Six2 and inhibits the proliferation of metanephric mesenchymal cells in vitro. Biochem. Biophys. Res. Commun. 2013, 440, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Wnt/β-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Cadigan, K.M.; Nusse, R. Wnt signaling: A common theme in animal development. Genes Dev. 1997, 11, 3286–3305. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.M.; Liu, Y.G.; Shen, Q.W.; Shi, X.E.; Yang, G.S. Activation canonical Wnt signaling by LiCl induces porcine skeletal muscle satellite cells differentiation into slow muscle. Chin. J. Anim. Sci. 2012, 28, 26–32. [Google Scholar]
- Marija, M.; Vladanka, T.; Jelena, M.; Milena, S. Quercetin and lithium chloride modulate Wnt signaling in pluripotent embryonal carcinoma NT2/D1 cells. Arch. Biol. Sci. 2013, 65, 201–209. [Google Scholar] [CrossRef]
- Kaufmann, L.; Marinescu, G.; Nazarenko, I.; Thiele, W.; Oberle, C.; Sleeman, J.; Blattner, C. LiCl induces TNF-α and FasL production, thereby stimulating apoptosis in cancer cells. Cell Commun. Signal. CCS 2011, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Zanni, G.; Martino, E.D.; Omelyanenko, A.; Andäng, M.; Delle, U.; Elmroth, K.; Blomgren, K. Lithium increases proliferation of hippocampal neural stem/progenitor cells and rescues irradiation-induced cell cycle arrestin vitro. Oncotarget 2015, 6, 365–368. [Google Scholar]
- Laurenz, J.C.; Smith, S.B. Lithium chloride does not inhibit the proliferation of L6 myoblasts by decreasing intracellular free inositol. J. Anim. Sci. 1998, 76, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.P.; Wen, L.B.; Li, J.R.; Wang, Y.; Ni, B.; Wang, R.; Wang, X.; Sun, M.X.; Fan, H.J.; Mao, X. LiCl inhibits PRRSV infection by enhancing Wnt/β-catenin pathway and suppressing inflammatory responses. Antivir. Res. 2015, 117, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Kroeger, P.T., Jr.; Shoue, D.A.; Mezzacappa, F.M.; Gerlach, G.F.; Wingert, R.A.; Schulz, R.A. Knockdown of SCF(Skp2) function causes double-parked accumulation in the nucleus and DNA re-replication in Drosophila plasmatocytes. PLoS ONE 2013, 8, e79019. [Google Scholar] [CrossRef] [PubMed]
- Skytt, D.M.; Klawonn, A.M.; Stridh, M.H.; Pajecka, K.; Patruss, Y.; Quintana-Cabrera, R.; Bolanos, J.P.; Schousboe, A.; Waagepetersen, H.S. siRNA knock down of glutamate dehydrogenase in astrocytes affects glutamate metabolism leading to extensive accumulation of the neuroactive amino acids glutamate and aspartate. Neurochem. Int. 2012, 61, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, W.; Ma, J.; Pan, Y.; Wang, L.; Zhang, W. Polycystin-1 mediates mechanical strain-induced osteoblastic mechanoresponses via potentiation of intracellular calcium and Akt/β-catenin pathway. PLoS ONE 2014, 9, e91730. [Google Scholar] [CrossRef] [PubMed]
- Ying, Y.; Zhu, H.; Liang, Z.; Ma, X.; Li, S. GLP1 protects cardiomyocytes from palmitate-induced apoptosis via Akt/GSK3b/b-catenin pathway. J. Mol. Endocrinol. 2015, 55, 245–262. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Song, H.; Zhong, L.; Yang, R.; Yang, X.Q.; Jiang, K.L.; Liu, B.Z. Lithium chloride promotes apoptosis in human leukemia NB4 cells by inhibiting glycogen synthase kinase-3 β. Int. J. Med. Sci. 2015, 12, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Lee, K.K.; Zhang, L.; Gerton, J.L. Stimulation of mTORC1 with l-leucine rescues defects associated with Roberts syndrome. PLoS Genet. 2013, 9, e1003857. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Sowa, N.; Cardenas, M.E.; Gerton, J.L. l-leucine partially rescues translational and developmental defects associated with zebrafish models of Cornelia de Lange syndrome. Hum. Mol. Genet. 2015, 24, 1540–1555. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Nguyen-McCarty, M.; Hexner, E.O.; Danet-Desnoyers, G.; Klein, P.S. Maintenance of hematopoietic stem cells through regulation of Wnt and mTOR pathways. Nat. Med. 2012, 18, 1778–1785. [Google Scholar] [CrossRef] [PubMed]
- Valerius, M.T.; Patterson, L.T.; Witte, D.P.; Potter, S.S. Microarray analysis of novel cell lines representing two stages of metanephric mesenchyme differentiation. Mech. Dev. 2002, 110, 151–164. [Google Scholar] [CrossRef]
- Hamada, N.; Grimm, C.; Mori, H.; DeGroot, L.J. Identification of a thyroid microsomal antigen by Western blot and immunoprecipitation. J. Clin. Endocrinol. Metab. 1985, 61, 120–128. [Google Scholar] [CrossRef] [PubMed]






© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Ju, P.; Zhou, Y.; Zhao, Y.; Xie, Y.; Long, Y.; Gu, Y.; Ni, D.; Lyv, Z.; Mao, Z.; et al. Six2 Is a Coordinator of LiCl-Induced Cell Proliferation and Apoptosis. Int. J. Mol. Sci. 2016, 17, 1504. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17091504
Liu J, Ju P, Zhou Y, Zhao Y, Xie Y, Long Y, Gu Y, Ni D, Lyv Z, Mao Z, et al. Six2 Is a Coordinator of LiCl-Induced Cell Proliferation and Apoptosis. International Journal of Molecular Sciences. 2016; 17(9):1504. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17091504
Chicago/Turabian StyleLiu, Jianing, Pan Ju, Yuru Zhou, Ya Zhao, Yajun Xie, Yaoshui Long, Yuping Gu, Dongsheng Ni, Zhongshi Lyv, Zhaomin Mao, and et al. 2016. "Six2 Is a Coordinator of LiCl-Induced Cell Proliferation and Apoptosis" International Journal of Molecular Sciences 17, no. 9: 1504. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17091504