A Role of Sp1 Binding Motifs in Basal and Large T-Antigen-Induced Promoter Activities of Human Polyomavirus HPyV9 and Its Variant UF-1


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
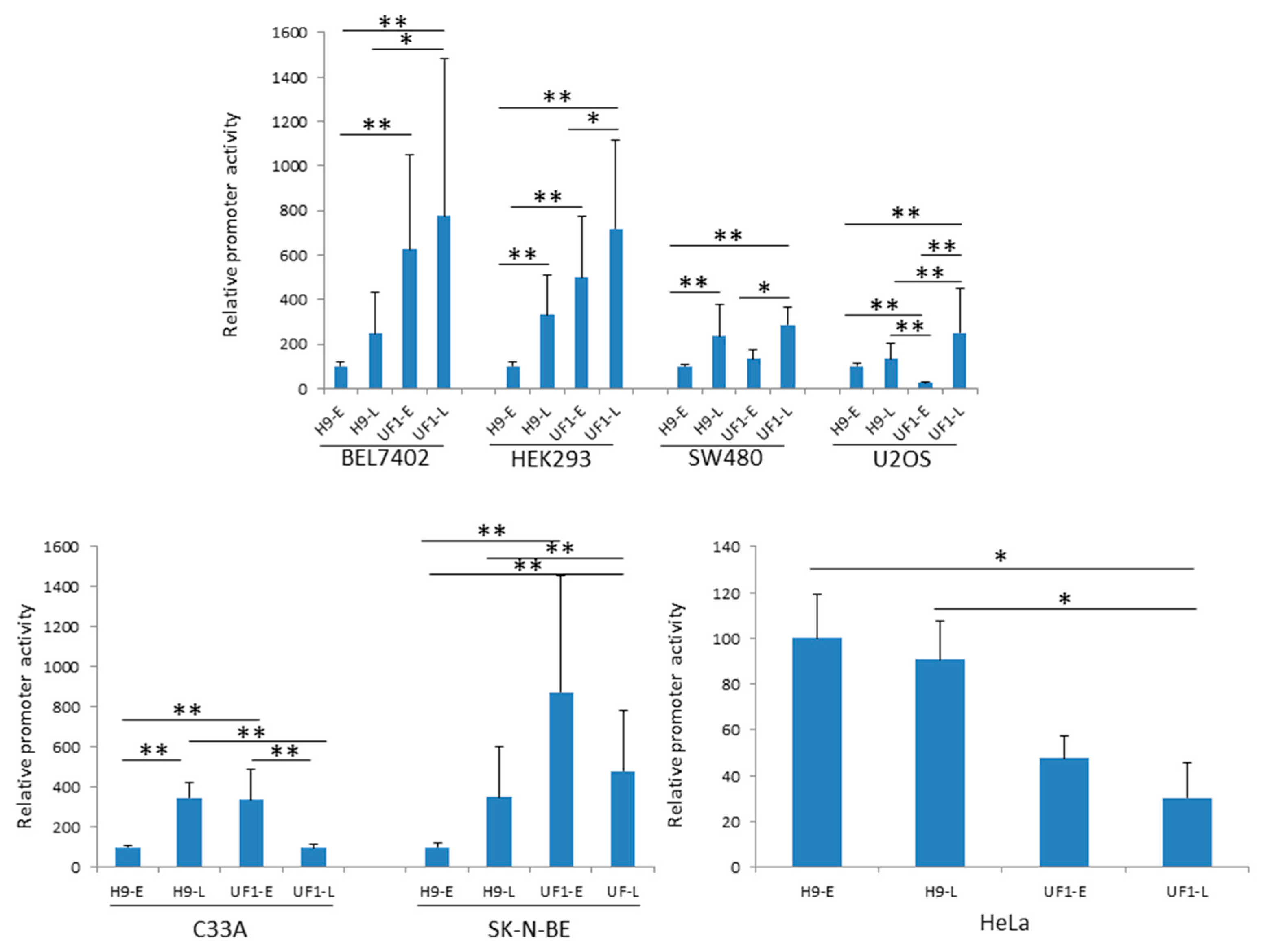
2.1. The Basal UF-1 Early and Late Promoter Activity is Higher than that of HPyV9 in Most Cell Lines Tested
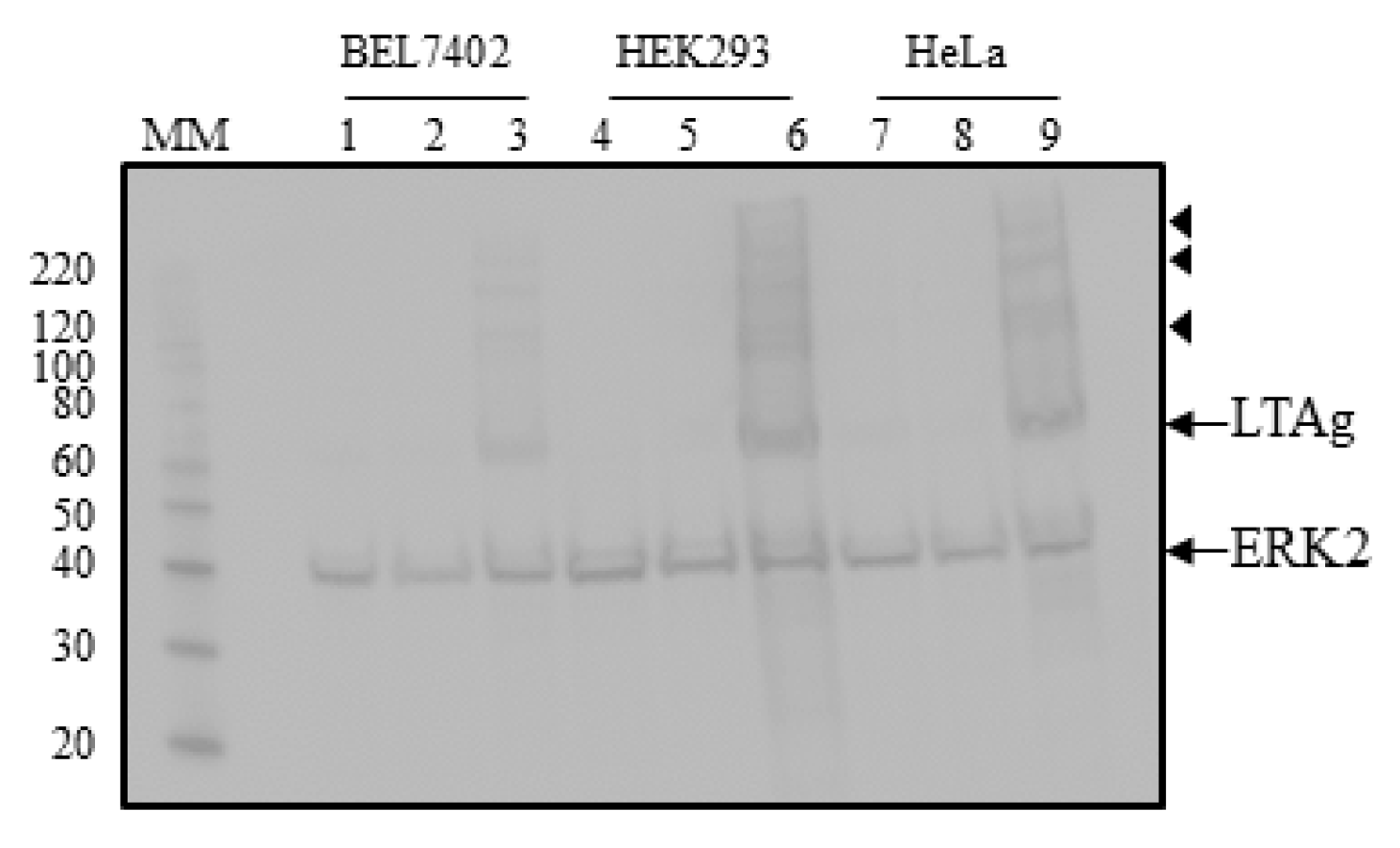
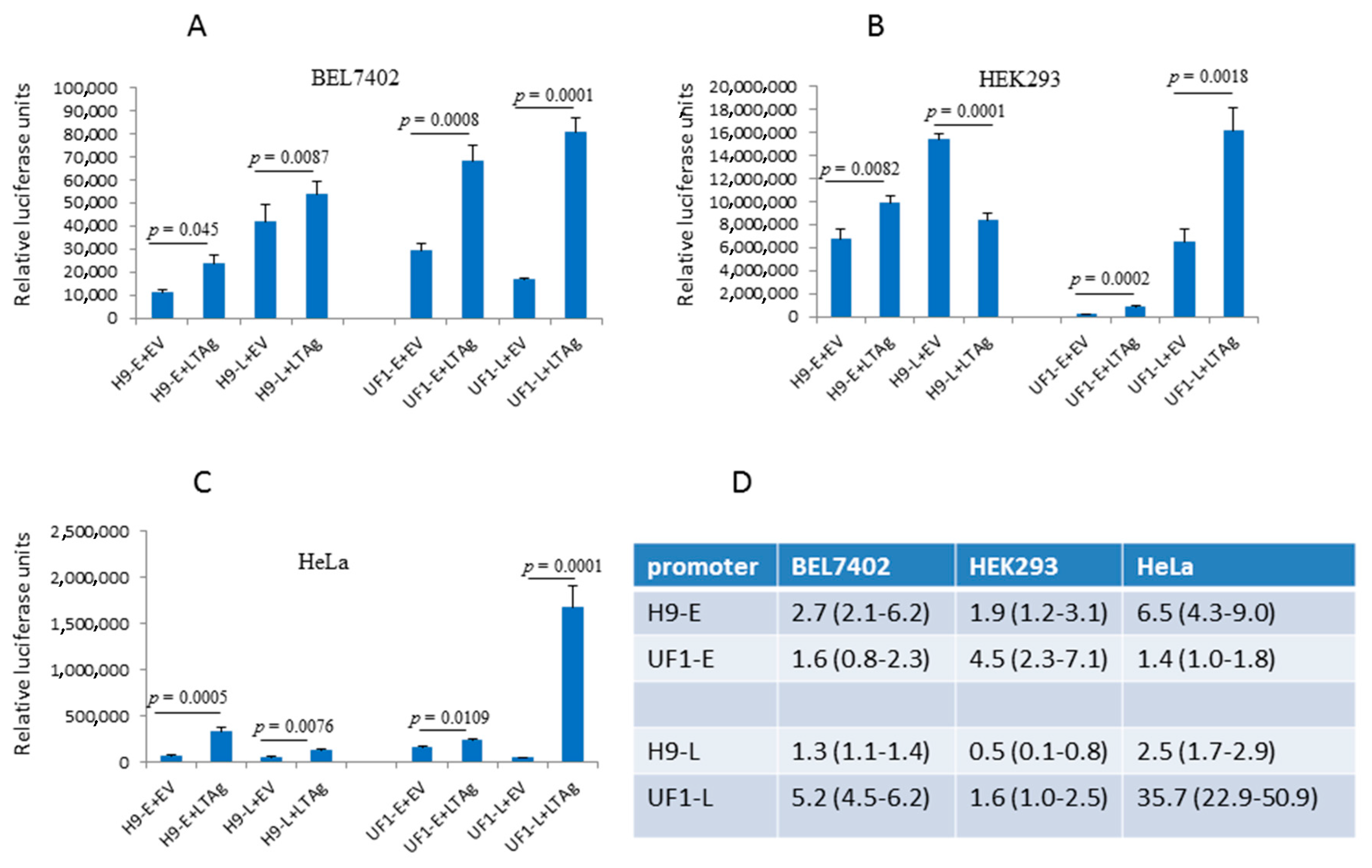
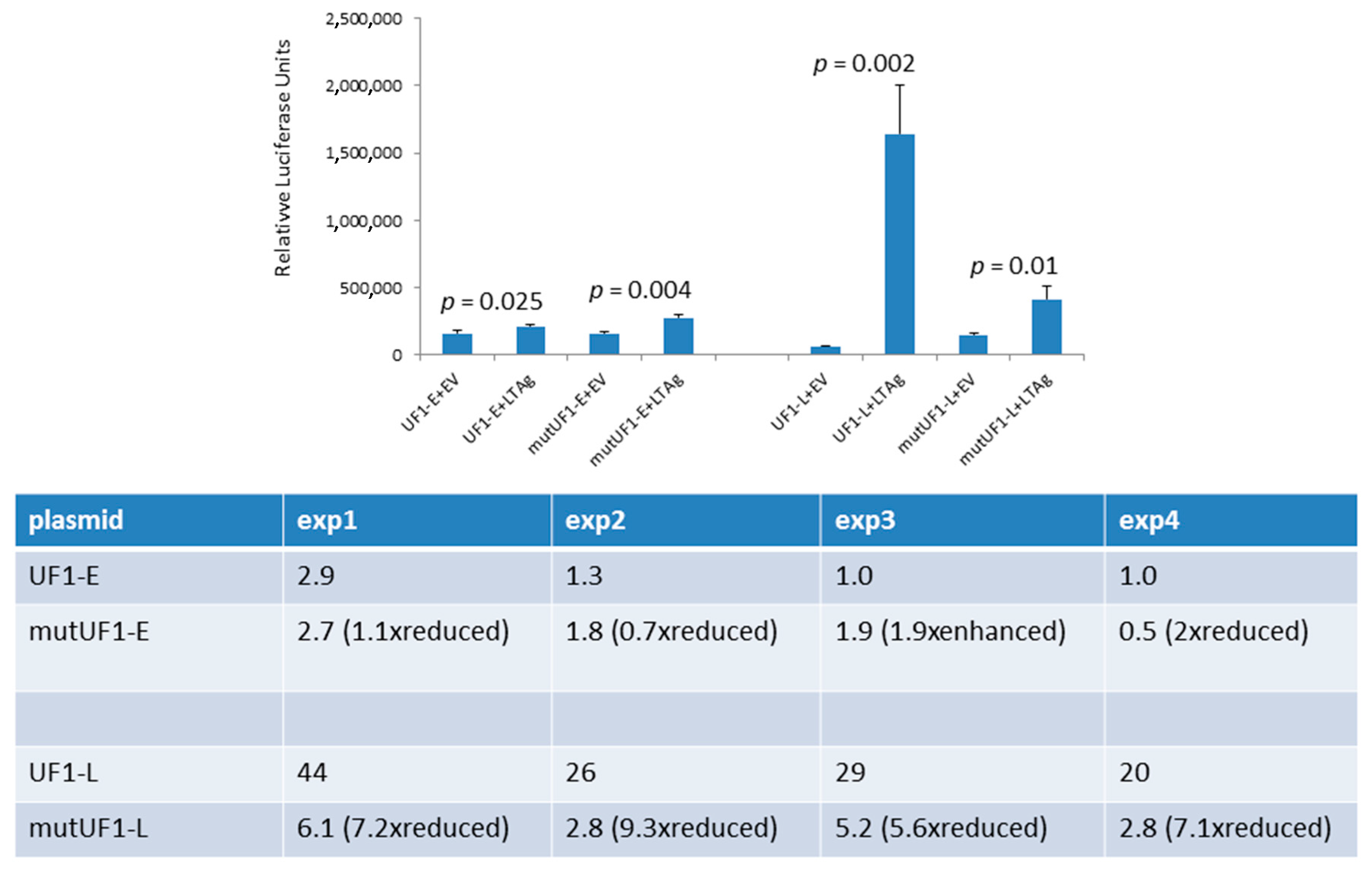
2.2. The UF1 Late Promoter is more Potently Activated by HPyV9 LTAg than the HPyV9 Late Promoter
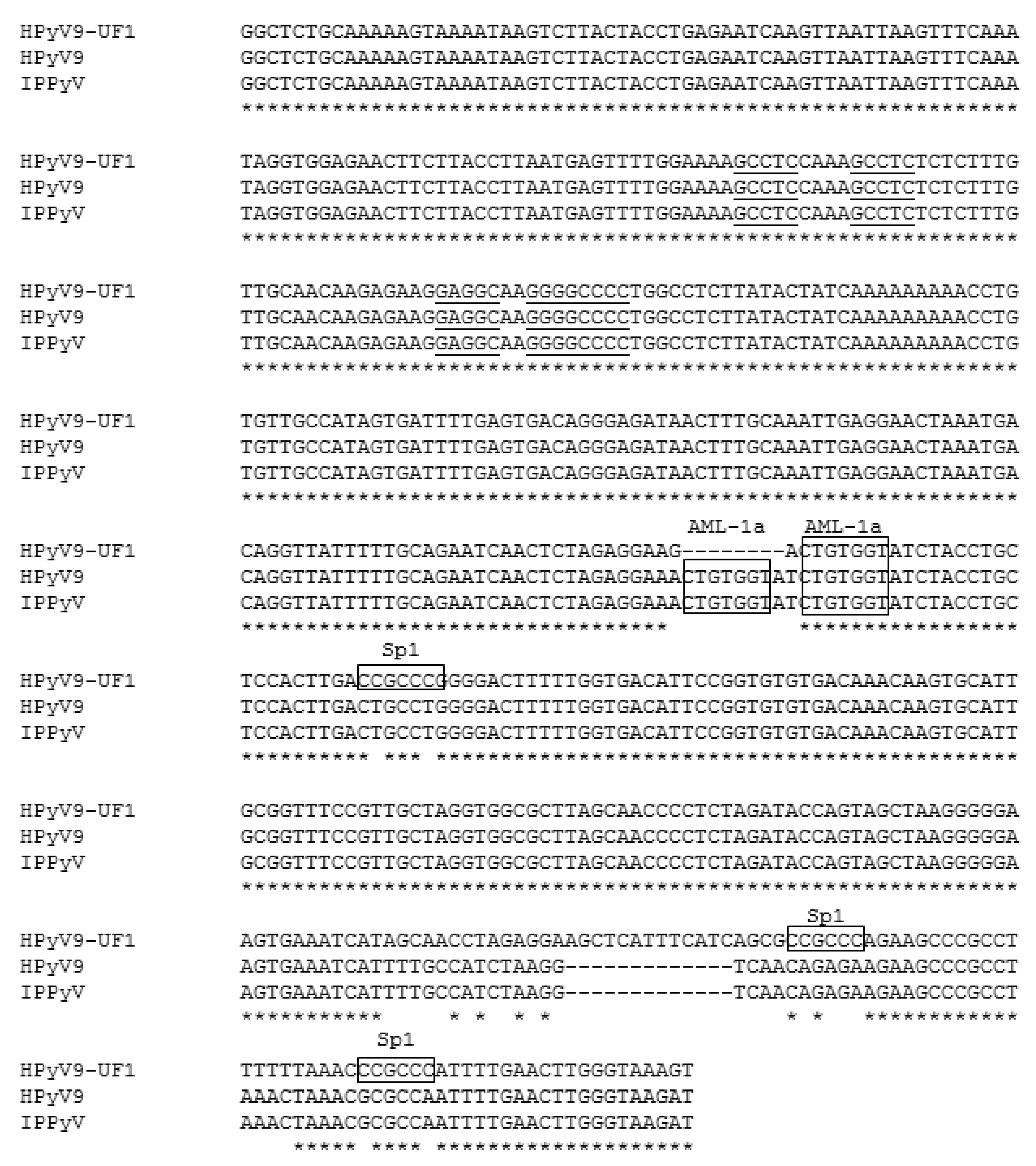
2.3. Sp1 Sites in the Late UF1 Promoter and HPyV9 LTAg
3. Discussions
4. Materials and Methods
4.1. Cell Lines
4.2. Plasmids
4.3. Transfection
4.4. Luciferase Assay
4.5. Protein Concentration Assay
4.6. Western Blotting
4.7. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| EV | empty vector |
| HPyV9 | human polyomavirus 9 |
| H9-E | early promoter HPyV9 |
| H9-L | late promoter HPyV9 |
| LTAg | large T-antigen |
| NCCR | non-coding control region |
| PCR | Polymerase Chain Reaction |
| UF1-E | early promoter UF1 |
| UF1-L | late promoter UF1 |
Appendix A
References
- Moens, U.; Krumbholz, A.; Ehlers, B.; Zell, R.; Johne, R.; Calvignac-Spencer, S.; Lauber, C. Biology, evolution, and medical importance of polyomaviruses: An update. Infect. Genet. Evol. 2017, 54, 18–38. [Google Scholar] [CrossRef] [PubMed]
- Buck, C.B.; Van Doorslaer, K.; Peretti, A.; Geoghegan, E.M.; Tisza, M.J.; An, P.; Katz, J.P.; Pipas, J.M.; McBride, A.A.; Camus, A.C.; et al. The Ancient Evolutionary History of Polyomaviruses. PLoS Pathog. 2016, 12, e1005574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moens, U.; Calvignac-Spencer, S.; Lauber, C.; Ramqvist, T.; Feltkamp, M.C.W.; Daugherty, M.D.; Verschoor, E.J.; Ehlers, B. ICTV Report Consortium. ICTV Virus Taxonomy Profile: Polyomaviridae. J. Gen. Virol. 2017, 98, 1159–1160. [Google Scholar] [PubMed]
- Gardner, S.D.; Field, A.M.; Coleman, D.V.; Hulme, B. New human papovavirus (B.K.) isolated from urine after renal transplantation. Lancet 1971, 1, 1253–1257. [Google Scholar] [CrossRef]
- Padgett, B.L.; Walker, D.L.; ZuRhein, G.M.; Eckroade, R.J.; Dessel, B.H. Cultivation of papova-like virus from human brain with progressive multifocal leucoencephalopathy. Lancet 1971, 1, 1257–1260. [Google Scholar] [CrossRef]
- Allander, T.; Andreasson, K.; Gupta, S.; Bjerkner, A.; Bogdanovic, G.; Persson, M.A.; Dalianis, T.; Ramqvist, T.; Andersson, B. Identification of a third human polyomavirus. J. Virol. 2007, 81, 4130–4136. [Google Scholar] [CrossRef] [PubMed]
- Gheit, T.; Dutta, S.; Oliver, J.; Robitaille, A.; Hampras, S.; Combes, J.D.; McKay-Chopin, S.; Le Calvez-Kelm, F.; Fenske, N.; Cherpelis, B.; et al. Isolation and characterization of a novel putative human polyomavirus. Virology 2017, 506, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Gosert, R.; Rinaldo, C.H.; Funk, G.A.; Egli, A.; Ramos, E.; Drachenberg, C.B.; Hirsch, H.H. Polyomavirus BK with rearranged noncoding control region emerge in vivo in renal transplant patients and increase viral replication and cytopathology. J. Exp. Med. 2008, 205, 841–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosert, R.; Kardas, P.; Major, E.O.; Hirsch, H.H. Rearranged JC virus noncoding control regions found in progressive multifocal leukoencephalopathy patient samples increase virus early gene expression and replication rate. J. Virol. 2010, 84, 10448–10456. [Google Scholar] [CrossRef] [PubMed]
- NCBI Resource Coordinators. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2017, 45, D12–D17. [Google Scholar]
- Song, X.; Van Ghelue, M.; Ludvigsen, M.; Nordbø, S.A.; Ehlers, B.; Moens, U. Characterization of the non-coding control region of polyomavirus KI isolated from nasopharyngeal samples from patients with respiratory symptoms or infection and from blood from healthy blood donors in Norway. J. Gen. Virol. 2016, 97, 1647–1657. [Google Scholar] [CrossRef] [PubMed]
- Moens, U.; Van Ghelue, M.; Ludvigsen, M.; Korup-Schulz, S.; Ehlers, B. Early and late promoters of BKPyV, MCPyV, TSPyV, and HPyV12 are among the strongest of all known human polyomaviruses in 10 different cell lines. J. Gen. Virol. 2015, 96, 2293–2303. [Google Scholar] [CrossRef] [PubMed]
- Scuda, N.; Hofmann, J.; Calvignac-Spencer, S.; Ruprecht, K.; Liman, P.; Kuhn, J.; Hengel, H.; Ehlers, B. A novel human polyomavirus closely related to the african green monkey-derived lymphotropic polyomavirus. J. Virol. 2011, 85, 4586–4590. [Google Scholar] [CrossRef] [PubMed]
- Sauvage, V.; Foulongne, V.; Cheval, J.; Ar Gouilh, M.; Pariente, K.; Dereure, O.; Manuguerra, J.C.; Richardson, J.; Lecuit, M.; Burguière, A.; et al. Human polyomavirus related to African green monkey lymphotropic polyomavirus. Emerg. Infect. Dis. 2011, 17, 1364–1370. [Google Scholar] [CrossRef] [PubMed]
- Trusch, F.; Klein, M.; Finsterbusch, T.; Kuhn, J.; Hofmann, J.; Ehlers, B. Seroprevalence of human polyomavirus 9 and cross-reactivity to African green monkey-derived lymphotropic polyomavirus. J. Gen. Virol. 2012, 93, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Karachaliou, M.; Chatzi, L.; Roumeliotaki, T.; Kampouri, M.; Kyriklaki, A.; Koutra, K.; Chalkiadaki, G.; Michel, A.; Stiaki, E.; Kogevinas, M.; et al. Common infections with polyomaviruses and herpesviruses and neuropsychological development at 4 years of age, the Rhea birth cohort in Crete, Greece. J. Child Psychol. Psychiatry 2016, 57, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Karachaliou, M.; Waterboer, T.; Casabonne, D.; Chalkiadaki, G.; Roumeliotaki, T.; Michel, A.; Stiakaki, E.; Chatzi, L.; Pawlita, M.; Kogevinas, M.; et al. The Natural History of Human Polyomaviruses and Herpesviruses in Early Life—The Rhea Birth Cohort in Greece. Am. J. Epidemiol. 2016, 183, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Gossai, A.; Waterboer, T.; Nelson, H.H.; Michel, A.; Willhauck-Fleckenstein, M.; Farzan, S.F.; Hoen, A.G.; Christensen, B.C.; Kelsey, K.T.; Marsit, C.J.; et al. Seroepidemiology of Human Polyomaviruses in a US Population. Am. J. Epidemiol. 2016, 83, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Sroller, V.; Hamsikova, E.; Ludvikova, V.; Musil, J.; Nemeckova, S.; Salakova, M. Seroprevalence rates of HPyV6, HPyV7, TSPyV, HPyV9, MWPyV and KIPyV polyomaviruses among the healthy blood donors. J. Med. Virol. 2016, 88, 1254–1261. [Google Scholar] [CrossRef] [PubMed]
- Van der Meijden, E.; Wunderink, H.F.; van der Blij-de Brouwer, C.S.; Zaaijer, H.L.; Rotmans, J.I.; Bavinck, J.N.; Feltkamp, M.C. Human polyomavirus 9 infection in kidney transplant patients. Emerg. Infect. Dis. 2014, 20, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Van der Meijden, E.; Bialasiewicz, S.; Rockett, R.J.; Tozer, S.J.; Sloots, T.P.; Feltkamp, M.C. Different serologic behavior of MCPyV, TSPyV, HPyV6, HPyV7 and HPyV9 polyomaviruses found on the skin. PLoS ONE 2013, 8, e81078. [Google Scholar] [CrossRef] [PubMed]
- Nicol, J.T.; Robinot, R.; Carpentier, A.; Carandina, G.; Mazzoni, E.; Tognon, M.; Touzé, A.; Coursaget, P. Age-specific seroprevalences of merkel cell polyomavirus, human polyomaviruses 6, 7, and 9, and trichodysplasia spinulosa-associated polyomavirus. Clin. Vaccine Immunol. 2013, 20, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Nicol, J.T.; Touzé, A.; Robinot, R.; Arnold, F.; Mazzoni, E.; Tognon, M.; Coursaget, P. Seroprevalence and cross-reactivity of human polyomavirus 9. Emerg. Infect. Dis. 2012, 18, 1329–1332. [Google Scholar] [CrossRef] [PubMed]
- Robles, C.; Casabonne, D.; Benavente, Y.; Costas, L.; Gonzalez-Barca, E.; Aymerich, M.; Campo, E.; Tardon, A.; Jiménez-Moleón, J.J.; Castano-Vinyals, G.; et al. Seroreactivity against Merkel cell polyomavirus and other polyomaviruses in chronic lymphocytic leukaemia, the MCC-Spain study. J. Gen. Virol. 2015, 96, 2286–2292. [Google Scholar] [CrossRef] [PubMed]
- Korup-Schulz, S.V.; Lucke, C.; Schmuck, R.; Moens, U.; Ehlers, B. Large T antigen variants of human polyomaviruses 9 and 12 and seroreactivity against their N-terminus. J. Gen. Virol. 2017, 98, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Lednicky, J.A.; Butel, J.S.; Luetke, M.C.; Loeb, J.C. Complete genomic sequence of a new Human polyomavirus 9 strain with an altered noncoding control region. Virus Genes 2014, 49, 490–492. [Google Scholar] [CrossRef] [PubMed]
- Siebrasse, E.A.; Bauer, I.; Holtz, L.R.; Le, B.M.; Lassa-Claxton, S.; Canter, C.; Hmiel, P.; Shenoy, S.; Sweet, S.; Turmelle, Y.; et al. Human polyomaviruses in children undergoing transplantation, United States, 2008–2010. Emerg. Infect. Dis. 2012, 18, 1676–1679. [Google Scholar] [CrossRef] [PubMed]
- Csoma, E.; Sapy, T.; Meszaros, B.; Gergely, L. Novel human polyomaviruses in pregnancy: Higher prevalence of BKPyV, but no WUPyV, KIPyV and HPyV9. J. Clin. Virol. 2012, 55, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Hampras, S.S.; Giuliano, A.R.; Lin, H.Y.; Fisher, K.J.; Abrahamsen, M.E.; McKay-Chopin, S.; Gheit, T.; Tommasino, M.; Rollison, D.E. Natural History of Polyomaviruses in Men: The HPV Infection in Men (HIM) Study. J. Infect. Dis. 2015, 211, 1437–1446. [Google Scholar] [CrossRef] [PubMed]
- Foulongne, V.; Sauvage, V.; Hebert, C.; Dereure, O.; Cheval, J.; Gouilh, M.A.; Pariente, K.; Segondy, M.; Burguière, A.; Manuguerra, J.C.; et al. Human skin microbiota: High diversity of DNA viruses identified on the human skin by high throughput sequencing. PLoS ONE 2012, 7, e38499. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, B.; Wieland, U. The novel human polyomaviruses HPyV6, 7, 9 and beyond. APMIS 2013, 121, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Johnston, S.D.; Yu, X.M.; Mertz, J.E. The major transcriptional transactivation domain of simian virus 40 large T antigen associates nonconcurrently with multiple components of the transcriptional preinitiation complex. J. Virol. 1996, 70, 1191–1202. [Google Scholar] [PubMed]
- Gilinger, G.; Alwine, J.C. Transcriptional activation by simian virus 40 large T antigen: Requirements for simple promoter structures containing either TATA or initiator elements with variable upstream factor binding sites. J. Virol. 1993, 67, 6682–6688. [Google Scholar] [PubMed]
- Briggs, M.R.; Kadonaga, J.T.; Bell, S.P.; Tjian, R. Purification and biochemical characterization of the promoter-specific transcription factor, Sp1. Science 1986, 234, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Messeguer, X.; Escudero, R.; Farre, D.; Nunez, O.; Martinez, J.; Alba, M.M. PROMO: Detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 2002, 18, 333–334. [Google Scholar] [CrossRef] [PubMed]
- Farre, D.; Roset, R.; Huerta, M.; Adsuara, J.E.; Rosello, L.; Alba, M.M.; Messeguer, X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 2003, 31, 3651–3653. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Thul, P.J.; Akesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Björk, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, eaal3321. [Google Scholar] [CrossRef] [PubMed]
- De Luca, P.; Majello, B.; Lania, L. Sp3 represses transcription when tethered to promoter DNA or targeted to promoter proximal RNA. J. Biol. Chem. 1996, 271, 8533–8536. [Google Scholar] [CrossRef] [PubMed]
- Lania, L.; Majello, B.; De Luca, P. Transcriptional regulation by the Sp family proteins. Int. J. Biochem. Cell Biol. 1997, 29, 1313–1323. [Google Scholar] [CrossRef]
- Gupta, T.; Robles, M.T.; Schowalter, R.M.; Buck, C.B.; Pipas, J.M. Expression of the small T antigen of Lymphotropic Papovavirus is sufficient to transform primary mouse embryo fibroblasts. Virology 2016, 487, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Toptan, T.; Yousem, S.A.; Ho, J.; Matsushima, Y.; Stabile, L.P.; Fernandez-Figueras, M.T.; Bhargava, R.; Ryo, A.; Moore, P.S.; Chang, Y. Survey for human polyomaviruses in cancer. JCI Insight 2016, 1, e85562. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.K.; Mann, K. Glycosylation of simian virus 40 T antigen and localization of glycosylated T antigen in the nuclear matrix. Virology 1987, 156, 268–281. [Google Scholar] [CrossRef]
- Klockmann, U.; Deppert, W. Acylation: A new post-translational modification specific for plasma membrane-associated simian virus 40 large T-antigen. FEBS Lett. 1983, 151, 257–259. [Google Scholar] [CrossRef]
- Grasser, F.A.; Scheidtmann, K.H.; Tuazon, P.T.; Traugh, J.A.; Walter, G. In vitro phosphorylation of SV40 large T antigen. Virology 1988, 165, 13–22. [Google Scholar] [CrossRef]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moens, U.; Song, X.; Van Ghelue, M.; Lednicky, J.A.; Ehlers, B. A Role of Sp1 Binding Motifs in Basal and Large T-Antigen-Induced Promoter Activities of Human Polyomavirus HPyV9 and Its Variant UF-1. Int. J. Mol. Sci. 2017, 18, 2414. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18112414
Moens U, Song X, Van Ghelue M, Lednicky JA, Ehlers B. A Role of Sp1 Binding Motifs in Basal and Large T-Antigen-Induced Promoter Activities of Human Polyomavirus HPyV9 and Its Variant UF-1. International Journal of Molecular Sciences. 2017; 18(11):2414. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18112414
Chicago/Turabian StyleMoens, Ugo, Xiaobo Song, Marijke Van Ghelue, John A. Lednicky, and Bernhard Ehlers. 2017. "A Role of Sp1 Binding Motifs in Basal and Large T-Antigen-Induced Promoter Activities of Human Polyomavirus HPyV9 and Its Variant UF-1" International Journal of Molecular Sciences 18, no. 11: 2414. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18112414