Overexpression and Down-Regulation of Barley Lipoxygenase LOX2.2 Affects Jasmonate-Regulated Genes and Aphid Fecundity

Abstract
:
1. Introduction
2. Results
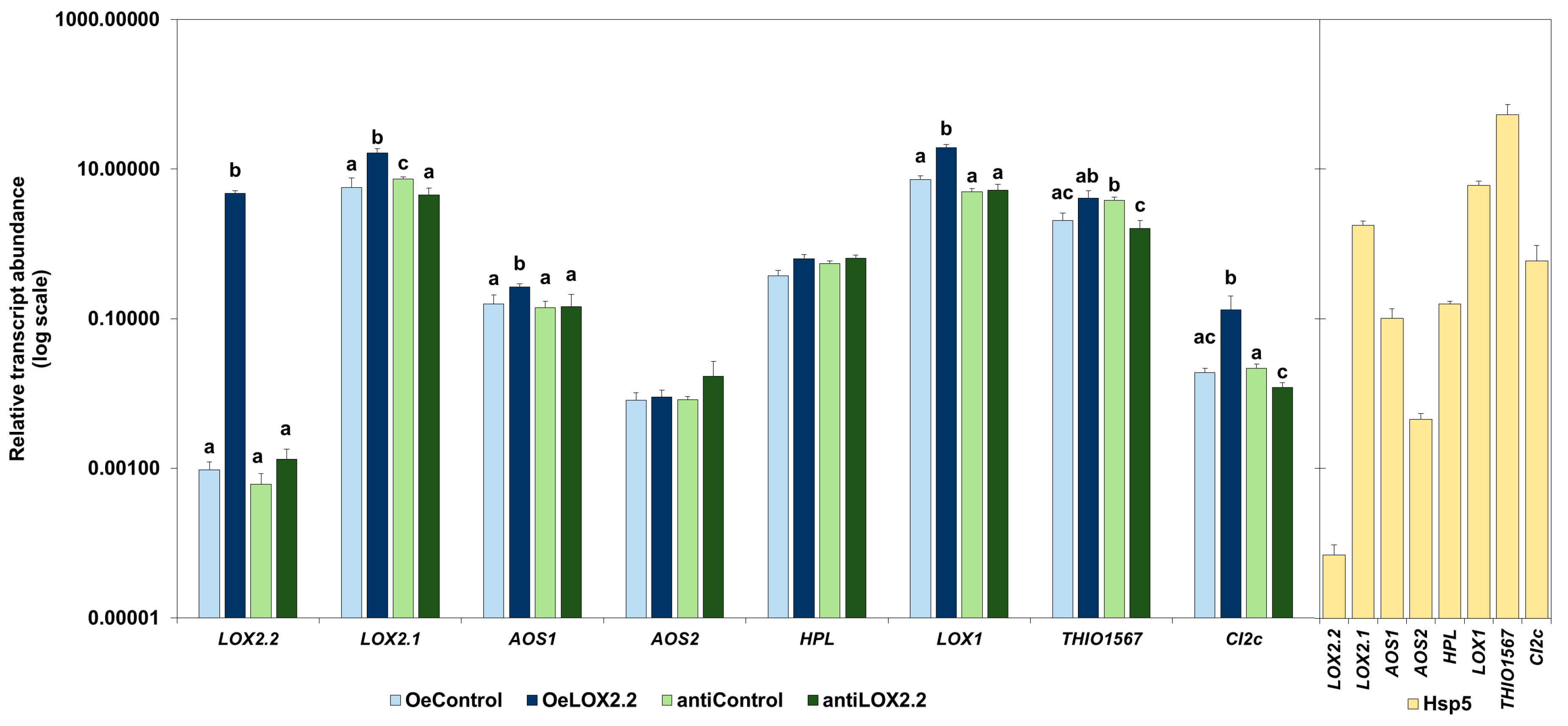
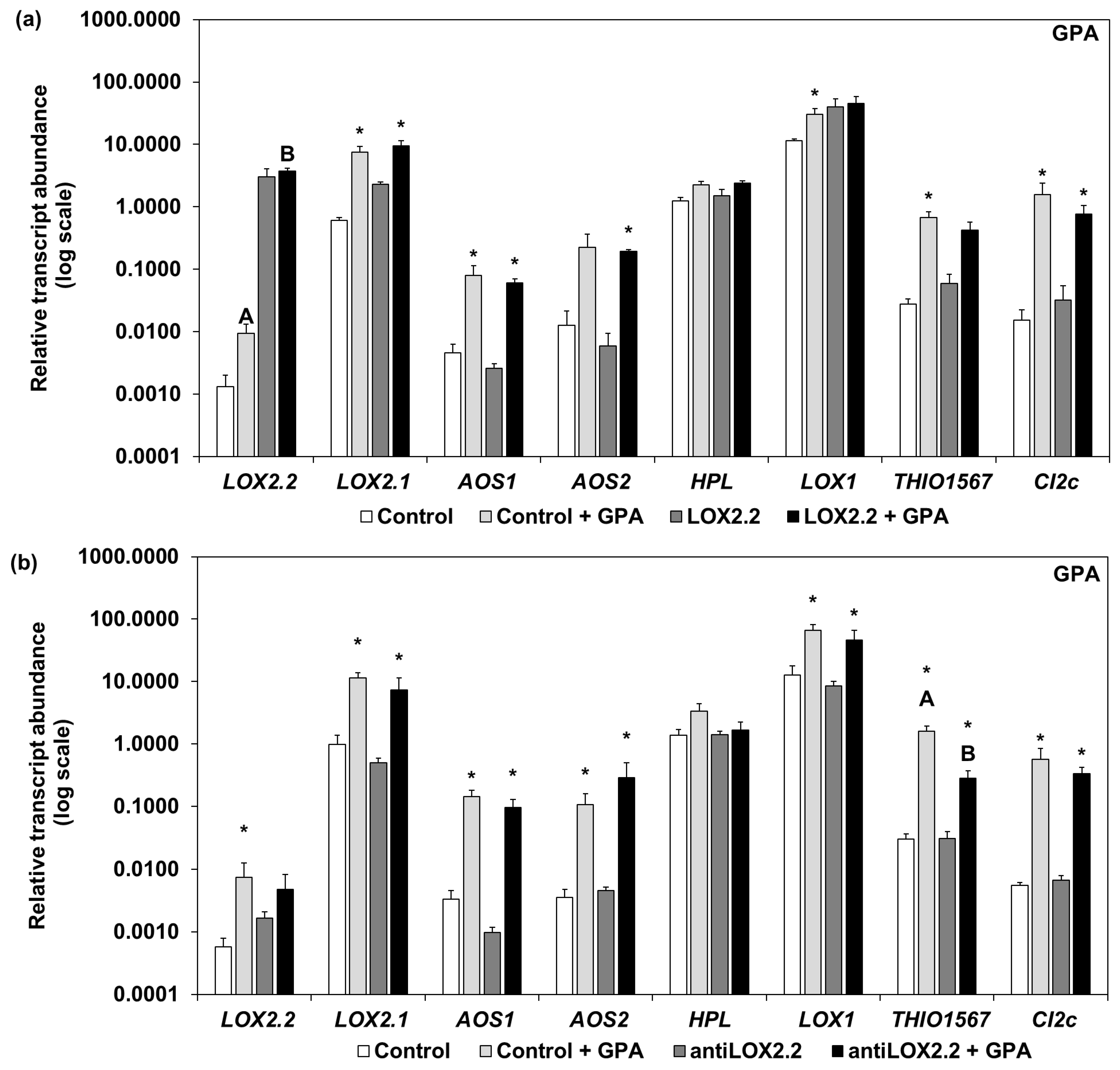
2.1. Phenotypes and Transcript Abundance of Jasmonic Acid (JA)-Regulated Genes
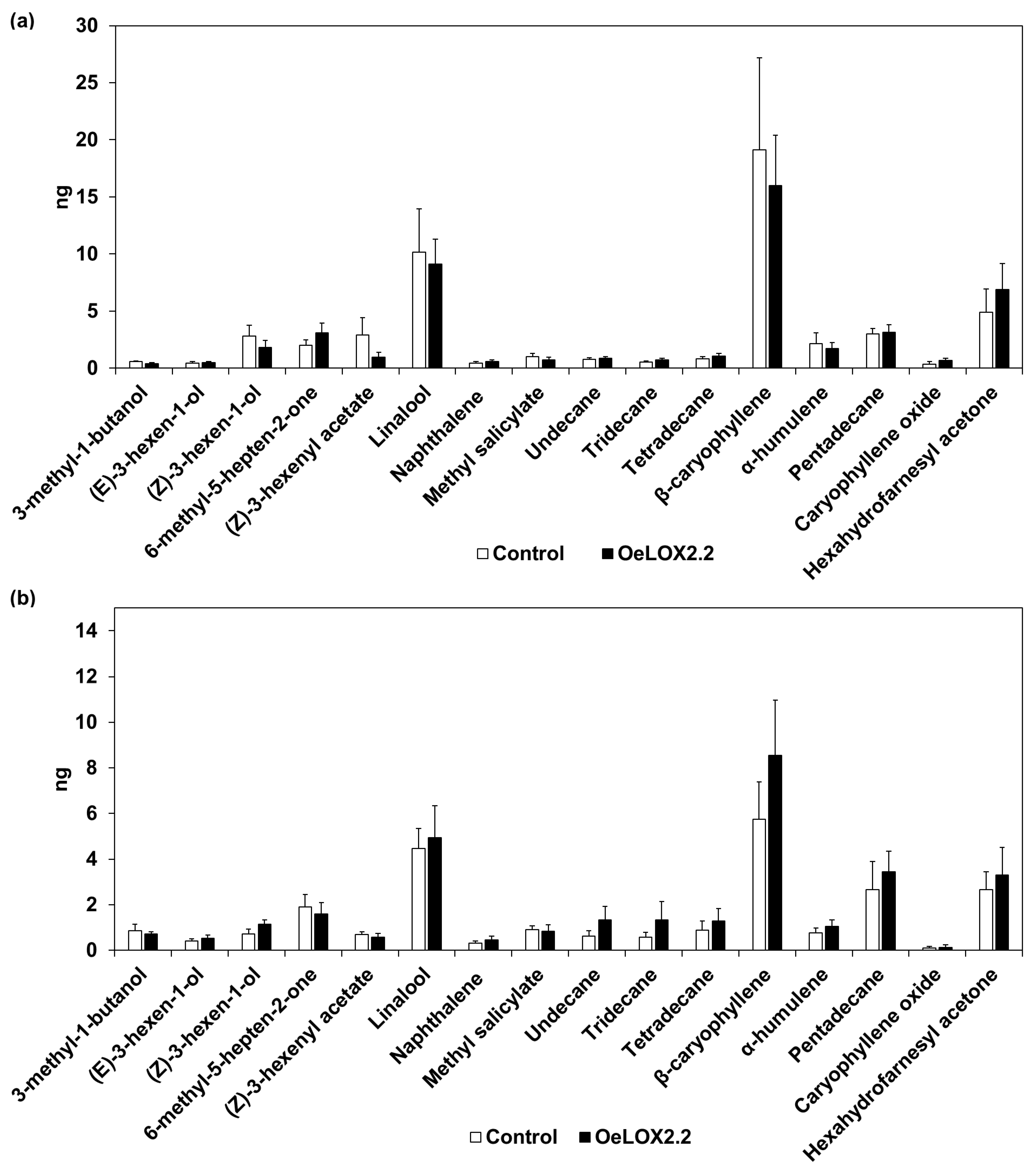
2.2. Volatile Composition with and without Aphids in LOX2.2 Overexpressing Plants
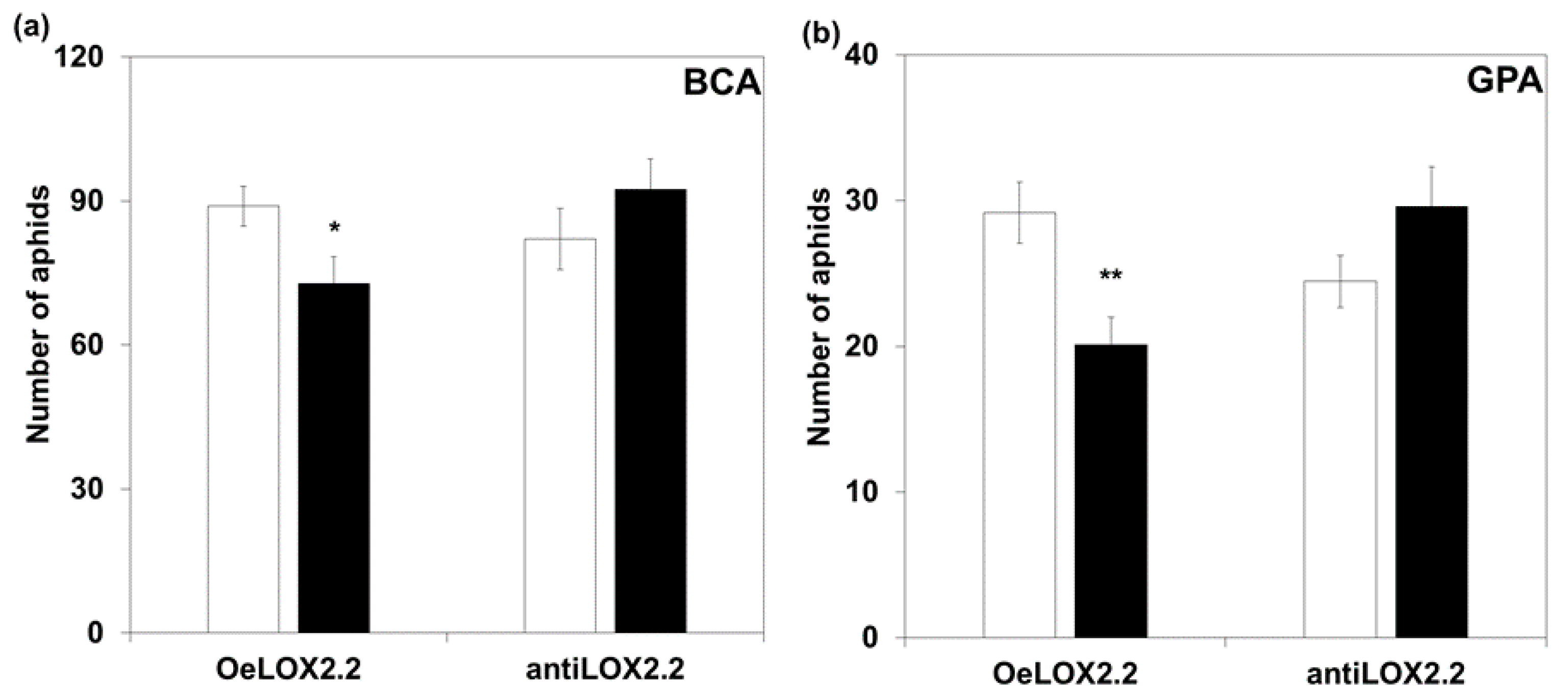
2.3. Bird Cherry-Oat Aphid (BCA) and Green Peach Aphid (GPA) Settling, Fecundity, and Lifespan on LOX2.2 Transformed Plants
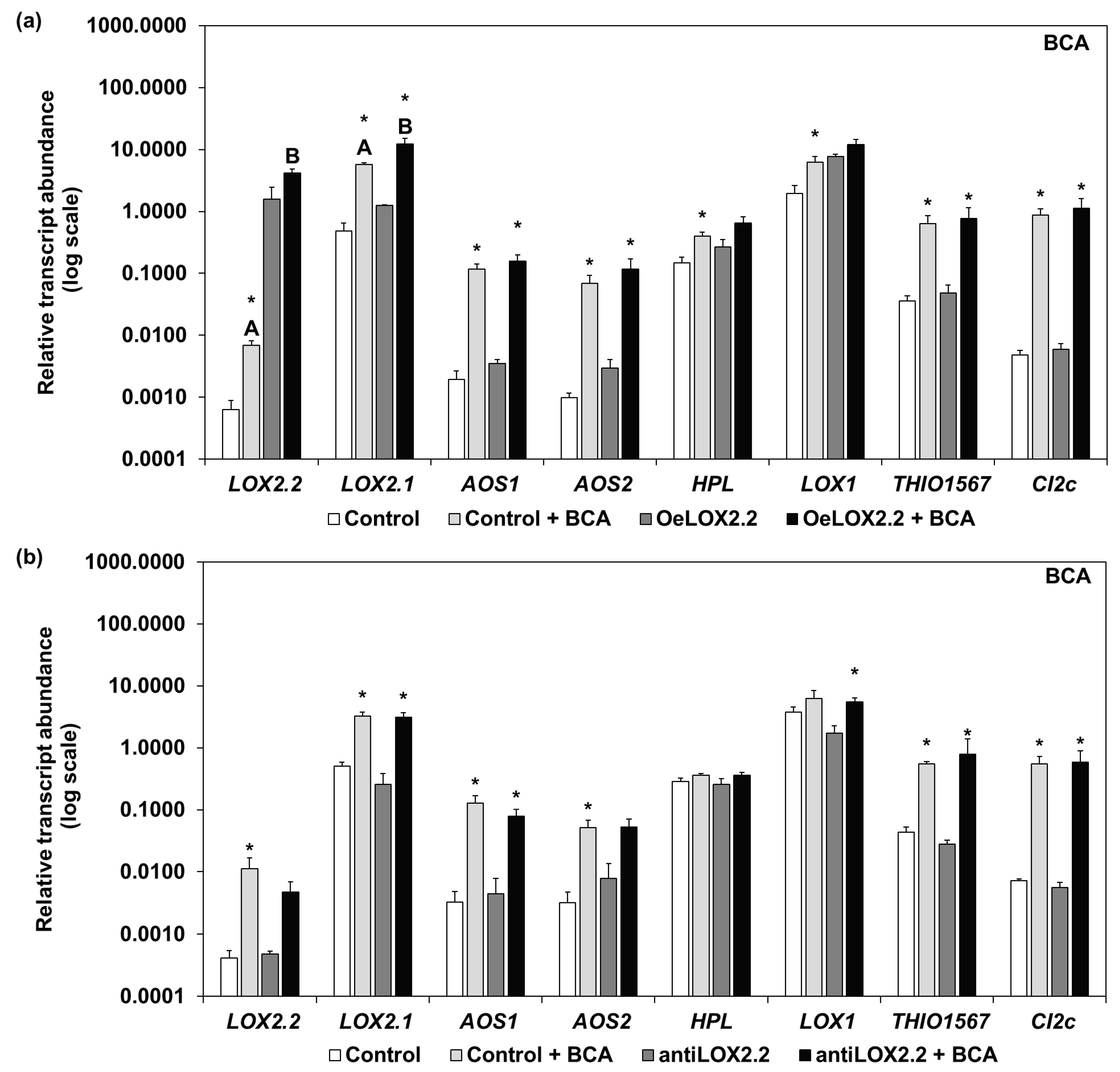
2.4. Transcript Abundance of Genes Regulated by JA in Aphid Infested Tissue
3. Discussion
4. Materials and Methods
4.1. Aphid Rearing
4.2. Plant Cultivation
4.3. Plasmid Constructs, Plant Transformation and Selection
4.4. RNA Extraction and RT-qPCR
4.5. Volatile Collection and Analysis
4.6. Biological Tests with Aphids
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AOS | Allene oxide synthase |
| BCA | Bird cherry-oat aphid |
| GLVs | Green leaf volatiles |
| GPA | Green peach aphid |
| HPL | Hydroperoxide lyase |
| JA | Jasmonic acid |
| LOX | Lipoxygenase |
References
- Quisenberry, S.; Ni, X. Feeding injury. In Aphids as Crop Pests; van Emden, H., Harrington, R., Eds.; CABI Publishing: Wallingford, UK, 2007; pp. 331–352. [Google Scholar]
- Saheed, S.A.; Botha, C.E.J.; Liu, L.; Jonsson, L. Comparison of structural damage caused by Russian wheat aphid (Diuraphis noxia) and Bird cherry-oat aphid (Rhopalosiphum padi) in a susceptible barley cultivar, Hordeum vulgare cv. Clipper. Physiol. Plant. 2007, 129, 429–435. [Google Scholar] [CrossRef]
- Irwin, M.E.; Thresh, J. Epidemiology of barley yellow dwarf virus: A study in ecological complexity. Annu. Rev. Phytopathol. 1990, 28, 393–424. [Google Scholar] [CrossRef]
- Ordon, F.; Perovic, D. Virus resistance in barley. In Translational Genomics for Crop Breeding: Volume 1—Biotic Stress; Varshney, R., Tuberosa, R., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2013; pp. 63–75. [Google Scholar]
- Miller, W.A.; Rasochová, L. Barley yellow dwarf viruses. Annu. Rev. Phytopathol. 1997, 35, 167–190. [Google Scholar] [CrossRef] [PubMed]
- Åhman, I.; Tuvesson, S.; Johansson, M. Does indole alkaloid gramine confer resistance in barley to aphid Rhopalosiphum padi? J. Chem. Ecol. 2000, 26, 233–255. [Google Scholar] [CrossRef]
- Mehrabi, S.; Åhman, I.; Jonsson, L.M.V. Transcript abundance of resistance- and susceptibility-related genes in a barley breeding pedigree with partial resistance to the bird cherry-oat aphid (Rhopalosiphum padi L.). Euphytica 2014, 198, 211–222. [Google Scholar] [CrossRef]
- Delp, G.; Gradin, T.; Åhman, I.; Jonsson, L. Microarray analysis of the interaction between the aphid Rhopalosiphum padi and host plants reveals both differences and similarities between susceptible and partially resistant barley lines. Mol. Genet. Genom. 2009, 281, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Vörös, K.; Feussner, I.; Kühn, H.; Lee, J.; Graner, A.; Löbler, M.; Parthier, B.; Wasternack, C. Characterization of a methyljasmonate-inducible lipoxygenase from barley (Hordeum vulgare cv. Salome) leaves. Eur. J. Biochem. 1998, 251, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, A.; Hause, B.; Maucher, H.; Garbe, E.; Vörös, K.; Weichert, H.; Wasternack, C.; Feussner, I. Jasmonate-induced lipid peroxidation in barley leaves initiated by distinct 13-LOX forms of chloroplasts. Biol. Chem. 2002, 383, 1645–1657. [Google Scholar] [CrossRef] [PubMed]
- Wallis, J.G.; Browse, J. Mutants of Arabidopsis reveal many roles for membrane lipids. Prog. Lipid Res. 2002, 41, 254–278. [Google Scholar] [CrossRef]
- Tovuu, A.; Zulfugarov, I.S.; Wu, G.; Kang, I.S.; Kim, C.; Moon, B.Y.; An, G.; Lee, C.H. Rice mutants deficient in ω-3 fatty acid desaturase (FAD8) fail to acclimate to cold temperatures. Plant Physiol. Biochem. 2016, 109, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Liavonchanka, A.; Feussner, I. Lipoxygenases: Occurrence, functions and catalysis. J. Plant Physiol. 2006, 163, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Andreou, A.; Feussner, I. Lipoxygenases—Structure and reaction mechanism. Phytochemistry 2009, 70, 1504–1510. [Google Scholar] [CrossRef] [PubMed]
- Shibata, D.; Slusarenko, A.; Casey, R.; Bell, E. Lipoxygenases. Plant Mol. Biol. Report. 1994, 12, 41–42. [Google Scholar] [CrossRef]
- Wang, R.; Shen, W.; Liu, L.; Jiang, L.; Liu, Y.; Su, N.; Wan, J. A novel lipoxygenase gene from developing rice seeds confers dual position specificity and responds to wounding and insect attack. Plant Mol. Biol. 2008, 66, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Mosblech, A.; Feussner, I.; Heilmann, I. Oxylipins: Structurally diverse metabolites from fatty acid oxidation. Plant Physiol. Biochem. 2009, 47, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef] [PubMed]
- Lyons, R.; Manners, J.M.; Kazan, K. Jasmonate biosynthesis and signaling in monocots: A comparative overview. Plant Cell Rep. 2013, 32, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Maucher, H.; Hause, B.; Feussner, I.; Ziegler, J.; Wasternack, C. Allene oxide synthases of barley (Hordeum vulgare cv. Salome): Tissue specific regulation in seedling development. Plant J. 2000, 21, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Maucher, H.; Stenzel, I.; Miersch, O.; Stein, N.; Prasad, M.; Zierold, U.; Schweizer, P.; Dorer, C.; Hause, B.; Wasternack, C. The allene oxide cyclase of barley (Hordeum vulgare L.)—Cloning and organ-specific expression. Phytochemistry 2004, 65, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K. Green leaf volatiles: Hydroperoxide lyase pathway of oxylipin metabolism. Curr. Opin. Plant Biol. 2006, 9, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Mwenda, C.M.; Matsui, K. The importance of lipoxygenase control in the production of green leaf volatiles by lipase-dependent and independent pathways. Plant Biotechnol. 2014, 31, 445–452. [Google Scholar] [CrossRef]
- Froehlich, J.E. Tomato allene oxide synthase and fatty acid hydroperoxide lyase, two cytochrome P450s involved in oxylipin metabolism, are targeted to different membranes of chloroplast envelope. Plant Physiol. 2001, 125, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Koeduka, T.; Stumpe, M.; Matsui, K.; Kajiwara, T.; Feussner, I. Kinetics of barley FA hydroperoxide lyase are modulated by salts and detergents. Lipids 2003, 38, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- Scala, A.; Allmann, S.; Mirabella, R.; Haring, M.A.; Schuurink, R.C. Green leaf volatiles: A plant’s multifunctional weapon against herbivores and pathogens. Int. J. Mol. Sci. 2013, 14, 17781–17811. [Google Scholar] [CrossRef] [PubMed]
- Naeem ul Hassan, M.; Zainal, Z.; Ismail, I.; ul Hassan, M.N.; Zainal, Z.; Ismail, I. Green leaf volatiles: Biosynthesis, biological functions and their applications in biotechnology. Plant Biotechnol. J. 2015, 13, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Qi, J.; Ren, N.; Cheng, J.; Erb, M.; Mao, B.; Lou, Y. Silencing OsHI-LOX makes rice more susceptible to chewing herbivores, but enhances resistance to a phloem feeder. Plant J. 2009, 60, 638–648. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Ren, N.; Qi, J.; Lu, J.; Xiang, C.; Ju, H.; Cheng, J.; Lou, Y. The 9-lipoxygenase Osr9-LOX1 interacts with the 13-lipoxygenase-mediated pathway to regulate resistance to chewing and piercing-sucking herbivores in rice. Physiol. Plant. 2014, 152, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Qi, J.; Zhu, X.; Mao, B.; Zeng, L.; Wang, B.; Li, Q.; Zhou, G.; Xu, X.; Lou, Y.; et al. The rice hydroperoxide lyase OsHPL3 functions in defense responses by modulating the oxylipin pathway. Plant J. 2012, 71, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.A.; Nemchenko, A.; Borrego, E.; Murray, I.; Sobhy, I.S.; Bosak, L.; Deblasio, S.; Erb, M.; Robert, C.A.M.; Vaughn, K.A.; et al. The maize lipoxygenase, ZmLOX10, mediates green leaf volatile, jasmonate and herbivore-induced plant volatile production for defense against insect attack. Plant J. 2013, 74, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Weichert, H.; Stenzel, I.; Berndt, E.; Wasternack, C.; Feussner, I. Metabolic profiling of oxylipins upon salicylate treatment in barley leaves—Preferential induction of the reductase pathway by salicylate. FEBS Lett. 1999, 464, 133–137. [Google Scholar] [CrossRef]
- Beβer, K.; Jarosch, B.; Langen, G.; Kogel, K.-H. Expression analysis of genes induced in barley after chemical activation reveals distinct disease resistance pathways. Mol. Plant Pathol. 2000, 1, 277–286. [Google Scholar]
- Walia, H.; Wilson, C.; Condamine, P.; Liu, X.; Ismail, A.M.; Close, T.J. Large-scale expression profiling and physiological characterization of jasmonic acid-mediated adaptation of barley to salinity stress. Plant Cell Environ. 2007, 30, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Losvik, A.; Beste, L.; Mehrabi, S.; Jonsson, L. The protease inhibitor CI2c gene induced by bird cherry-oat aphid in barley inhibits green peach aphid fecundity in transgenic Arabidopsis. Int. J. Mol. Sci. 2017, 18, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Florack, D.E.A.; Stiekema, W.J. Thionins: Properties, possible biological roles and mechanisms of action. Plant Mol. Biol. 1994, 26, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, H.; Apel, K. Thionins. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 227–240. [Google Scholar] [CrossRef]
- Reimann-Philipps, U.; Schrader, G.; Martinoia, E.; Barkholt, V.; Apel, K. Intracellular thionins of barley. J. Biol. Chem. 1989, 264, 8978–8984. [Google Scholar]
- Bate, N.J.; Rothstein, S.J. C6-volatiles derived from the lipoxygenase pathway induce a subset of defense-related genes. Plant J. 1998, 16, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Components of C6-aldehyde-induced resistance in Arabidopsis thaliana against a necrotrophic fungal pathogen, Botritis cinarea. Plant Sci. 2006, 170, 715–723. [Google Scholar] [CrossRef]
- Shiojiri, K.; Kishimoto, K.; Ozawa, R.; Kugimiya, S.; Urashimo, S.; Arimura, G.; Horiuchi, J.; Nishioka, T.; Matsui, K.; Takabayashi, J. Changing green leaf volatile biosynthesis in plants: An approach for improving plant resistance against both herbivores and pathogens. Proc. Natl. Acad. Sci. USA 2006, 103, 16672–16676. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.; Fokar, M.; Abd, H.; Zhang, H.; Allen, R.D.; Paré, P.W. (Z)-3-Hexenol induces defense genes and downstream metabolites in maize. Planta 2005, 220, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Wang, J.; Luo, S. Cloning of the key genes in maize oxylipins pathways and their roles in herbivore induced defense. Chin. Sci. Bull. 2005, 50, 2457–2466. [Google Scholar] [CrossRef]
- Bell, E.; Creelman, R.A.; Mullet, J.E. A chloroplast lipoxygenase is required for wound-induced jasmonic acid accumulation in Arabidopsis. Proc. Natl. Acad. Sci. USA 1995, 92, 8675–8679. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, S.; Sugimoto, K.; Koeduka, T.; Matsui, K. Arabidopsis lipoxygenase 2 is essential for formation of green leaf volatiles and five-carbon volatiles. FEBS Lett. 2016, 590, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Walling, L.L. Avoiding effective defenses: Strategies employed by phloem-feeding insects. Plant Physiol. 2008, 146, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Morkunas, I.; Mai, V.C.; Gabryś, B. Phytohormonal signaling in plant responses to aphid feeding. Acta Physiol. Plant. 2011, 33, 2057–2073. [Google Scholar] [CrossRef]
- Nalam, V.J.; Keeretaweep, J.; Sarowar, S.; Shah, J. Root-derived oxylipins promote green peach aphid performance on Arabidopsis foliage. Plant Cell 2012, 24, 1643–1653. [Google Scholar] [CrossRef] [PubMed]
- Chehab, E.W.; Kaspi, R.; Savchenko, T.; Rowe, H.; Negre-Zakharov, F.; Kliebenstein, D.; Dehesh, K. Distinct roles of jasmonates and aldehydes in plant-defense responses. PLoS ONE 2008, 3, e1904. [Google Scholar] [CrossRef] [PubMed]
- Kettles, G.J.; Drurey, C.; Schoonbeek, H.; Maule, A.J.; Hogenhout, S. Resistance of Arabidopsis thaliana to the green peach aphid, Myzus persicae, involves camalexin and is regulated by microRNAs. New Phytol. 2013, 198, 1178–1190. [Google Scholar] [CrossRef] [PubMed]
- Hellens, R.; Mullineaux, P.; Klee, H. A guide to Agrobacterium binary Ti vectors. Trends Plant Sci. 2000, 5, 446–451. [Google Scholar] [CrossRef]
- Bartlett, J.G.; Alves, S.C.; Smedley, M.; Snape, J.W.; Harwood, W.A. High-throughput Agrobacterium-mediated barley transformation. Plant Methods 2008, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Glinwood, R.; Ahmed, E.; Qvarfordt, E.; Ninkovic, V. Olfactory learning of plant genotypes by a polyphagous insect predator. Oecologia 2011, 166, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, I.I.J.; White, P.F.P. Simple estimation of intrinsic increase rates for aphids and tetranychid mites. J. Appl. Ecol. 1977, 14, 757–766. [Google Scholar] [CrossRef]
- Web Statistical Calculators for Categorical Data Analysis. Available online: www.astatsa.com (accessed on 3 October 2017).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparisons between Plants with and without BCA | Difference | p-Value |
| Sum of LOX derived volatiles: (E)-3-hexen-1-ol, (Z)-3-hexen-1-ol and (Z)-3-hexenyl acetate | 0.049 | |
| (Z)-3-hexen-1-ol | 0.029 | |
| β-Caryophyllene | 0.044 | |
| Linalool | 0.046 | |
| Comparisons within control line with and without BCA | ||
| Sum of LOX derived volatiles: (E)-3-hexen-1-ol, (Z)-3-hexen-1-ol and (Z)-3-hexenyl acetate | −4.34 | 0.028 |
| (Z)-3-hexen-1-ol | −2.07 | 0.022 |
| BCA | Control (n = 6) | OeLOX2.2 (n = 6) | Control (n = 6) | antiLOX2.2 (n = 6) |
| Pre-reproductive days | 6.0 ± 0.0 | 6.2 ± 0.2 | 6.2 ± 0.2 | 6.2 ± 0.2 |
| Number of nymphs/aphid | 58.7 ± 2.3 | 60.0 ± 5.1 | 52.2 ± 6.1 | 57.8 ± 3.4 |
| Nymphs/reproductive day | 4.95 ± 0.29 | 4.56 ± 0.29 | 4.55 ± 0.57 | 5.07 ± 0.34 |
| Reproductive life (days) | 12.0 ± 0.6 | 13.2 ± 0.9 | 11.5 ± 0.2 | 11.5 ± 0.6 |
| Life span (days) | 26.8 ± 1.8 | 29.2 ± 2.3 | 26.8 ± 1.7 | 30.3 ± 1.7 |
| rm | 0.43 ± 0.01 | 0.41 ± 0.012 | 0.41 ± 0.022 | 0.42 ± 0.008 |
| GPA | Control (n = 8) | OeLOX2.2 (n = 5) | Control (n = 6) | antiLOX2.2 (n = 8) |
| Pre-reproductive days | 10.3 ± 0.6 | 10.8 ± 0.7 | 12.0 ± 0.6 | 11.6 ± 0.6 |
| Number of nymphs/aphid | 17.7 ± 3.0 | 18.2 ± 2.4 | 17.8 ± 1.2 | 18.3 ± 3.2 |
| Nymphs/reproductive day | 1.03 ± 0.11 | 1.06 ± 0.11 | 1.0 ± 0.1 | 0.97 ± 0.13 |
| Reproductive life (days) | 16.9 ± 1.5 | 17.0 ± 0.9 | 18.2 ± 1.1 | 18.0 ± 1.3 |
| Life span (days) | 33.1 ± 1.8 | 30.8 ± 2.4 | 38.5 ± 1.6 | 33.8 ± 1.6 |
| rm | 0.17 ± 0.013 | 0.17 ± 0.013 | 0.15 ± 0.009 | 0.15 ± 0.015 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Losvik, A.; Beste, L.; Glinwood, R.; Ivarson, E.; Stephens, J.; Zhu, L.-H.; Jonsson, L. Overexpression and Down-Regulation of Barley Lipoxygenase LOX2.2 Affects Jasmonate-Regulated Genes and Aphid Fecundity. Int. J. Mol. Sci. 2017, 18, 2765. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18122765
Losvik A, Beste L, Glinwood R, Ivarson E, Stephens J, Zhu L-H, Jonsson L. Overexpression and Down-Regulation of Barley Lipoxygenase LOX2.2 Affects Jasmonate-Regulated Genes and Aphid Fecundity. International Journal of Molecular Sciences. 2017; 18(12):2765. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18122765
Chicago/Turabian StyleLosvik, Aleksandra, Lisa Beste, Robert Glinwood, Emelie Ivarson, Jennifer Stephens, Li-Hua Zhu, and Lisbeth Jonsson. 2017. "Overexpression and Down-Regulation of Barley Lipoxygenase LOX2.2 Affects Jasmonate-Regulated Genes and Aphid Fecundity" International Journal of Molecular Sciences 18, no. 12: 2765. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18122765