ITGB1b-Deficient Rare Minnows Delay Grass Carp Reovirus (GCRV) Entry and Attenuate GCRV-Triggered Apoptosis

 , ,
, ,
Abstract
:1. Introduction
2. Results
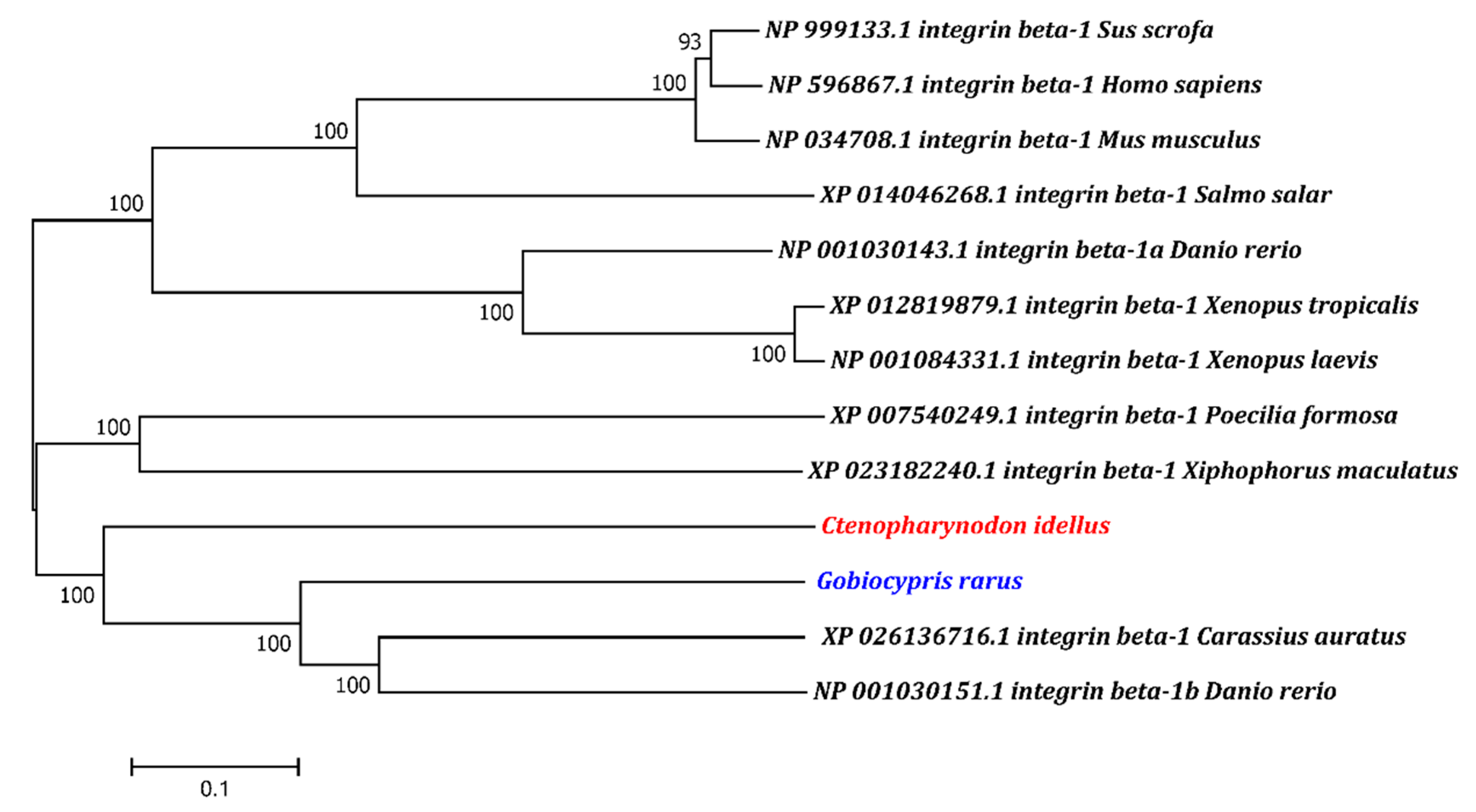
2.1. Characterization and Phylogenetic Analysis of the ITGB1b Gene
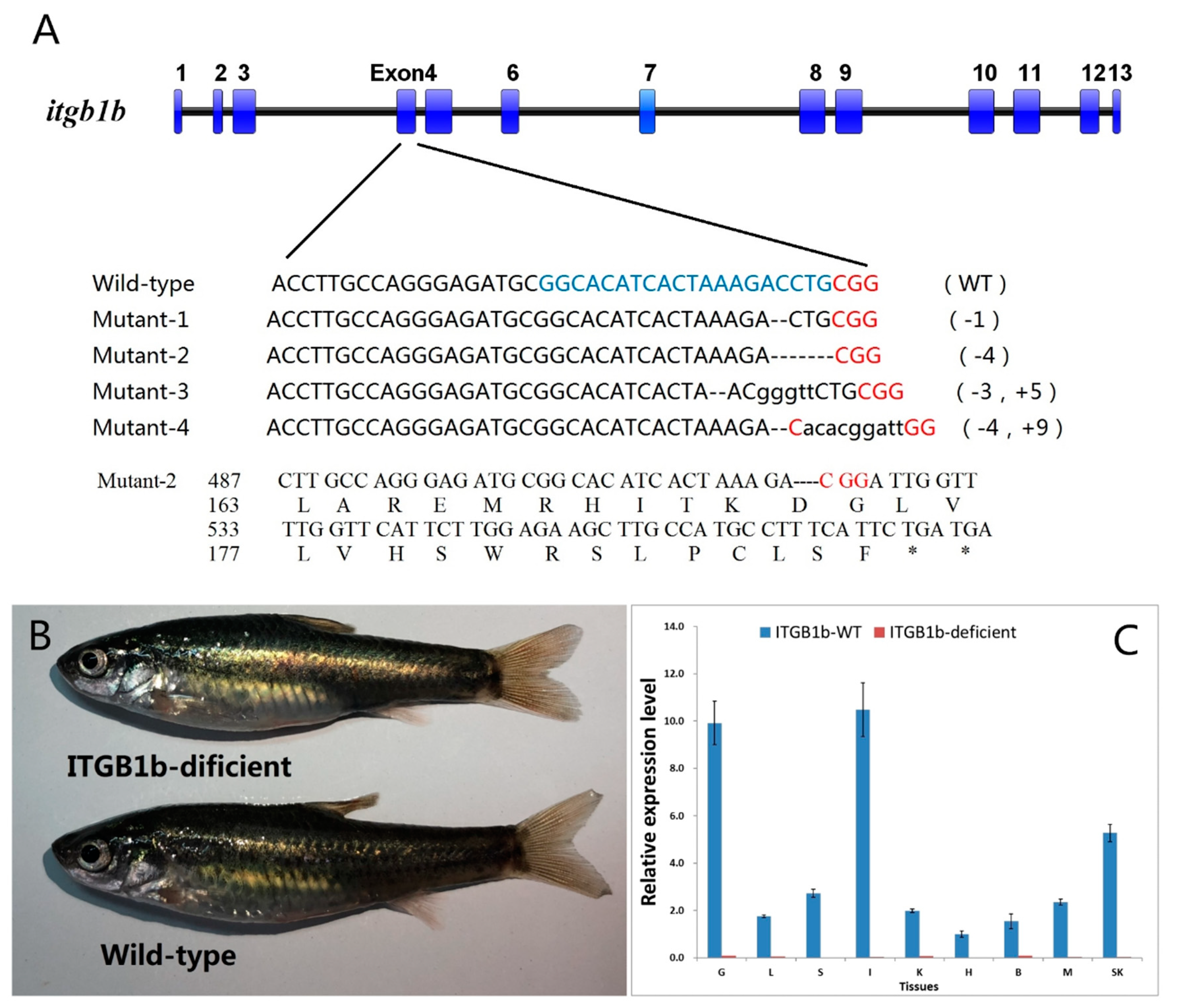
2.2. Generation of an ITGB1b-Deficient Rare Minnow Using the CRISPR/Cas9 System
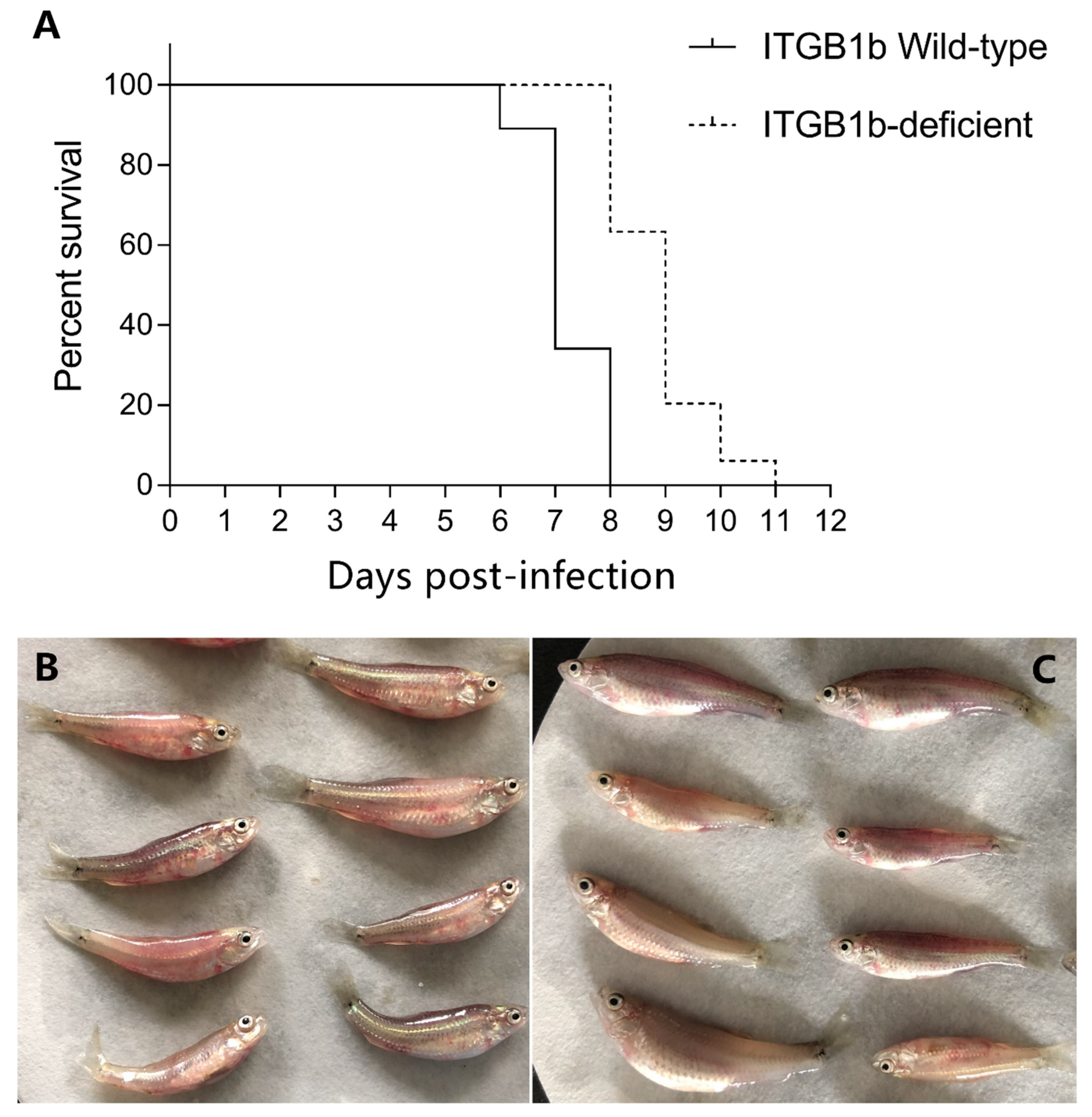
2.3. ITGB1-Deficient Rare Minnow Exhibits Delayed Death Following GCRV Infection
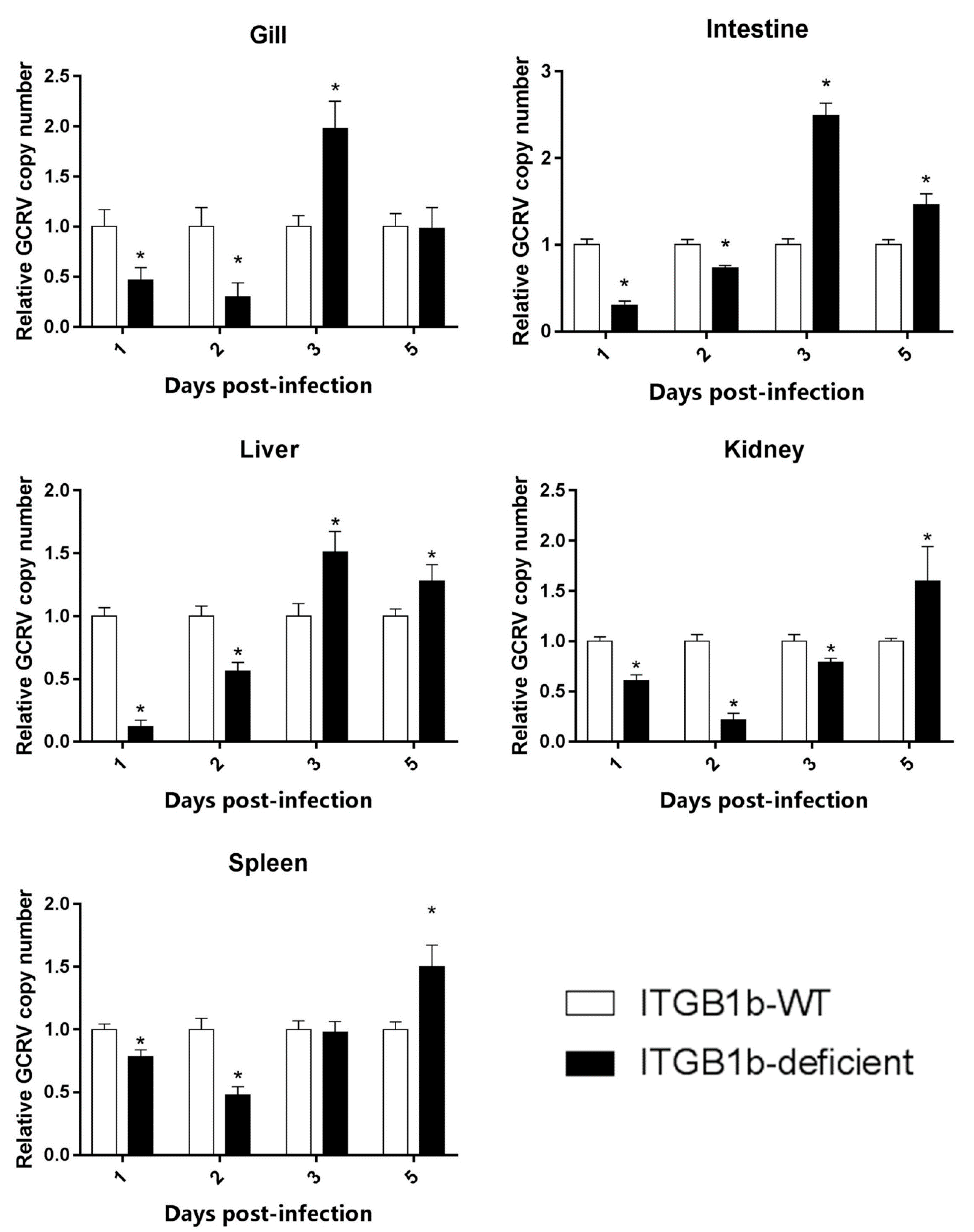
2.4. The Efficiency of GCRV Entry Is Reduced in the ITGB1b-Deficient Rare Minnow
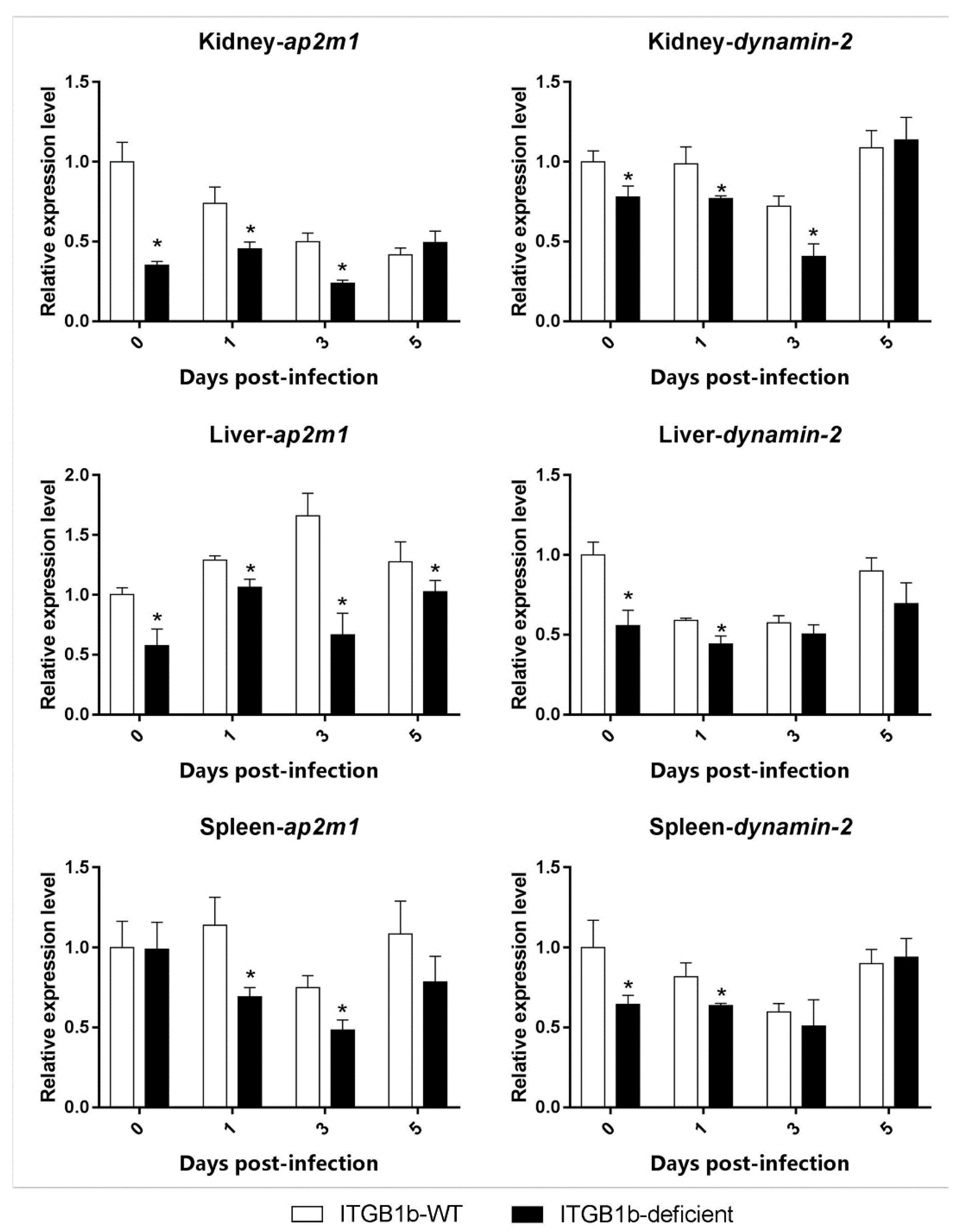
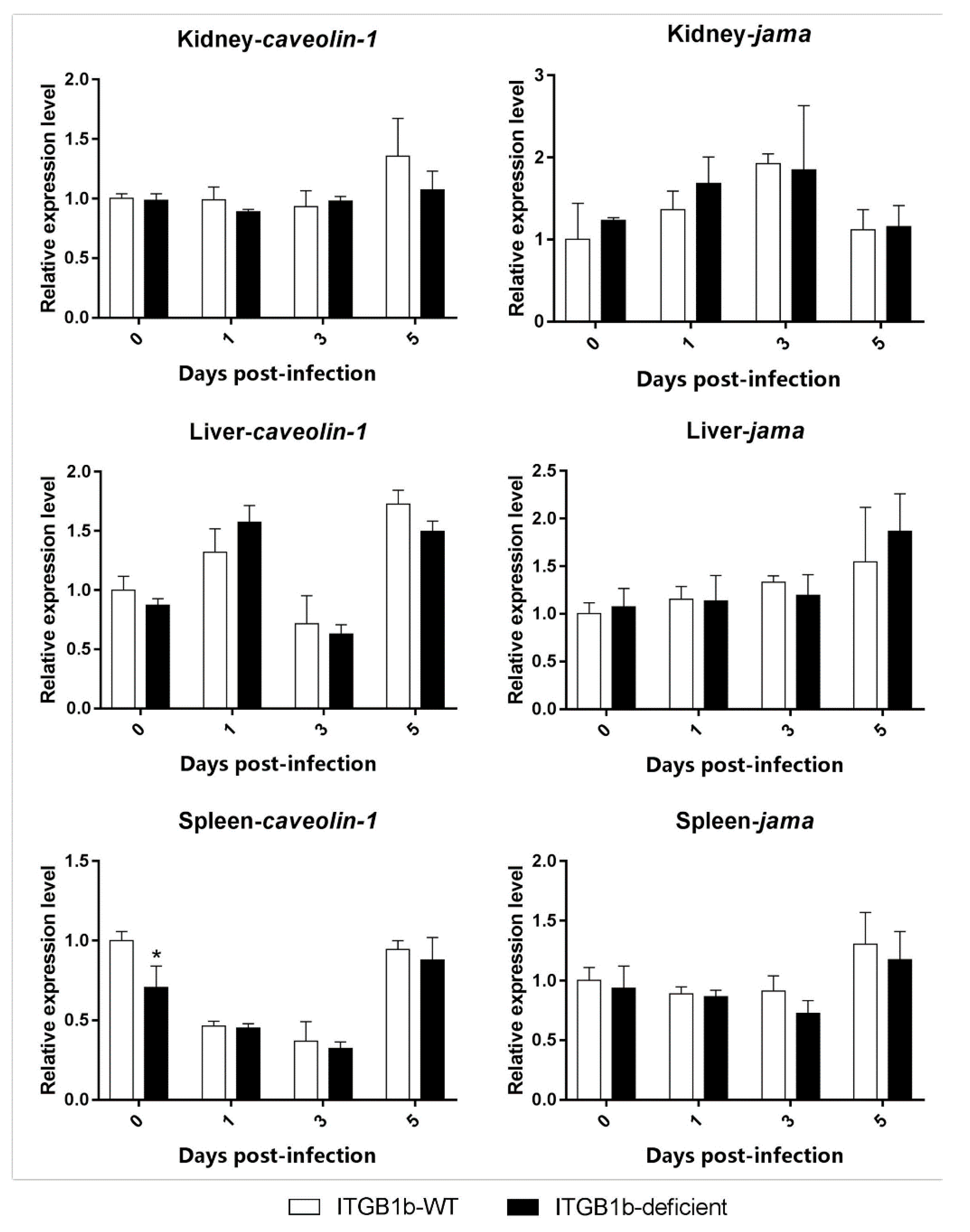
2.5. Clathrin-Mediated Endocytosis Is Attenuated in the ITGB1b-Deficient Rare Minnow
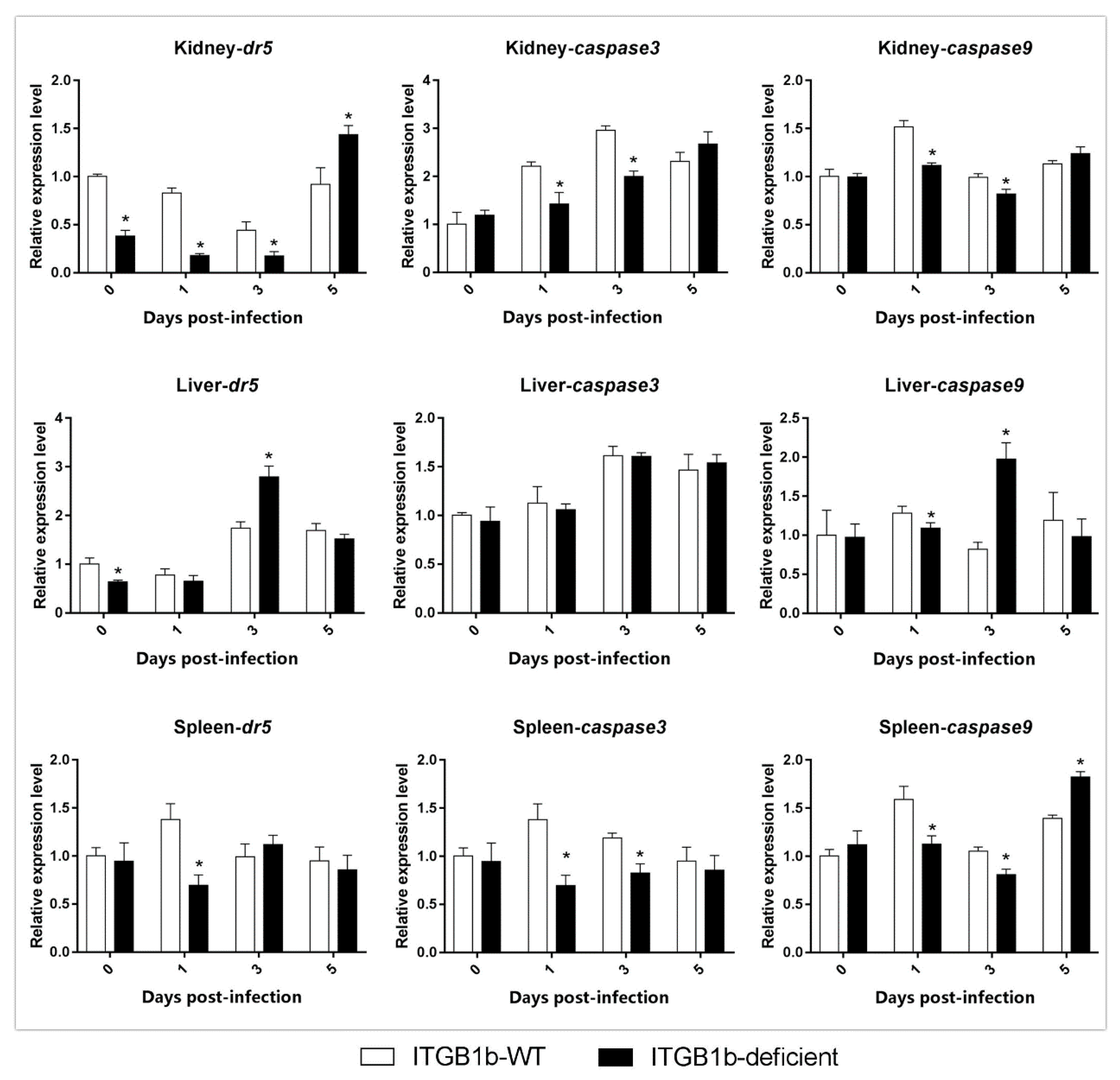
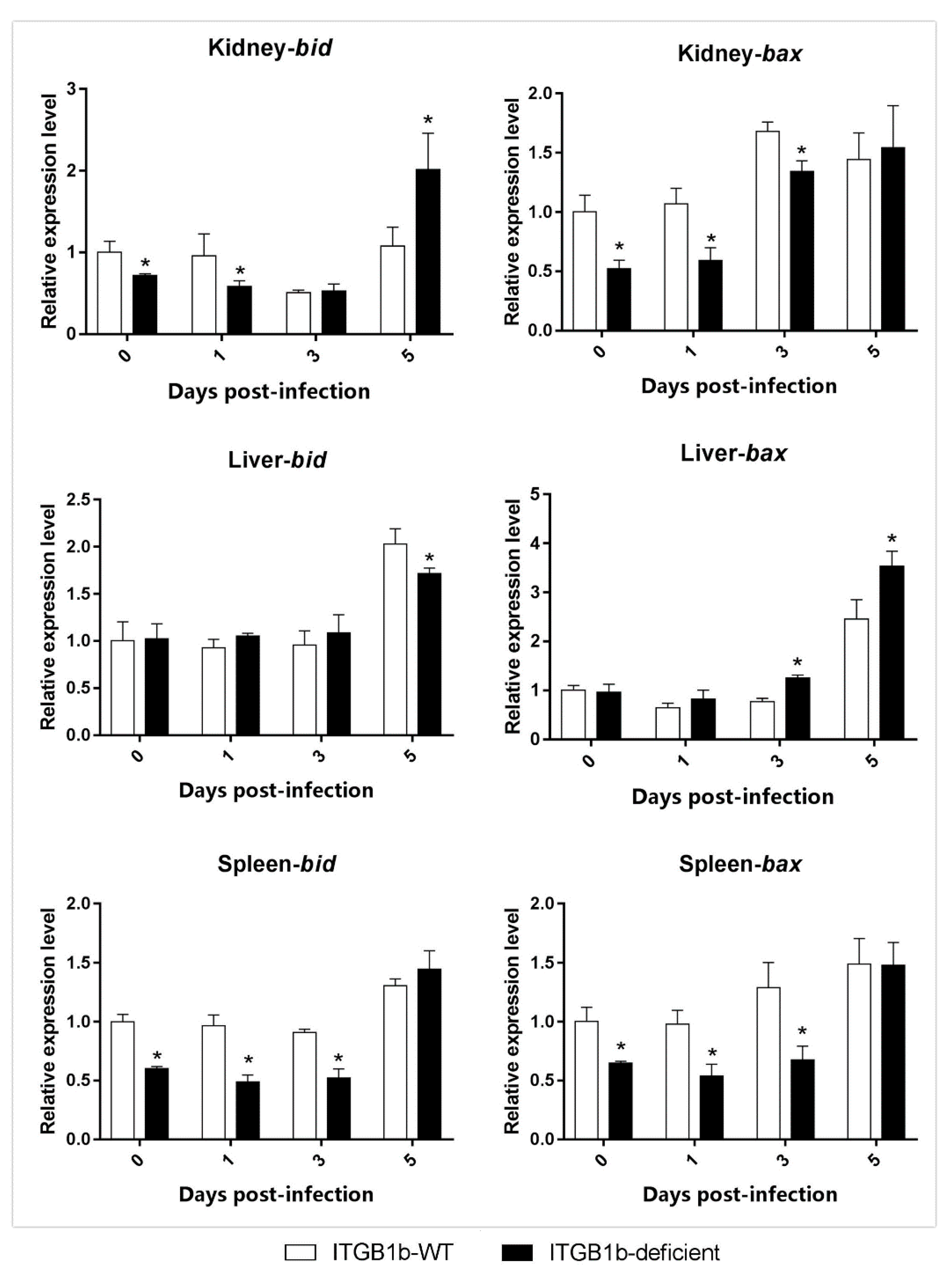
2.6. GCRV-Induced Apoptosis Is Attenuated in ITGB1b-Deficient Rare Minnows
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Experimental Animals and Sample Collection
4.3. Cloning and Sequence Analysis of ITGB1b
4.4. Cas9 Target Site Design and sgRNA Synthesis
4.5. Cas9 mRNA Synthesis
4.6. Microinjection and Identification of Mutations
4.7. Production of ITGB1b-Deficient Rare Minnows
4.8. GCRV Infection
4.9. Detection of the Relative Copy Number of GCRV and Genes
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GCRV | grass carp reovirus |
| ITGB1 | integrin β-1 |
| Dpi | days post-infection |
| dsRNA | double-stranded RNA |
| sgRNA | small guide RNA |
| OCSS | oral squamous cell carcinoma |
| SPC-A-1 | lung cancer cell line |
| WT | wild-type |
| JAM-A | junction adhesion molecule-A |
| AP-2 | adaptor protein complex 2 |
| NPXY | Asn-Pro-X-Tyr |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| BAX | Bcl-2 Associated X Protein |
| BAK | Bcl-2 homologous antagonist killer |
| DR5 | death receptor 5 |
| BHRF1 | BamHI fragment H rightward open reading frame 1 |
| HCV | hepatitis C virus |
| RT-qPCR | real-time quantitative PCR |
| CZRC | China Zebrafish Resource Center |
| TRAIL | TNF-related apoptosis-inducing ligand |
| Ct value | cycle threshold value |
| ANOVA | analysis of Variance |
References
- Food and Agriculture Organization of the United Nations (FAO). Fishery and Aquaculture Statistics Yearbook; FAO: Rome, Italy, 2017. [Google Scholar]
- Wang, Y.; Lu, Y.; Zhang, Y.; Ning, Z.; Li, Y.; Zhao, Q.; Lu, H.; Huang, R.; Xia, X.; Feng, Q.; et al. The draft genome of the grass carp (Ctenopharyngodon idellus) provides genomic insights into its evolution and vegetarian diet adaptation. Nat. Genet. 2015, 47, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Mertens, P.P.; Becnel, J.; Belaganahalli, S.; Bergoin, M.; Brussaard, C.P.; Chappell, J.D.; Ciarlet, M.; del Vas, M.; Dermody, T.S.; et al. Family Reoviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: San Diego, CA, USA, 2012; pp. 538–637. [Google Scholar]
- Chen, G.; He, L.; Luo, L.; Huang, R.; Liao, L.; Li, Y.; Zhu, Z.; Wang, Y. Transcriptomics Sequencing Provides Insights into Understanding the Mechanism of Grass Carp Reovirus Infection. Int. J. Mol. Sci. 2018, 19, 488. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.H.; Chen, H.X.; Liu, P.L.; Liu, H.Q.; Guo, W.; Yi, Y.L. Observations on the ultra-thin sections of the main organs and tissues of hemorrhagic Gobiocypris rarus artificially infected by grass carp hemorrhagic virus (GCHV). Acta Hydrobiol. Sin. 1993, 17, 343–346. (In Chinese) [Google Scholar]
- Liang, X.; Zha, J. Toxicogenomic applications of Chinese rare minnow (Gobiocypris rarus) in aquatic toxicology. Comp. Biochem. Physiol. Part D Genom. Proteom. 2016, 19, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.A.; Brugge, J.S. Integrins and signal transduction pathways: The road taken. Science 1995, 268, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Giancotti, F.G.; Ruoslahti, E. Integrin signaling. Science 1999, 285, 1028–1033. [Google Scholar] [CrossRef] [PubMed]
- Gahmberg, C.G.; Fagerholm, S.C.; Nurmi, S.M.; Chavakis, T.; Marchesan, S.; Grönholm, M. Regulation of integrin activity and signaling. Biochim. Biophys. Acta 2009, 1790, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Summerford, C.; Bartlett, J.S.; Samulski, R.J. αVβ5 integrin: a co-receptor for adeno-associated virus type 2 infection. Nat. Med. 1999, 5, 78. [Google Scholar] [CrossRef] [PubMed]
- Neff, S.; Sá-Carvalho, D.; Rieder, E.; Mason, P.W.; Blystone, S.D.; Brown, E.J.; Baxt, B. Foot-and-mouth disease virus virulent for cattle utilizes the integrin alpha(v)beta3 as its receptor. J. Virol. 1998, 72, 3587–3594. [Google Scholar] [PubMed]
- Gavrilovskaya, I.N.; Brown, E.J.; Ginsberg, M.H.; Mackow, E.R. Cellular entry of hantaviruses which cause hemorrhagic fever with renal syndrome is mediated by beta3 integrins. J. Virol. 1999, 73, 3951–3959. [Google Scholar] [PubMed]
- Wang, X.; Li, L.; Liu, D. Expression analysis of integrin β1 isoforms during zebrafish embryonic development. Gene Expr. Patterns 2014, 16, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Li, J.; Yan, M.X. Integrative analyses identify osteopontin, LAMB3 and ITGB1 as critical pro-metastatic genes for lung cancer. PLoS ONE 2013, 8, e55714. [Google Scholar] [CrossRef] [PubMed]
- Hunt, S.; Jones, A.V.; Hinsley, E.E.; Whawell, S.A.; Lambert, D.W. MicroRNA-124 suppresses oral squamous cell carcinoma motility by targeting ITGB1. FEBS Lett. 2011, 585, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Han, T.S.; Hur, K.; Xu, G.; Choi, B.; Okugawa, Y.; Toiyama, Y.; Chang, A.N. MicroRNA-29c mediates initiation of gastric carcinogenesis by directly targeting ITGB1. Gut 2015, 64, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Keller, M.; Bader, B.L.; Korytář, T.; Finke, S.; Ziegler, U.; Groschup, M.H. Integrins modulate the infection efficiency of West Nile virus into cells. J. Gen. Virol. 2013, 94, 1723–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Wang, H.; Luo, L.; Li, Y.; Huang, R.; Liao, L.; Wang, Y. Bid-deficient fish delay grass carp reovirus (GCRV) replication and attenuate GCRV-triggered apoptosis. Oncotarget 2017, 8, 76408. [Google Scholar] [CrossRef] [PubMed]
- Jackson, T.; Sheppard, D.; Denyer, M.; Blakemore, W.; King, A.M. The epithelial integrin αvβ6 is a receptor for foot-and-mouth disease virus. J. Virol. 2000, 74, 4949–4956. [Google Scholar] [CrossRef] [PubMed]
- Jackson, T.; Mould, A.P.; Sheppard, D.; King, A.M. Integrin αvβ1 is a receptor for foot-and-mouth disease virus. J. Virol. 2002, 76, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Brown, S.L.; Stupack, D.G.; Puente, X.S.; Cheresh, D.A.; Nemerow, G.R. Integrin αvβ1 is an adenovirus coreceptor. J. Virol. 2001, 75, 5405–5409. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Huang, D.Y.; Huong, S.M.; Huang, E.S. Integrin αvβ3 is a coreceptor for human cytomegalovirus. Nat. Med. 2005, 11, 515. [Google Scholar] [CrossRef] [PubMed]
- Akula, S.M.; Pramod, N.P.; Wang, F.Z.; Chandran, B. Integrin α3β1 (CD 49c/29) is a cellular receptor for Kaposi’s sarcoma-associated herpesvirus (KSHV/HHV-8) entry into the target cells. Cell 2002, 108, 407–419. [Google Scholar] [CrossRef]
- Graham, K.L.; Takada, Y.; Coulson, B.S. Rotavirus spike protein VP5* binds α2β1 integrin on the cell surface and competes with virus for cell binding and infectivity. J. Gen. Virol. 2006, 87, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Du, J.; Zhou, J.; Chang, H.; Xie, Q. Integrin activation and viral infection. Virol Sin. 2008, 23, 1–7. [Google Scholar] [CrossRef]
- Maginnis, M.S.; Forrest, J.C.; Kopecky-Bromberg, S.A.; Dickeson, S.K.; Santoro, S.A.; Zutter, M.M.; Nemerow, G.R.; Bergelson, J.M.; Dermody, T.S. β1 Integrin Mediates Internalization of Mammalian Reovirus. J. Virol. 2006, 80, 2760–2770. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.L.; Wang, J.L.; Gao, N.; Chen, Z.T.; Tian, Y.P.; An, J. Up-regulated expression of beta3 integrin induced by dengue virus serotype 2 infection associated with virus entry into human dermal microvascular endothelial cells. Biochem. Biophys. Res. Commun. 2007, 356, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Gianni, T.; Gatta, V.; Campadelli-Fiume, G. αVβ3-integrin routes herpes simplex virus to an entry pathway dependent on cholesterol-rich lipid rafts and dynamin2. Proc. Natl. Acad. Sci. USA 2010, 107, 22260–22265. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Wan, Q.; Shang, X.; Su, J. Large-scale SNP screenings identify markers linked with GCRV resistant traits through transcriptomes of individuals and cell lines in Ctenopharyngodon idella. Sci. Rep. 2017, 7, 1184. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zeng, W.; Yin, L.; Wang, Y.; Liu, C.; Li, Y.; Shi, C. Comparative study on physical–chemical and biological characteristics of grass carp reovirus from different genotypes. J. World Aquac. Soc. 2016, 47, 862–872. [Google Scholar] [CrossRef]
- Zona, L.; Lupberger, J.; Sidahmed-Adrar, N.; Thumann, C.; Harris, H.J.; Barnes, A.; Florentin, J.; Tawar, R.G.; Xiao, F.; Turek, M. HRas Signal Transduction Promotes Hepatitis C Virus Cell Entry by Triggering Assembly of the Host Tetraspanin Receptor Complex. Cell Host Microbe 2013, 13, 302–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, E.S.; Connolly, J.L.; Forrest, J.C.; Chappell, J.D.; Dermody, T.S. Utilization of sialic acid as a coreceptor enhances reovirus attachment by multistep adhesion strengthening. J. Biol. Chem. 2001, 276, 2200–2211. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.A.; Shelling, P.; Wetzel, J.D.; Johnson, E.M.; Wilson, G.A.R.; Forrest, J.C.; Aurrand-Lions, M.; Imhof, B.; Stehle, T.; Dermody, T.S. Junctional adhesion molecule A serves as a receptor for prototype and field-isolate strains of mammalian reovirus. J. Virol. 2005, 79, 7967–7978. [Google Scholar] [CrossRef] [PubMed]
- Boll, W.; Rapoport, I.; Brunner, C.; Modis, Y.; Prehn, S.; Kirchhausen, T. The μ2 subunit of the clathrin adaptor AP-2 binds to FDNPVY and YppØ sorting signals at distinct sites. Traffic 2002, 3, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Fan, Y.; Zhou, Y.; Liu, W.; Jiang, N.; Zhang, J.; Zeng, L. Efficient resistance to grass carp reovirus infection in JAM-A knockout cells using CRISPR/Cas9. Fish Shellfish Immunol. 2018, 76, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Maginnis, M.S.; Mainou, B.A.; Derdowski, A.; Johnson, E.M.; Zent, R.; Dermody, T.S. NPXY motifs in the β1 integrin cytoplasmic tail are required for functional reovirus entry. J. Virol. 2008, 82, 3181–3191. [Google Scholar] [CrossRef] [PubMed]
- Schulz, W.L.; Haj, A.K.; Schiff, L.A. Reovirus Uses Multiple Endocytic Pathways for Cell Entry. J. Virol. 2012, 86, 12665–12675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kominsky, D.J.; Bickel, R.J.; Tyler, K.L. Reovirus-induced apoptosis requires both death receptor- and mitochondrial-mediated caspase-dependent pathways of cell death. Cell Death Differ. 2002, 9, 926–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kominsky, D.J.; Bickel, R.J.; Tyler, K.L. Reovirus-induced apoptosis requires mitochondrial release of Smac/DIABLO and involves reduction of cellular inhibitor of apoptosis protein levels. J. Virol. 2002, 76, 11414–11424. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.; Meintzer, S.M.; Gibson, S.; Widmann, C.; Garrington, T.P.; Johnson, G.L.; Tyler, K.L. Reovirus-induced apoptosis is mediated by TRAIL. J. Virol. 2000, 74, 8135–8139. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Nijhawan, D.; Budhardjo, I.; Srinivasula, S.M.; Ahmad, M.; Alnemri, E.S.; Wang, X. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997, 91, 479–489. [Google Scholar] [CrossRef]
- Jia, R.; Cao, L.-P.; Du, J.-L.; Liu, Y.-J.; Wang, J.-H.; Jeney, G.; Yin, G.-J. Grass carp reovirus induces apoptosis and oxidative stress in grass carp (Ctenopharyngodon idellus) kidney cell line. Virus Res. 2014, 185, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Farhana, L.; Dawson, M.I.; Fontana, J.A. Apoptosis induction by a novel retinoid-related molecule requires nuclear factor-κB activation. Cancer Res. 2005, 65, 4909–4917. [Google Scholar] [CrossRef] [PubMed]
- Teodoro, J.G.; Branton, P.E. Regulation of apoptosis by viral gene products. J. Virol. 1997, 71, 1739–1746. [Google Scholar] [PubMed]
- Polster, B.M.; Pevsner, J.; Hardwick, J.M. Viral Bcl-2 homologs and their role in virus replication and associated diseases. Biochim. Biophys. Acta 2004, 1644, 211–227. [Google Scholar] [CrossRef] [PubMed]
- Hay, S.; Kannourakis, G. A time to kill: Viral manipulation of the cell death program. J. Gen. Virol. 2002, 83, 1547–1564. [Google Scholar] [CrossRef] [PubMed]
- Sander, J.D.; Maeder, M.L.; Reyon, D.; Voytas, D.F.; Joung, J.K.; Dobbs, D. ZiFiT (Zinc Finger Targeter): An updated zinc finger engineering tool. Nucleic Acids Res. 2010, 38, W462–W468. [Google Scholar] [CrossRef] [PubMed]
- Chang, N.; Sun, C.; Gao, L.; Zhu, D.; Xu, X.; Zhu, X.; Xiong, J.W.; Xi, J.J. Genome editing with RNA-guided Cas9 nuclease in zebrafish embryos. Cell Res. 2013, 23, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Talbot, J.C.; Amacher, S.L. A streamlined CRISPR pipeline to reliably generate zebrafish frameshifting alleles. Zebrafish 2014, 11, 583–585. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences (5′to 3′) | Usage |
|---|---|---|
| ITGB1b-F | ATATCAGCTCTACTAGGATTTGTCT | ITGB1B cDNA cloning |
| ITGB1b-R | AATGTTTCTCATATTGGGGGTTCAC | |
| q-ITGB1b-F | GGCTACCTGCTGGTGTGTCC | qRT-PCR for ITGB1B |
| q-ITGB1b-R | TCGTTGAAGCCCAGCGGTTT | |
| gRNA-F | TGTAATACGACTCACTATAGGAGAAGCAGGG AACACTGGTTTTAGAGCTAGAAATAGC | gRNA amplification |
| gRNA-R | AAAAAAAGCACCGACTCGGTGCCAC | |
| T-F | ATGTTCAAGAGGGCAGAGGA | Target site detection |
| T-R | ACTGGAGACTTTGCCCGTAG | |
| S6-F | AGCGCAGCAGGCAATTACTATCT | qRT-PCR for GCRV segment S6 |
| S6-R | ATCTGCTGGTAATGCGGAACG | |
| β-F | TGTAGCCACGCTCGGTCAG | qRT-PCR for β-actin |
| β-R | GGTATCGTGATGGACTCTGGTG | |
| AP2-F | TGCGTTCCGTGTGAACGTGA | qRT-PCR for AP2m1 |
| AP2-R | TGGTAACGGCAGCCAACCAG | |
| Dynamin2-F | GCACCATATCCACCCCCGTG | qRT-PCR for Dynamin-2 |
| Dynamin2-R | TTGGACCTCTCACTGCGGGA | |
| JAMA-F | TGGTGAAGGCGTCACTCAGC | qRT-PCR for JAM-A |
| JAMA-R | TGTCTCCGTGCTCAAGGTGC | |
| Caveolin-F | GCCATGAGAGGCTGCAGGAG | qRT-PCR for Caveolin-1 |
| Caveolin-R | ACAGAGGAAGCCGTGTGGGA | |
| DR5-F | TCCTCTCCGCTCCAGCCATT | qRT-PCR for DR5 |
| DR5-R | AAGAGTTGGCGGTGTTGGGG | |
| Caspase-9F | CGTCCGTCTGGTCATCTATCC | qRT-PCR for Caspase-9 |
| Caspase-9R | GAACTGAGGCAAACCACAATC | |
| Caspase-3F | TCGTAATGGGACAGACAGGG | qRT-PCR for Caspase-3 |
| Caspase-3R | GCCATCGGTGCCATAAATC | |
| Bid-F | ACAGAAACGTCAACGTTCCTCA | qRT-PCR for Bid |
| Bid-R | CTACCTGAGCCTCTACAGCATTGA | |
| Bax-F | TCTGTGCGTCCGCCATCTTC | qRT-PCR for BAX |
| Bax-R | GCTCAGCCTCAGTTACGCCC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, G.; Xiong, L.; Wang, Y.; He, L.; Huang, R.; Liao, L.; Zhu, Z.; Wang, Y. ITGB1b-Deficient Rare Minnows Delay Grass Carp Reovirus (GCRV) Entry and Attenuate GCRV-Triggered Apoptosis. Int. J. Mol. Sci. 2018, 19, 3175. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103175
Chen G, Xiong L, Wang Y, He L, Huang R, Liao L, Zhu Z, Wang Y. ITGB1b-Deficient Rare Minnows Delay Grass Carp Reovirus (GCRV) Entry and Attenuate GCRV-Triggered Apoptosis. International Journal of Molecular Sciences. 2018; 19(10):3175. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103175
Chicago/Turabian StyleChen, Geng, Lv Xiong, Yumeng Wang, Libo He, Rong Huang, Lanjie Liao, Zuoyan Zhu, and Yaping Wang. 2018. "ITGB1b-Deficient Rare Minnows Delay Grass Carp Reovirus (GCRV) Entry and Attenuate GCRV-Triggered Apoptosis" International Journal of Molecular Sciences 19, no. 10: 3175. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103175