Study on Cardiotoxicity and Mechanism of “Fuzi” Extracts Based on Metabonomics

 ,
,
Abstract
:1. Introduction
2. Results
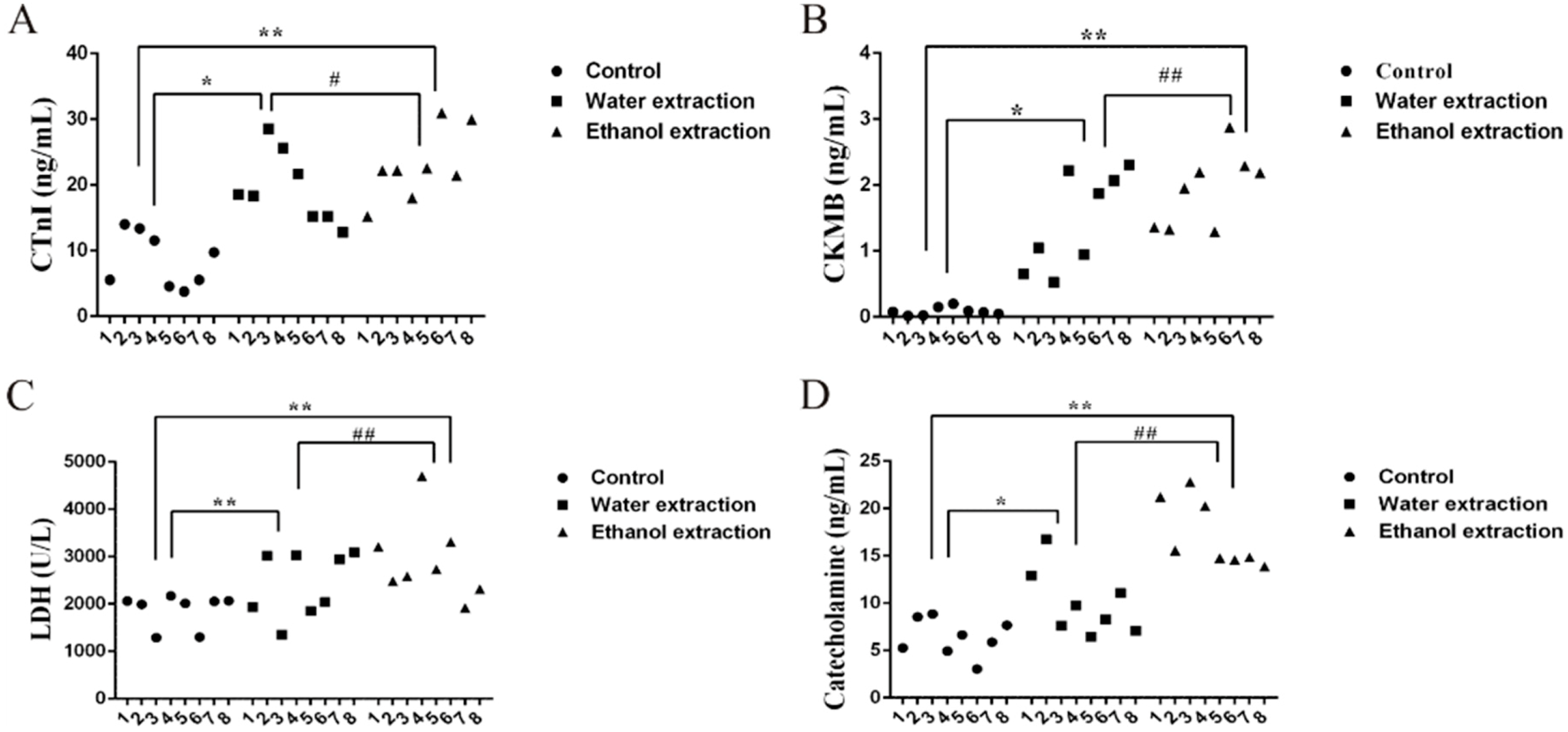
2.1. Cardiotoxicity of Water and Ethanol Extracts of FZ
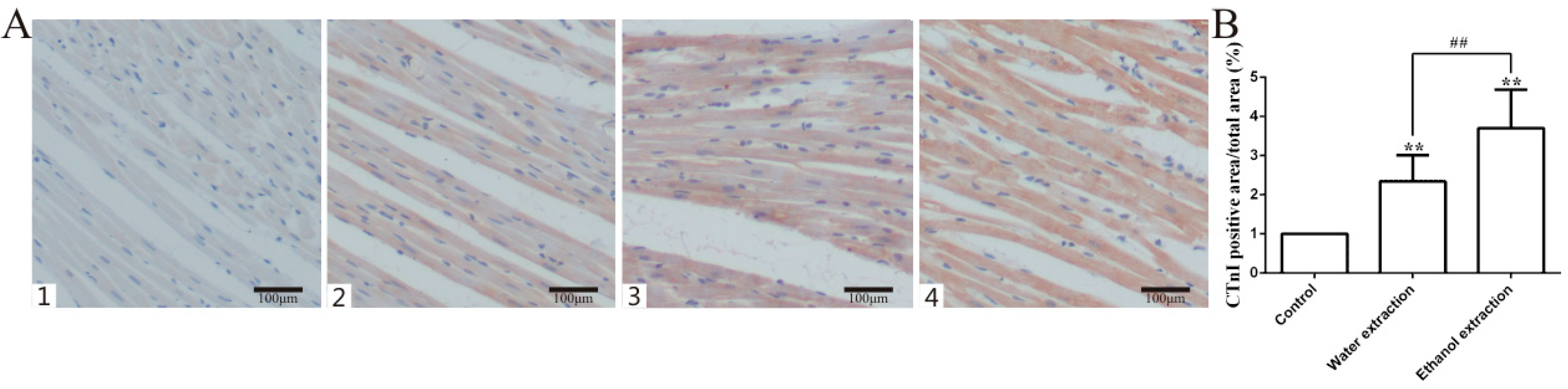
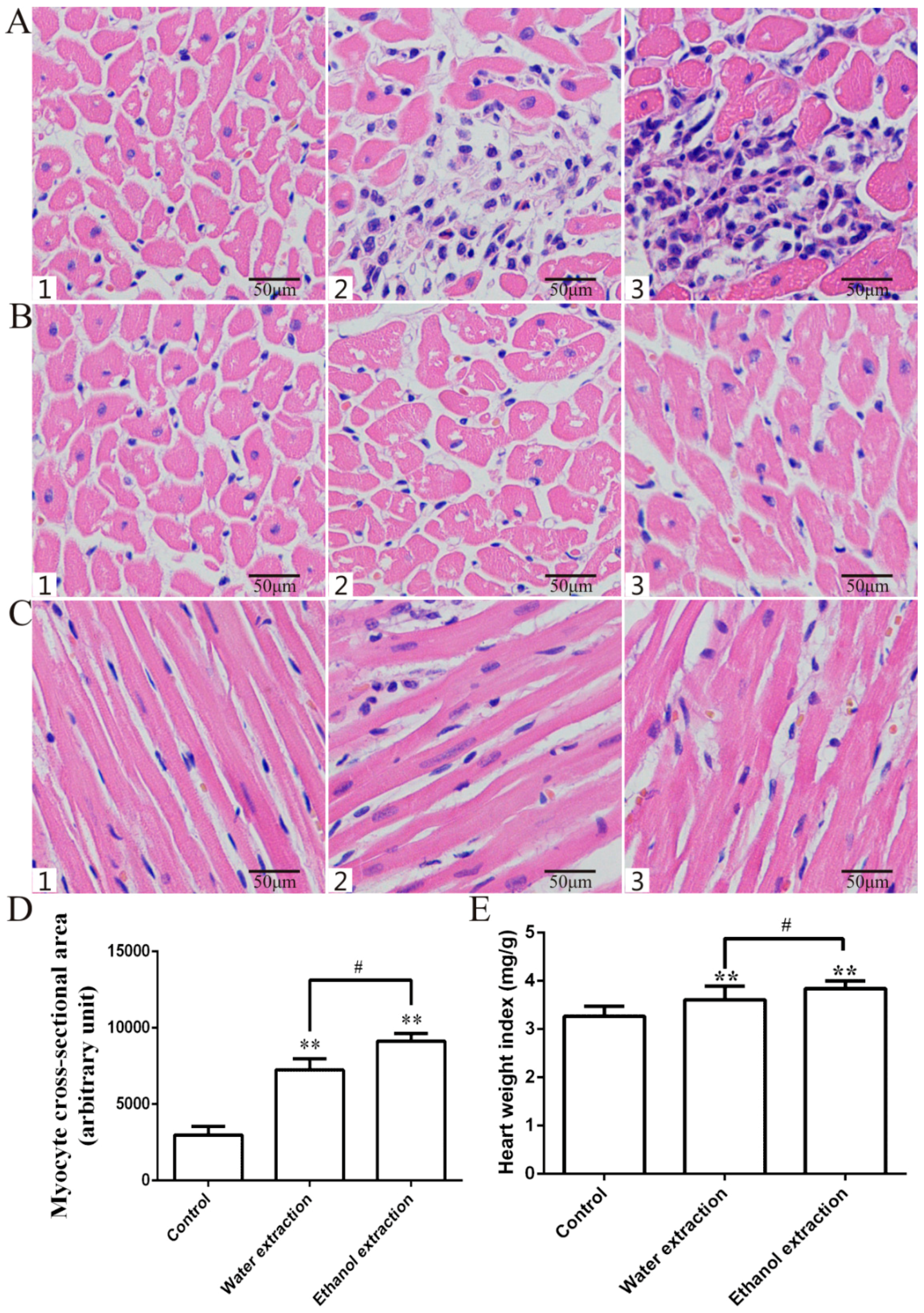
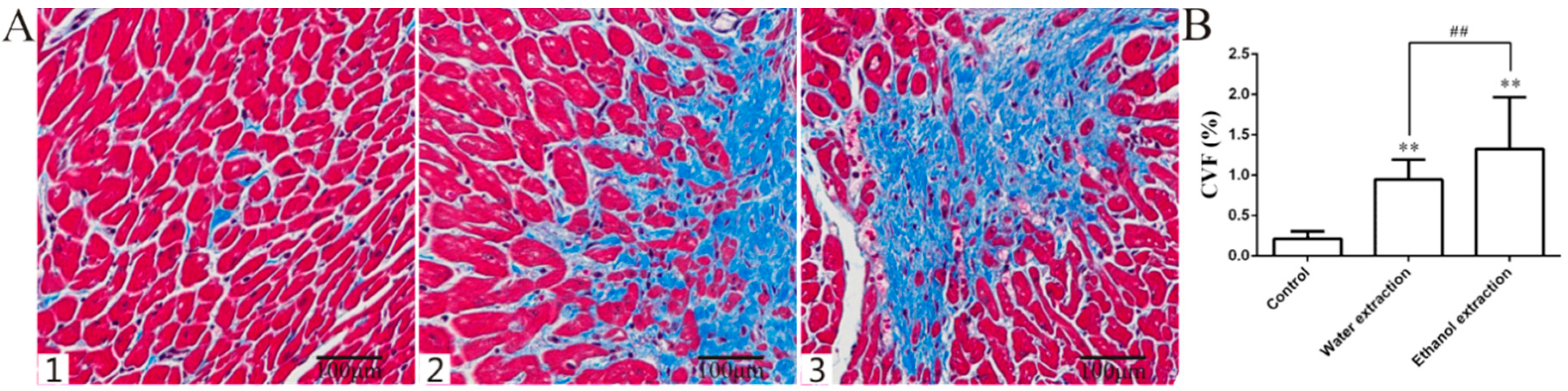
2.2. Pathology Results
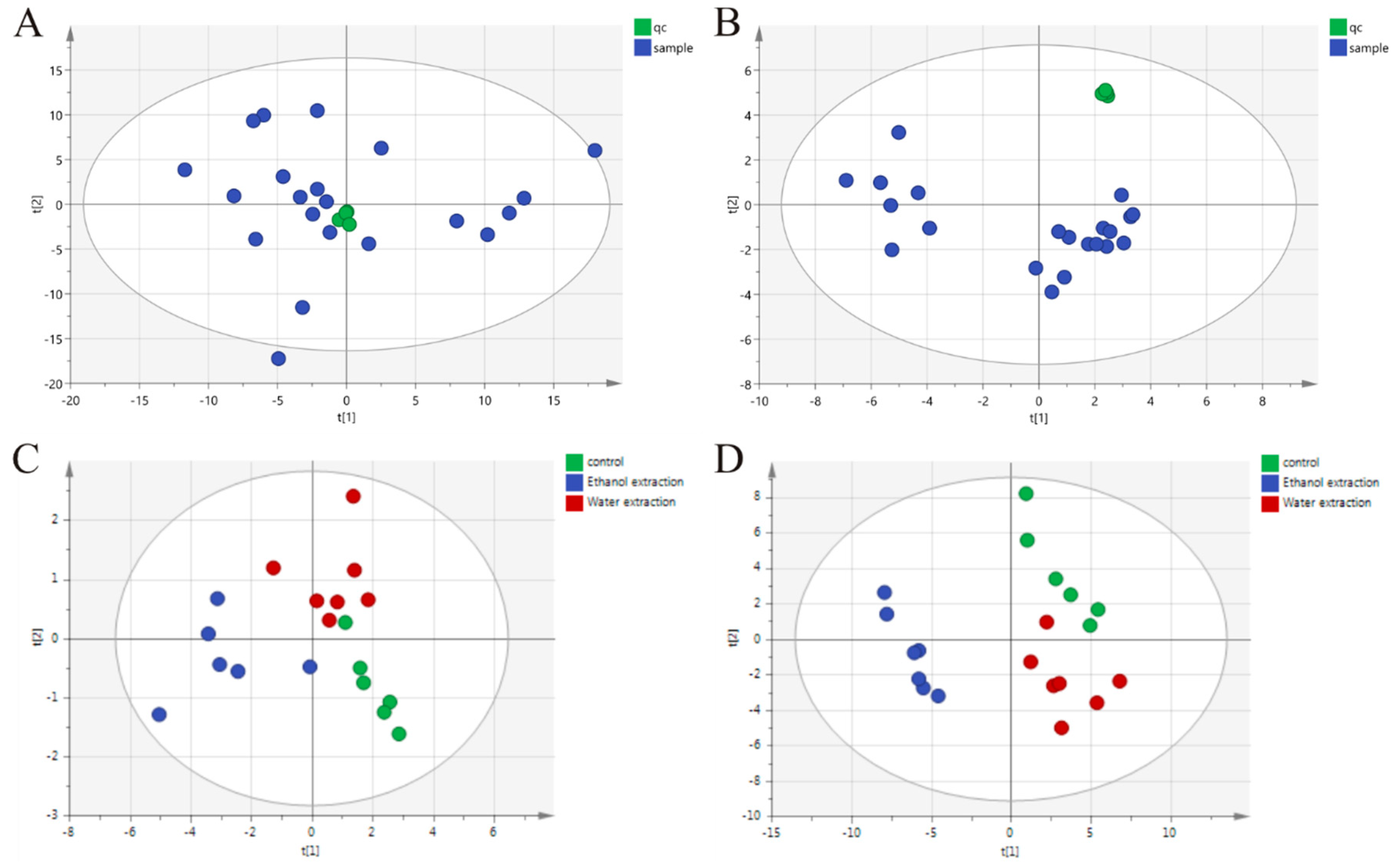
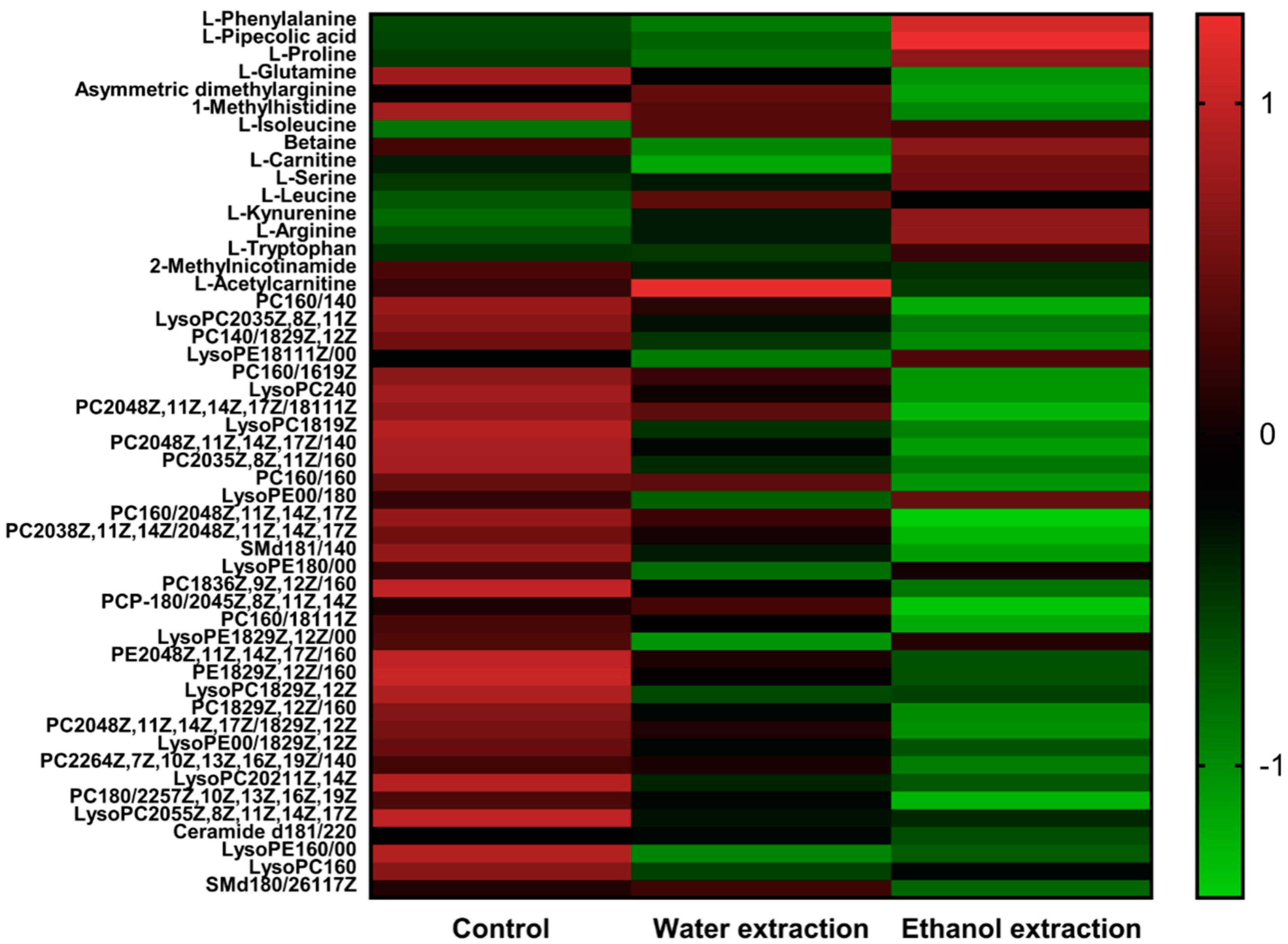
2.3. Identification of Differential Metabolites
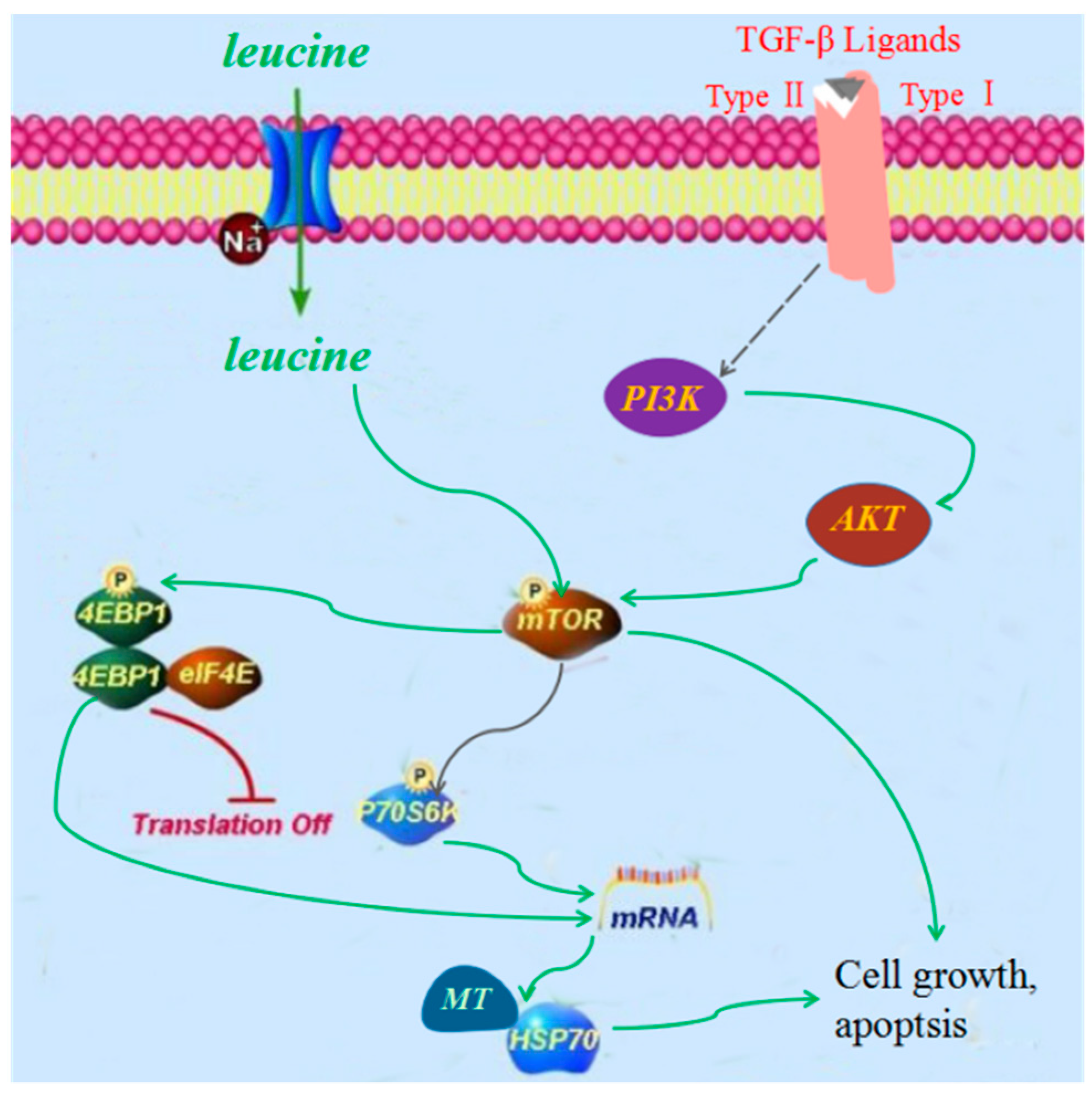
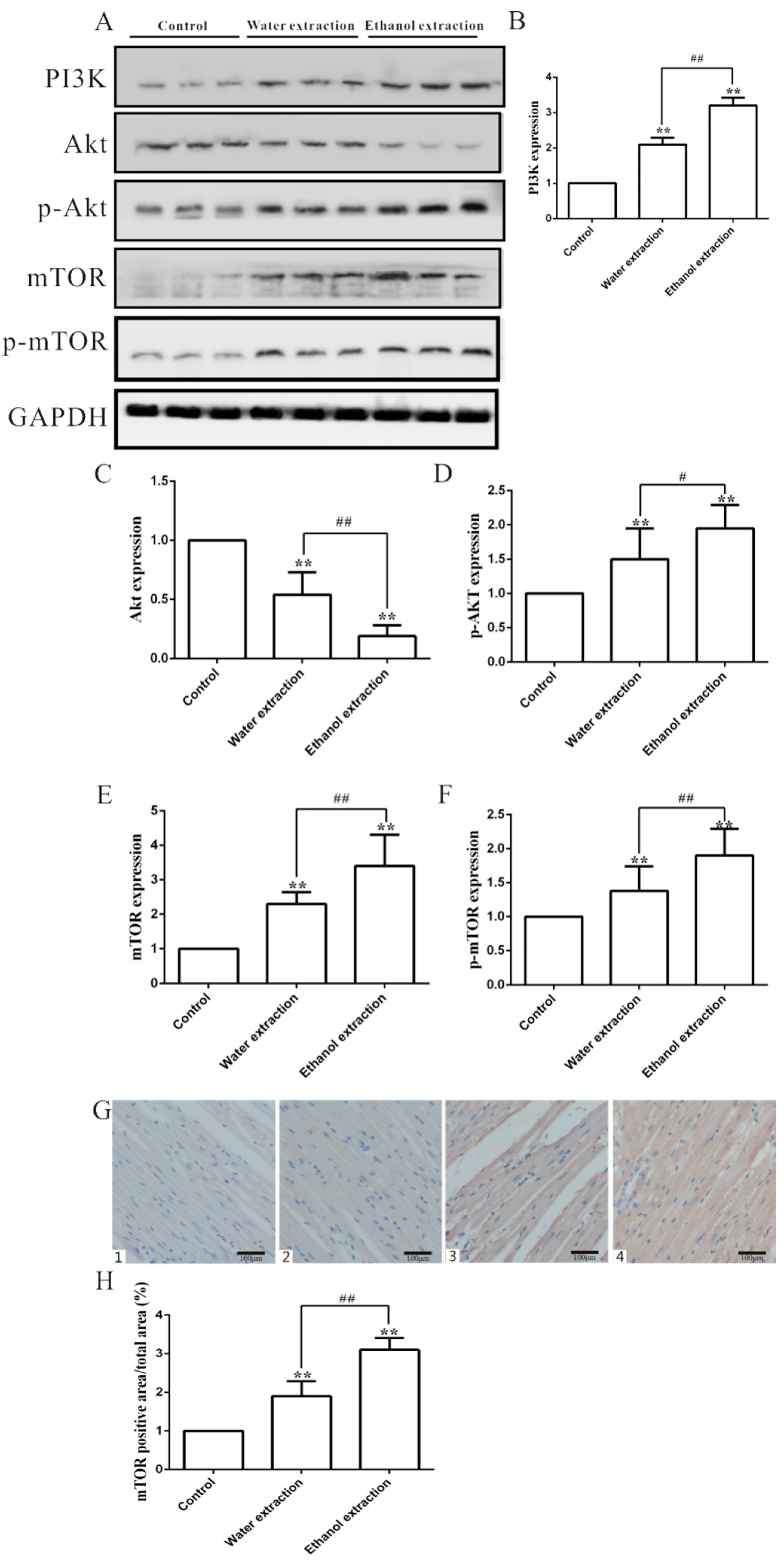
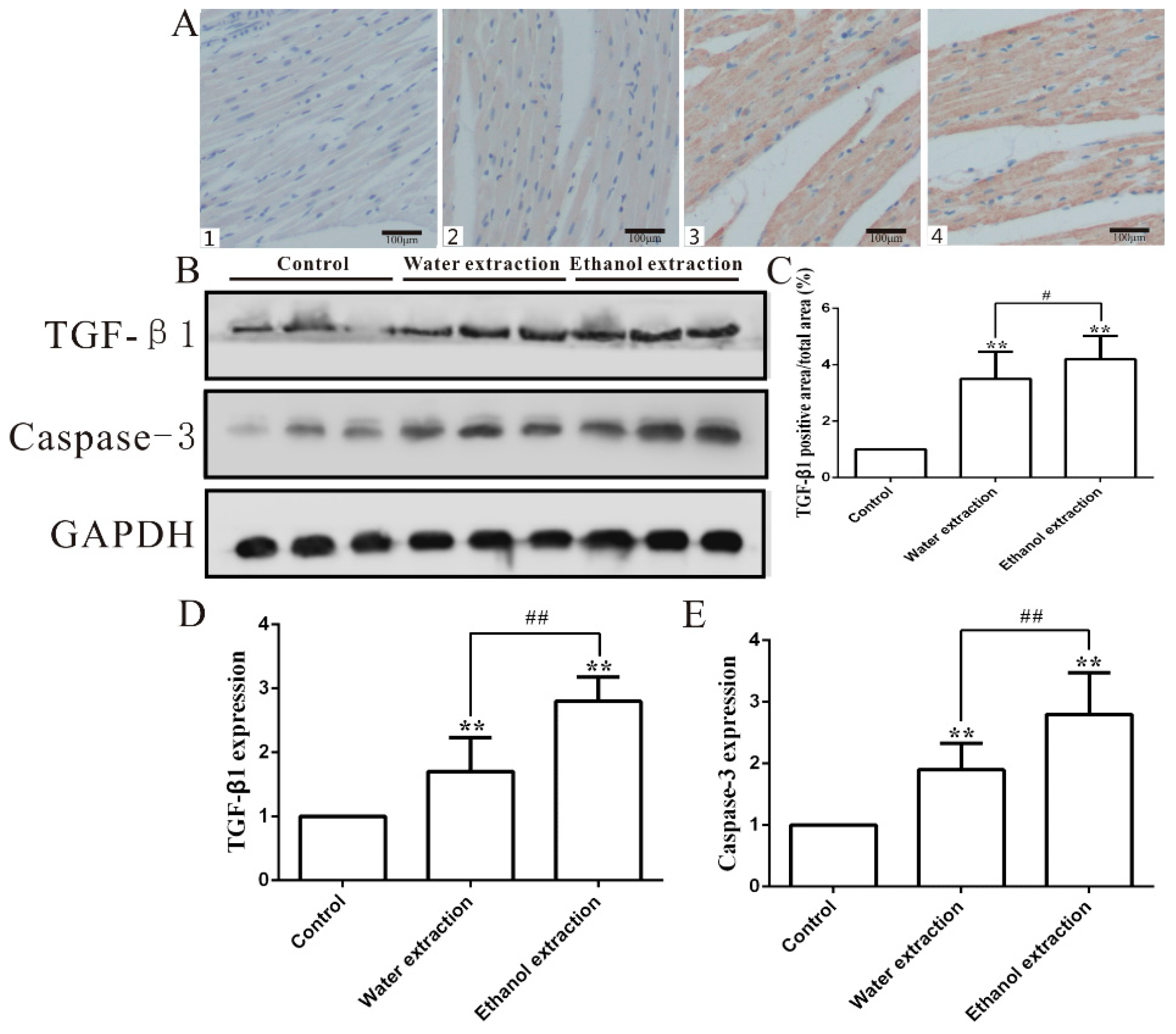
2.4. Cardiotoxicity of FZ Extracts in Rats via the PI3K/Akt/mTOR Signaling Pathway
2.5. Water and Ethanol Extracts of FZ Induce Cardiomyocyte Apoptosis in Rats
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Materials
4.3. Water and Ethanol Extracts of FZ
4.3.1. Water Extracts of FZ
4.3.2. Ethanol Extracts of FZ
4.4. Grouping and Experimental Design
4.5. Serum Sample Preparation
4.5.1. Analysis of Serum Samples by Reversed-Phase Chromatography
4.5.2. Method of Sample Treatment for Hydrophilic Chromatographic Analysis of Serum
4.6. Chromatography
4.6.1. Reversed-Phase Chromatographic Separation
4.6.2. Hydrophilic Chromatographic Separation
4.6.3. QC
4.7. Mass Spectrometry
4.8. Histological and Morphological Analyses of the Heart
4.9. Measurement of Serum CTnI and CK-MB Activity Levels in Serum and Cardiac Tissue
4.10. Measurement of Myocardial Enzymes
4.11. Western Blotting of PI3K, Akt, mTOR, TGF-β1, and Caspase-3
4.12. Immunohistochemistry of CTnI, mTOR, and TGF-β1
4.13. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
Ethics Approval
References
- Li, X.M. Complementary and alternative medicine in pediatric allergic disorders. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 161–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Penchala, S.; Prabhu, S.; Wang, J.; Huang, Y. Molecular basis of traditional chinese medicine in cancer chemoprevention. Curr. Drug Discov. Technol. 2010, 7, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Singhuber, J.; Zhu, M.; Prinz, S.; Kopp, B. Aconitum in traditional chinese medicine: A valuable drug or an unpredictable risk? J. Ethnopharmacol. 2009, 126, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, R.; Motoya, W.; Atsumi, T.; Mouri, C.; Kakiuchi, N.; Mikage, M. The relationship between growth of the aerial part and alkaloid content variation in cultivated aconitum carmichaeli debeaux. J. Nat. Med. 2011, 65, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.G.; Wang, F.P.; Gao, F.; Yan, L.P.; Chen, D.L.; Liu, Y. A pharmacophylogenetic study of aconitum l. (ranunculaceae) from china. Acta Phytotax. Sin. 2006, 44, 1–46. [Google Scholar] [CrossRef]
- Shaw, D. Toxicological risks of Chinese herbs. Planta Med. 2010, 76, 2012–2018. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Dong, Z.; Wang, Q.; Qian, G.; Huang, W.; Jiang, Z.; Leung, K.S.; Zhao, Z. Toxicity assessment of nine types of decoction pieces from the daughter root of aconitum carmichaeli (fuzi) based on the chemical analysis of their diester diterpenoid alkaloids. Planta Med. 2010, 76, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.Y.; Tomlinson, B.; Tse, L.K.; Chan, J.C.; Chan, W.W.; Critchley, J.A. Aconitine poisoning due to chinese herbal medicines: A. review. Vet. Hum. Toxicol. 1994, 36, 452–455. [Google Scholar] [PubMed]
- Sun, B.; Li, L.; Wu, S.; Zhang, Q.; Li, H.; Chen, H.; Li, F.; Dong, F.; Yan, X. Metabolomic analysis of biofluids from rats treated with aconitum alkaloids using nuclear magnetic resonance and gas chromatography/time-of-flight mass spectrometry. Anal. Biochem. 2009, 395, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Li, Y.Y.; Wang, D.; Huang, X.; Liu, Q. Diterpenoid alkaloids from the chinese traditional herbal ′fuzi′ and their cytotoxic activity. Molecules 2012, 17, 5187–5194. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Cai, H.; Zhang, Y.; Cong, X.; Zhang, C.; Cai, B. Identification of metabolites of crude and processed fructus corni in rats by microdialysis sampling coupled with electrospray ionization linear quadrupole ion trap mass spectrometry. J. Pharm. Biomed. Anal. 2011, 56, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Ni, B.; Zhang, A.; Wang, M.; Dong, H.; Wang, X. Metabolomics study on fuzi and its processed products using ultra-performance liquid-chromatography/electrospray-ionization synapt high-definition mass spectrometry coupled with pattern recognition analysis. Analyst 2012, 137, 170–185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Zhu, Y.; Fan, X.H.; Zhang, B.L. Efficacy-oriented compatibility for component-based chinese medicine. Acta Pharmacol. Sin. 2015, 36, 654–658. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, H.; Xie, J.; Ally, M.S.; Hou, Z.; Xu, Y.; Zhang, Y. A novel method for evaluating the cardiotoxicity of traditional chinese medicine compatibility by using support vector machine model combined with metabonomics. Evid. Based Complement. Altern. Med. 2016, 2016, 6012761. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Hao, J.; Jones, S.C.; Yee, M.S.; Roth, J.C.; Dixon, I.M. Decreased smad 7 expression contributes to cardiac fibrosis in the infarcted rat heart. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, H1685–H1696. [Google Scholar] [CrossRef] [PubMed]
- Brugge, J.; Hung, M.C.; Mills, G.B. A new mutational aktivation in the pi3k pathway. Cancer Cell 2007, 12, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Shyur, L.F.; Yang, N.S. Metabolomics for phytomedicine research and drug development. Curr. Opin. Chem. Biol. 2008, 12, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.H.; Sun, H.; Han, Y.; Yan, G.L.; Yuan, Y.; Song, G.C.; Yuan, X.X.; Xie, N.; Wang, X.J. Ultraperformance liquid chromatography-mass spectrometry based comprehensive metabolomics combined with pattern recognition and network analysis methods for characterization of metabolites and metabolic pathways from biological data sets. Anal. Chem. 2013, 85, 7606–7612. [Google Scholar] [CrossRef] [PubMed]
- Kawano, H.; Do, Y.S.; Kawano, Y.; Starnes, V.; Barr, M.; Law, R.E.; Hsueh, W.A. Angiotensin ii has multiple profibrotic effects in human cardiac fibroblasts. Circulation 2000, 101, 1130–1137. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Siddiqui, M.A. Signaling networks regulating cardiac myocyte survival and death. Curr. Opin. Investig. Drugs 2009, 10, 928–937. [Google Scholar] [PubMed]
- Wu, J.; Sun, C.; Wang, R.; Li, J.; Zhou, M.; Yan, M.; Xue, X.; Wang, C. Cardioprotective effect of paeonol against epirubicin-induced heart injury via regulating mir-1 and pi3k/akt pathway. Chem.-Biol. Interact. 2018, 286, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Hu, Z.; Li, B.; Wang, Z.; Chen, S. Ivabradine improved left ventricular function and pressure overload-induced cardiomyocyte apoptosis in a transverse aortic constriction mouse model. Mol. Cell. Biochem. 2018, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.H.; Wang, X.T.; Sun, Y.H.; He, W.K.; Liang, J.B.; Mo, B.H.; Li, L. Pim1 overexpression prevents apoptosis in cardiomyocytes after exposure to hypoxia and oxidative stress via upregulating cell autophagy. Cell. Physiol. Biochem. 2018, 49, 2138–2150. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, M.S.; Fukunaga, K. Inhibition of htra2/omi ameliorates heart dysfunction following ischemia/reperfusion injury in rat heart in vivo. Eur. J. Pharmacol. 2007, 557, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Kossmehl, P.; Kurth, E.; Faramarzi, S.; Habighorst, B.; Shakibaei, M.; Wehland, M.; Kreutz, R.; Infanger, M.; Danser, A.H.J.; Grosse, J.; et al. Mechanisms of apoptosis after ischemia and reperfusion: Role of the renin-angiotensin system. Apoptosis 2006, 11, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Hers, I.; Vincent, E.E.; Tavare, J.M. Akt signalling in health and disease. Cell. Signal. 2011, 23, 1515–1527. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.E.; Dupont, C.L.; Obornik, M.; Horak, A.; Nunes-Nesi, A.; McCrow, J.P.; Zheng, H.; Johnson, D.A.; Hu, H.; Fernie, A.R.; et al. Evolution and metabolic significance of the urea cycle in photosynthetic diatoms. Nature 2011, 473, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, A.; Poisson, L.M.; Rajendiran, T.M.; Khan, A.P.; Cao, Q.; Yu, J.; Laxman, B.; Mehra, R.; Lonigro, R.J.; Li, Y.; et al. Metabolomic profiles delineate potential role for sarcosine in prostate cancer progression. Nature 2009, 457, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Yu, Q.; Yan, X.; Hang, W.; Zheng, J.; Xing, J.; Huang, B. Direct infusion mass spectrometry or liquid chromatography mass spectrometry for human metabonomics? A serum metabonomic study of kidney cancer. Analyst 2010, 135, 2970–2978. [Google Scholar] [CrossRef] [PubMed]
- Junot, C.; Madalinski, G.; Tabet, J.C.; Ezan, E. Fourier transform mass spectrometry for metabolome analysis. Analyst 2010, 135, 2203–2219. [Google Scholar] [CrossRef] [PubMed]
- Zoncu, R.; Efeyan, A.; Sabatini, D.M. Mtor: From growth signal integration to cancer, diabetes and ageing. Nat. Rev. Mol. Cell Biol. 2011, 12, 21–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manickam, V.; Periyasamy, M.; Dhakshinamoorthy, V.; Panneerselvam, L.; Perumal, E. Recurrent exposure to ferric oxide nanoparticles alters myocardial oxidative stress, apoptosis and necrotic markers in male mice. Chem.-Biol. Interact. 2017, 278, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Wever, E.F.; Robles de Medina, E.O. Sudden death in patients without structural heart disease. J. Am. Coll. Cardiol. 2004, 43, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, G.J. Therapeutic strategies of drug repositioning targeting autophagy to induce cancer cell death: From pathophysiology to treatment. J. Hematol. Oncol. 2017, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, S.; Hosseini, M.; Shahidsales, S.; Maftouh, M.; Ferns, G.A.; Ghayour-Mobarhan, M.; Hassanian, S.M.; Avan, A. Targeting the akt/pi3k signaling pathway as a potential therapeutic strategy for the treatment of pancreatic cancer. Curr. Med. Chem. 2017, 24, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Nebigil, C.G.; Maroteaux, L. Functional consequence of serotonin/5-ht2b receptor signaling in heart: Role of mitochondria in transition between hypertrophy and heart failure? Circulation 2003, 108, 902–908. [Google Scholar] [CrossRef] [PubMed]
- Jian, B.; Xu, J.; Connolly, J.; Savani, R.C.; Narula, N.; Liang, B.; Levy, R.J. Serotonin mechanisms in heart valve disease i: Serotonin-induced up-regulation of transforming growth factor-beta1 via g-protein signal transduction in aortic valve interstitial cells. Am. J. Pathol. 2002, 161, 2111–2121. [Google Scholar] [CrossRef]
- Jian, B.; Narula, N.; Li, Q.Y.; Mohler, E.R., 3rd; Levy, R.J. Progression of aortic valve stenosis: Tgf-beta1 is present in calcified aortic valve cusps and promotes aortic valve interstitial cell calcification via apoptosis. Ann. Thorac. Surg. 2003, 75, 457–465, discussion 465–466. [Google Scholar] [CrossRef]
- Razavi, B.M.; Hosseinzadeh, H.; Abnous, K.; Khoei, A.; Imenshahidi, M. Protective effect of crocin against apoptosis induced by subchronic exposure of the rat vascular system to diazinon. Toxicol. Ind. Health 2016, 32, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Wu, M.; Wang, J.F.; Qiao, Y.J.; Wang, Z. Disruption of the intracellular ca2+ homeostasis in the cardiac excitation-contraction coupling is a crucial mechanism of arrhythmic toxicity in aconitine-induced cardiomyocytes. Biochem. Biophys. Res. Commun. 2007, 354, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, P.; Huang, Y.; Lu, Z.; Yang, Z.; Xu, L.; Sun, F.; Zhang, Y.; Duan, J. Camp-pka-camkii signaling pathway is involved in aggravated cardiotoxicity during fuzi and beimu combination treatment of experimental pulmonary hypertension. Sci. Rep. 2016, 6, 34903. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Yang, L.; Zhang, Z.; Ren, S.; Tang, X.; Zhou, W.; Wang, Y.; Ma, Z.; Gao, S.; Gao, Y. Human pxr-mediated transcriptional activation of cyp3a4 by ‘fuzi’ extracts. Toxicol. Mech. Meth. 2018, 1–19. [Google Scholar] [CrossRef] [PubMed]
- He, J.L.; Zhao, J.W.; Ma, Z.C.; Liang, Q.D.; Wang, Y.G.; Tan, H.L.; Xiao, C.R.; Tang, X.-L.; Gao, Y. [cardiotoxicity study of shenfu compatibility in rats based on metabonomics]. Zhongguo Zhong Yao Za Zhi 2015, 40, 2743–2747. [Google Scholar] [PubMed]
- Su, T.; Yu, H.; Kwan, H.Y.; Ma, X.Q.; Cao, H.H.; Cheng, C.Y.; Leung, A.K.; Chan, C.L.; Li, W.D.; Cao, H.; et al. Comparisons of the chemical profiles, cytotoxicities and anti-inflammatory effects of raw and rice wine-processed herba siegesbeckiae. J. Ethnopharmacol. 2014, 156, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Su, T.; Cheng, B.C.; Fu, X.Q.; Li, T.; Guo, H.; Cao, H.H.; Kwan, H.Y.; Tse, A.K.; Yu, H.; Cao, H.; et al. Comparison of the toxicities, activities and chemical profiles of raw and processed xanthii fructus. BMC Complement. Altern. Med. 2016, 16, 24. [Google Scholar]
- Sanchez Canedo, C.; Demeulder, B.; Ginion, A.; Bayascas, J.R.; Balligand, J.L.; Alessi, D.R.; Vanoverschelde, J.L.; Beauloye, C.; Hue, L.; Bertrand, L. Activation of the cardiac mtor/p70(s6k) pathway by leucine requires pdk1 and correlates with pras40 phosphorylation. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E761–E769. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yin, Y.; Tan, B.; Kong, X.; Wu, G. Leucine nutrition in animals and humans: Mtor signaling and beyond. Amino Acids 2011, 41, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, S.; Shibata, R.; Ohashi, K.; Ohashi, T.; Daida, H.; Walsh, K.; Murohara, T.; Ouchi, N. Adiponectin ameliorates doxorubicin-induced cardiotoxicity through akt protein-dependent mechanism. J. Biol. Chem. 2011, 286, 32790–32800. [Google Scholar] [CrossRef] [PubMed]
- Sciarretta, S.; Volpe, M.; Sadoshima, J. Mammalian target of rapamycin signaling in cardiac physiology and disease. Circ. Res. 2014, 114, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Barry, S.P.; Townsend, P.A. What causes a broken heart--molecular insights into heart failure. Int. Rev. Cell Mol. Biol. 2010, 284, 113–179. [Google Scholar] [PubMed]
- Zhou, Y.; He, X.; Chen, Y.; Huang, Y.; Wu, L.; He, J. Exendin-4 attenuates cardiac hypertrophy via ampk/mtor signaling pathway activation. Biochem. Biophys. Res. Commun. 2015, 468, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Soonpaa, M.H.; Chen, H.; Shen, W.; Payne, R.M.; Liechty, E.A.; Caldwell, R.L.; Shou, W.; Field, L.J. Acute doxorubicin cardiotoxicity is associated with p53-induced inhibition of the mammalian target of rapamycin pathway. Circulation 2009, 119, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Horbelt, D.; Denkis, A.; Knaus, P. A portrait of transforming growth factor beta superfamily signalling: Background matters. Int. J. Biochem. Cell Biol. 2012, 44, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Ye, D.; Wang, Y. Caspase-3 as a therapeutic target for heart failure. Expert Opin. Ther. Targets 2013, 17, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Akhurst, R.J.; Hata, A. Targeting the tgfbeta signalling pathway in disease. Nat. Rev. Drug Discov. 2012, 11, 790–811. [Google Scholar] [CrossRef] [PubMed]
- Carroll, B.; Korolchuk, V.I.; Sarkar, S. Amino acids and autophagy: Cross-talk and co-operation to control cellular homeostasis. Amino Acids 2015, 47, 2065–2088. [Google Scholar] [CrossRef] [PubMed]
- Draoui, N.; de Zeeuw, P.; Carmeliet, P. Angiogenesis revisited from a metabolic perspective: Role and therapeutic implications of endothelial cell metabolism. Open Biol. 2017, 7, 170219. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.S.; Oh, J.; Kang, S.K.; Park, S.; Lee, S.H.; Choi, D.; Chung, J.H.; Chung, Y.W.; Kang, S.M. Insulin protects cardiac myocytes from doxorubicin toxicity by sp1-mediated transactivation of survivin. PLoS ONE 2015, 10, e0135438. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-W.; He, J.-L.; Ma, Z.-C.; Liang, Q.-D.; Wang, Y.-G.; Tan, H.-L.; Xiao, C.-R.; Tang, X.-L.; Gao, Y. Mitochondrial toxicity effect of radix aconiti lateralis praeparata on h9c2 cardiomyocytes. Chin. J. Pharmacol. Toxicol. 2015, 29, 816–824. [Google Scholar]
- Wang, Y.-X.; Mi, H.-J.; Zhang, C.-L.; Hu, Y.; Shi, J.; Bi, Y.-A.; Wang, Z.-Z.; Xiao, W. Optimization on alcohol precipitation process for lonicerae flos and artemisiae annuae herba in reduning injection by central composite design and response surface methodology. Chin. Tradit. Herb. Drugs 2015, 46, 671–678. [Google Scholar]
- Song, E.-F.; Sun, W.-L.; Zhang, J.; Mei, S.-S. Clinical applications and safety evaluation study of aconite. China J. Tradit. Chin. Med. Pharm. 2013, 28, 895–901. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | tR (min) | Extract Mass | Formula | Compound | Fold Change | |
|---|---|---|---|---|---|---|
| Water Extract vs. Control | Alcohol Extract vs. Control | |||||
| 1 | 3.2376 | 204.1224 | C9H17NO4 | Acetylcarnitine | ↓/0.87 | ↑/1.02 |
| 2 | 4.7173 | 162.1120 | C7H15NO3 | Carnitine | ↑/1.17 | — |
| 3 | 4.75298 | 132.1016 | C6H13NO2 | Isoleucine | ↑/1.33 | ↑/1.35 |
| 4 | 4.82875 | 205.0966 | C11H12N2O2 | Tryptophan | ↑/1.11 | ↑/1.14 |
| 5 | 4.85995 | 209.0915 | C10H12N2O3 | Kynurenine | ↑/1.18 | ↑/1.11 |
| 6 | 4.93126 | 132.1016 | C6H13NO2 | Leucine | ↑/1.21 | ↑/1.39 |
| 7 | 4.9669 | 118.0861 | C5H11NO2 | Betaine | ↓/0.96 | ↓/0.86 |
| 8 | 6.4243 | 106.0500 | C3H7NO3 | Serine | ↑/1.126 | ↑/1.01 |
| 9 | 6.7497 | 203.1498 | C8H18N4O2 | Asymmetric dimethylarginine | ↓/0.60 | — |
| 10 | 7.2177 | 170.0920 | C7H11N3O2 | Methylhistidine | ↓/0.67 | ↓/0.97 |
| 11 | 7.24445 | 175.1185 | C6H14N4O2 | Arginine | ↑/1.39 | ↑/1.12 |
| Time (min) | A (v%) | B (v%) |
|---|---|---|
| 0 | 80 | 20 |
| 2 | 70 | 30 |
| 5 | 55 | 45 |
| 6.5 | 40 | 60 |
| 12 | 35 | 65 |
| 14 | 15 | 85 |
| 17.5 | 0 | 100 |
| 18 | 0 | 100 |
| 18.1 | 80 | 20 |
| 19.5 | 80 | 20 |
| Time (min) | A (v%) | B (v%) |
|---|---|---|
| 0 | 95 | 5 |
| 1 | 95 | 5 |
| 7 | 50 | 50 |
| 9 | 50 | 50 |
| 9.1 | 95 | 5 |
| 13 | 95 | 5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, G.; Yang, L.; Zhou, W.; Tang, X.; Wang, Y.; Ma, Z.; Gao, S.; Gao, Y. Study on Cardiotoxicity and Mechanism of “Fuzi” Extracts Based on Metabonomics. Int. J. Mol. Sci. 2018, 19, 3506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113506
Huang G, Yang L, Zhou W, Tang X, Wang Y, Ma Z, Gao S, Gao Y. Study on Cardiotoxicity and Mechanism of “Fuzi” Extracts Based on Metabonomics. International Journal of Molecular Sciences. 2018; 19(11):3506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113506
Chicago/Turabian StyleHuang, Guangyao, Liang Yang, Wei Zhou, Xianglin Tang, Yuguang Wang, Zengchun Ma, Shan Gao, and Yue Gao. 2018. "Study on Cardiotoxicity and Mechanism of “Fuzi” Extracts Based on Metabonomics" International Journal of Molecular Sciences 19, no. 11: 3506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113506