Identification and Characterization of the Entamoeba Histolytica Rab8a Binding Protein: A Cdc50 Homolog


Abstract
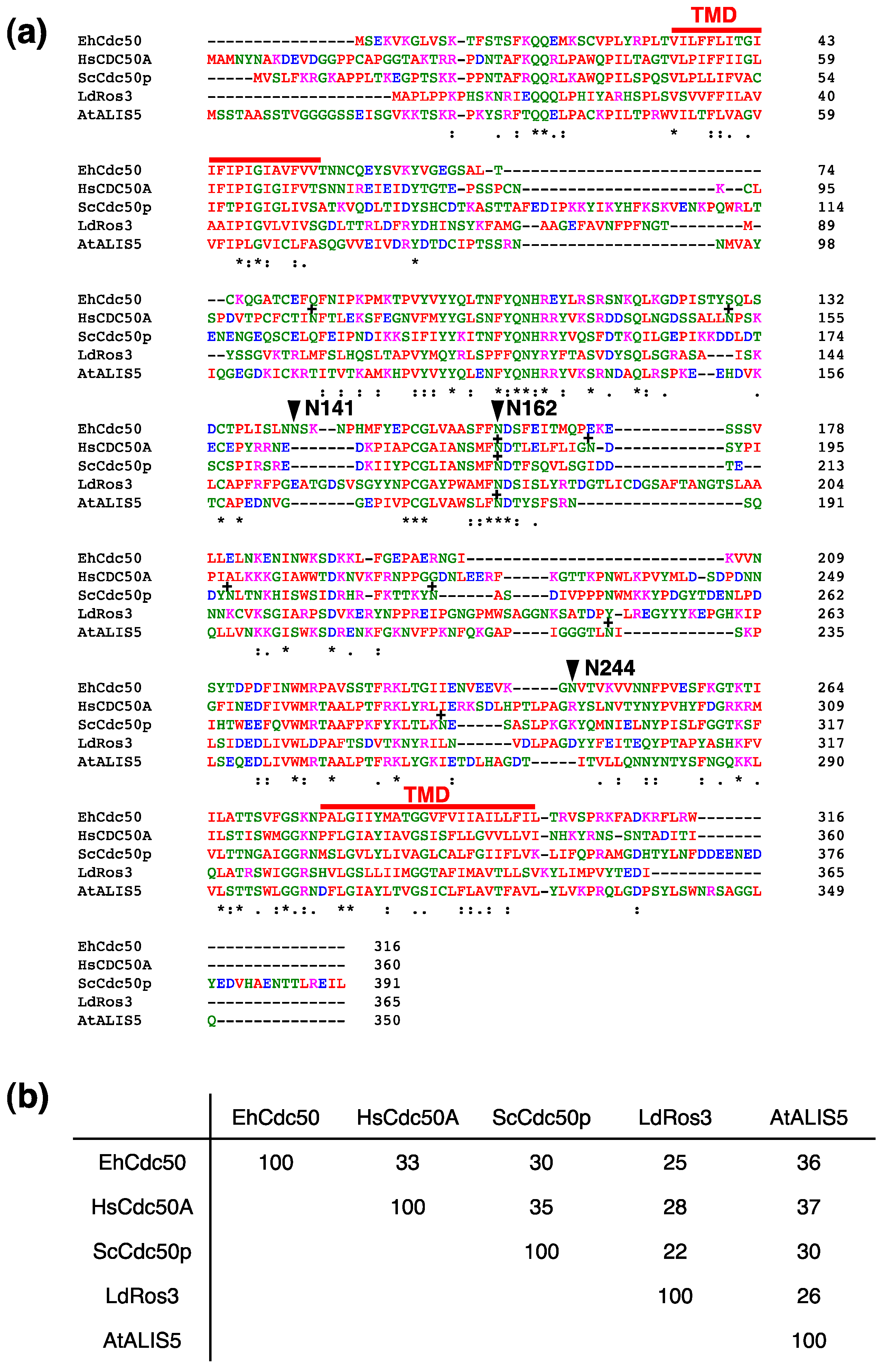
:1. Introduction
2. Results
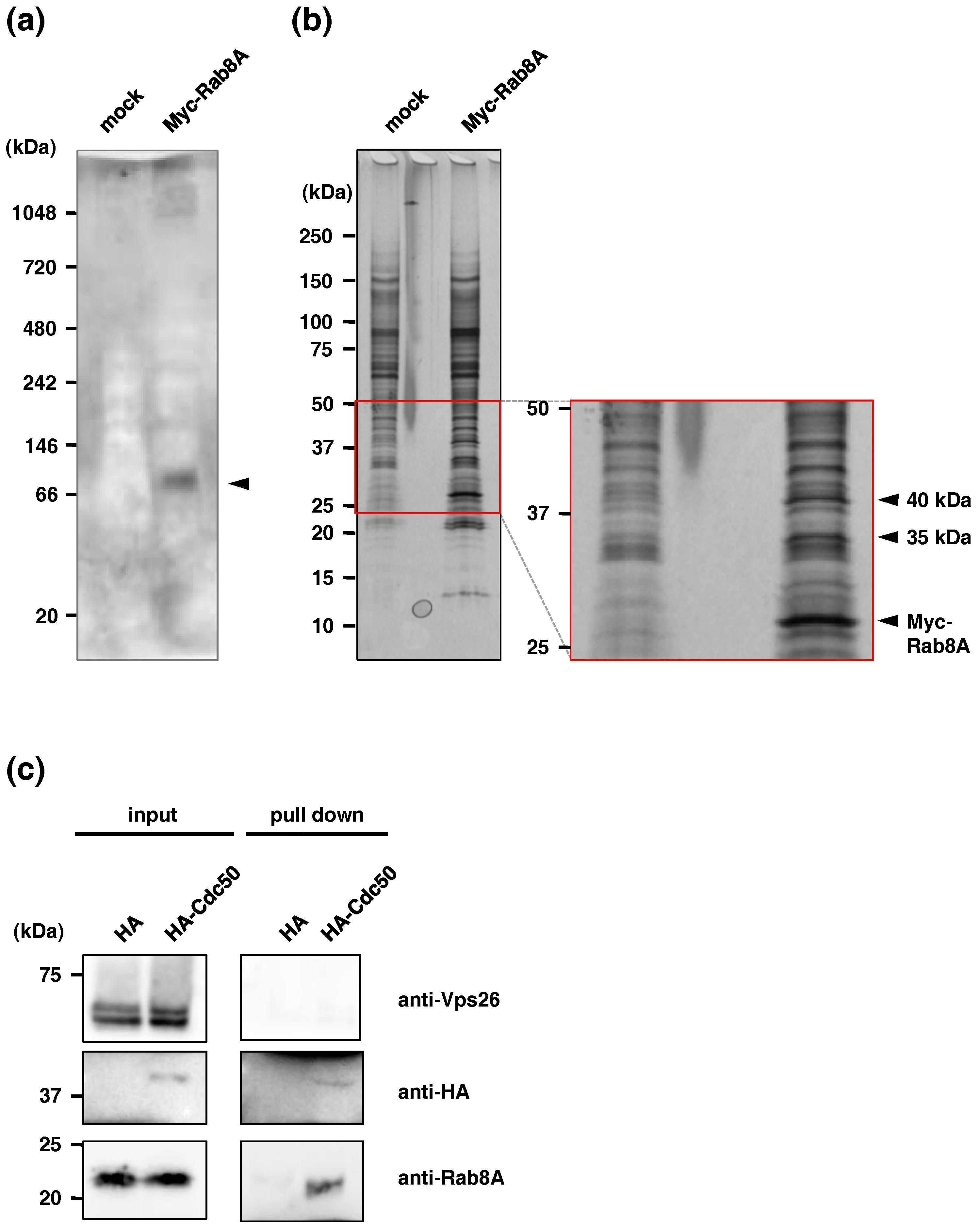
2.1. Identification of EhCdc50 As an EhRab8A Interacting Protein in E. histolytica
2.2. Confirmation of Binding of EhCdc50 and EhRab8A
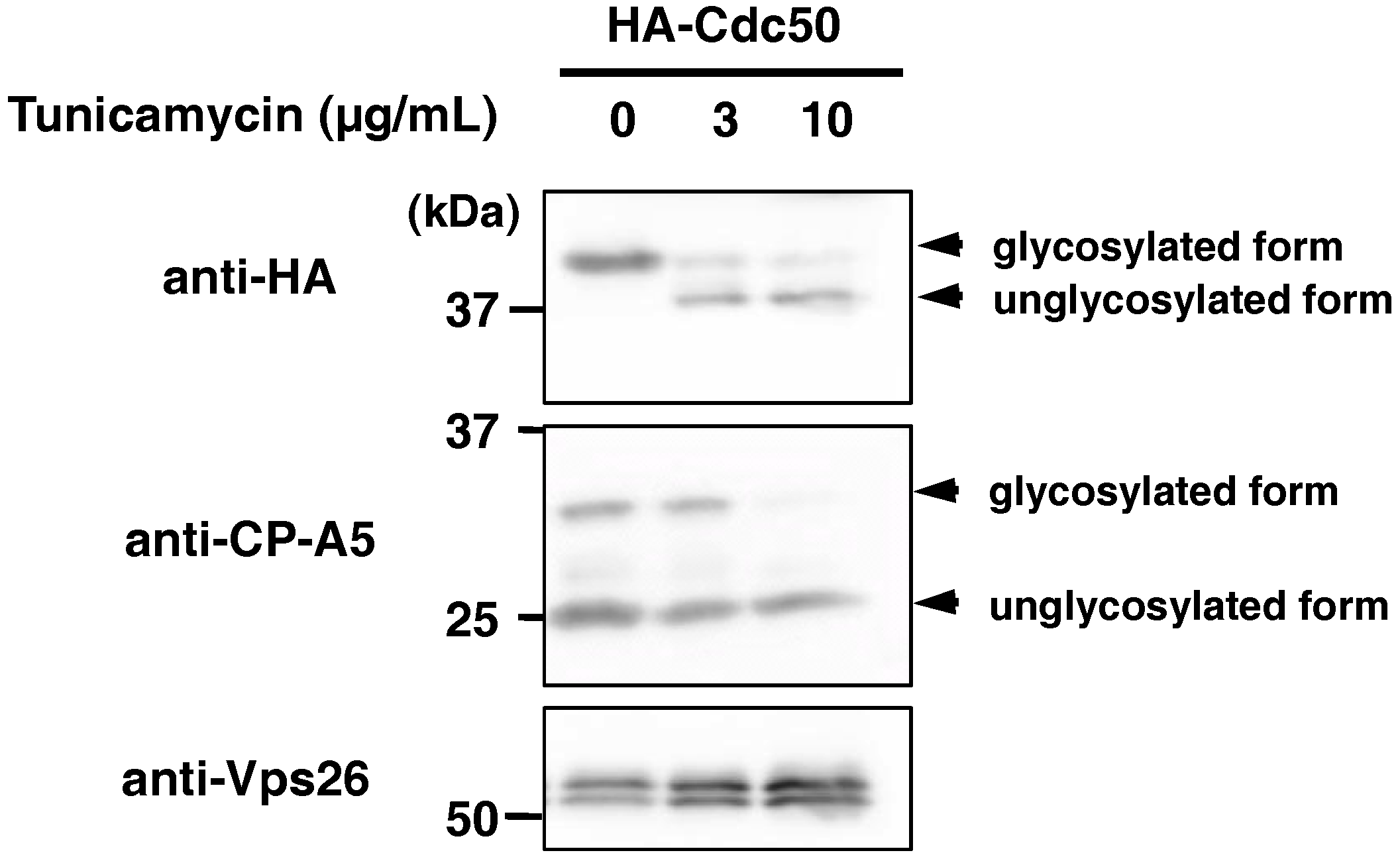
2.3. EhCdc50 is N-Glycosylated Protein
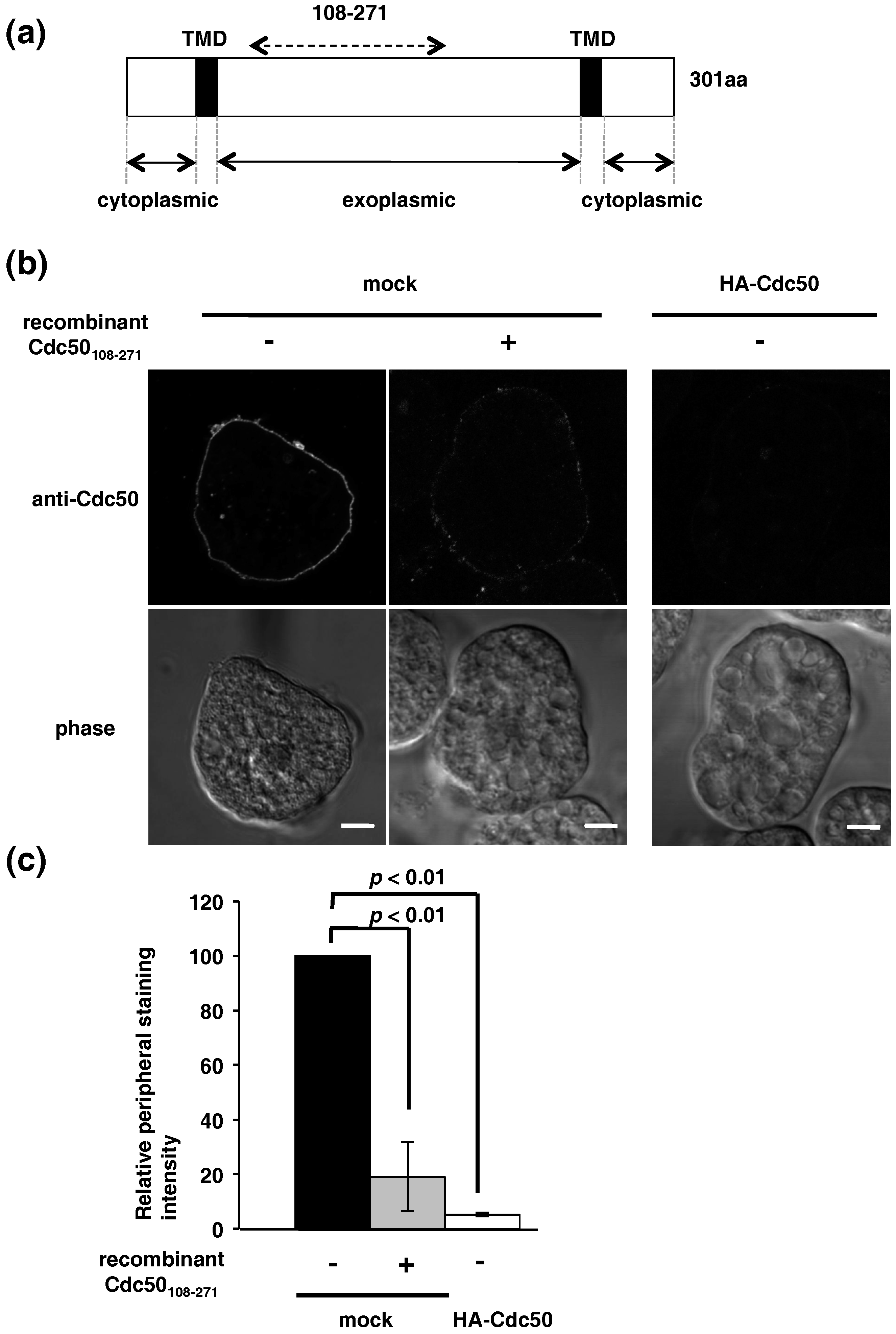
2.4. Localization of Endogenous EhCdc50 and Exogenous Epitope-Tagged EhCdc50 and Inhibition of Its Transport to the Plasma Membrane by Overexpression
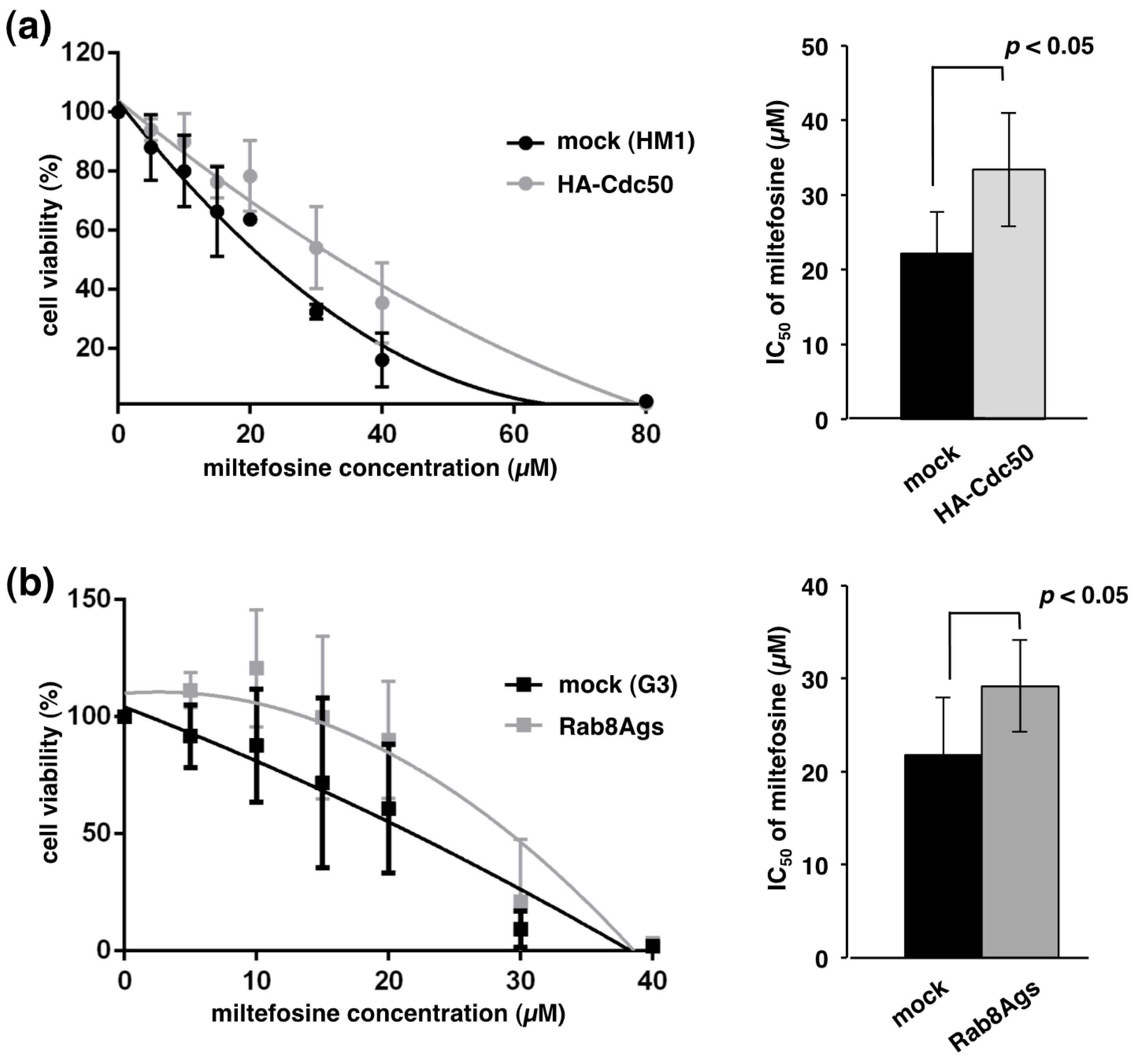
2.5. Overexpression of EhCdc50 or Gene Silencing of EhRab8A Showed Decreased Miltefosine Susceptibility
3. Discussion
3.1. Identification of EhCdc50 as EhRab8A Binding Protein
3.2. N-Linked Oligosaccharide Modification and Possible Role of EhCdc50 in the Exit From the ER
3.3. A Possible Novel Rab8A-Dependent and COPII-Independent Pathway for Cdc50 Traffic
3.4. EhRab8A and EhCdc50 Are Involved in Miltefosine Sensitivity in Entamoeba
4. Materials and Methods
4.1. Culture
4.2. Antibodies
4.3. Creation of E. histolytica Transformant Lines
4.4. Blue-Native Polyacrylamide gel Electrophoresis (BN-PAGE)
4.5. Coimmunoprecipitation
4.6. Liquid Chromatography-Tandem Mass Spectrometric Analysis (LC-MS/MS)
4.7. Production of EhCdc50 Antibody
4.8. Indirect Immunofluorescence Assay
4.9. Miltefosine Sensitivity Assay
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Annotation | ID | Molecular Weight (kDa) | Normalized Relative Ratio against Common Peptides (mock) | Normalized Relative Ratio against Common Peptides (mycRab8A) | E-Value (Species) |
|---|---|---|---|---|---|
| 40 kDa | |||||
| small GTP-binding protein Rab8A | EHI_199820 | 22 | 0 | 3.03 | 1.9 × 10−58 (Dictyostelium discoideum) |
| EhCP-C6 | EHI_127030 | 58 | 0 | 2.02 | |
| tldc domain-containing protein | EHI_134660 | 42 | 0 | 1.01 | 4 × 10−7 (Heterostelium album) |
| MIT domain protein | EHI_093860 | 76 | 0 | 1.01 | |
| Sphingomyelinase phosphodiesterase | EHI_100080 | 46 | 0 | 1.01 | 1.4 × 10−39 (Dictyostelium discoideum) |
| Vacuolar ATP synthase subunit δ | EHI_106350 | 40 | 0 | 1.01 | 1.3 × 10−100 (Dictyostelium discoideum) |
| AIG1 family protein | EHI_109120 | 23 | 0 | 1.01 | 5.6 × 10−8 (Homo sapiens) |
| Lysophospholipid acyltransferase 1 | EHI_086180 | 57 | 0 | 1.01 | 7 × 10−45 (Arabidopsis thaliana) |
| Ubiquitin | EHI_166800 | 9 | 0 | 1.01 | 5.2 × 10−33 (Encephalitozoon cuniculi) |
| Sulfate adenylyltransferase (mitosomal protein) | EHI_197160 | 48 | 0 | 1.01 | 8.8 × 10−141 (Desulfovibrio desulfuricans) |
| Glycerophosphodiester phosphodiesterase | EHI_068320 | 45 | 0.84 | 2.02 | 4.6 × 10−25 (Bacillus subtilis) |
| Sec61 alpha subunit | EHI_164500 | 52 | 1.68 | 6.06 | 5.7 × 10−66 (Dictyostelium discoideum) |
| C2 domain containing protein | EHI_069950 | 37 | 4.19 | 9.09 | 5.1 × 10−7 (Arabidopsis thaliana) |
| 35 kDa | |||||
| small GTP-binding protein Rab8A | EHI_199820 | 22 | 0 | 4.0 | 1.9 × 10−58 (Dictyostelium discoideum) |
| Cdc50 | EHI_142740 | 36 | 0 | 2.7 | 1.3 × 10−43 (Arabidopsis thaliana) |
| Guanine nucleotide-binding Ribosomal protein | EHI_050550 | 35 | 0.9 | 5.4 | 6.5 × 10−74 (Biomphalaria glabrata) |
| Translation initiation factor | EHI_180410 | 33 | 0.9 | 2.7 | 1.5 × 10−23 (Proteus mirabilis) |
| Guanine nucleotide-binding Ribosomal protein | EHI_171280 | 35 | 0.9 | 2.7 | 6.3 × 10−72 (Homo sapiens) |
| Nuclear pore protein | EHI_118780 | 38 | 2.8 | 9.4 | 5.8 × 10−18 (Chaetomium thermophilum) |
References
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Hutagalung, A.H.; Novick, P.J. Role of Rab GTPases in membrane traffic and cell physiology. Physiol. Rev. 2011, 91, 119–149. [Google Scholar] [CrossRef] [PubMed]
- Carroll, K.S.; Hanna, J.; Simon, I.; Krise, J.; Barbero, P.; Pfeffer, S.R. Role of Rab9 GTPase in facilitating receptor recruitment by TIP47. Science 2001, 292, 1373–1376. [Google Scholar] [CrossRef] [PubMed]
- McLauchlan, H.; Newell, J.; Morrice, N.; Osborne, A.; West, M.; Smythe, E. A novel role for Rab5-GDI in ligand sequestration into clathrin-coated pits. Curr. Biol. 1998, 8, 34–45. [Google Scholar] [CrossRef]
- Hales, C.M.; Vaerman, J.P.; Goldenring, J.R. Rab11 family interacting protein 2 associates with Myosin Vb and regulates plasma membrane recycling. J. Biol. Chem. 2002, 277, 50415–50421. [Google Scholar] [CrossRef] [PubMed]
- Ménasché, G.; Pastural, E.; Feldmann, J.; Certain, S.; Ersoy, F.; Dupuis, S.; Wulffraat, N.; Bianchi, D.; Fischer, A.; le Deist, F.; et al. Mutations in RAB27A cause Griscelli syndrome associated with haemophagocytic syndrome. Nat. Genet. 2000, 25, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Novick, P.J. A ras-like protein is required for a post-Golgi event in yeast secretion. Cell 1987, 49, 527–538. [Google Scholar] [CrossRef]
- Mizuno-Yamasaki, E.; Rivera-Molina, F.; Novick, P. GTPase networks in membrane traffic. Annu. Rev. Biochem. 2012, 81, 637–659. [Google Scholar] [CrossRef]
- Homma, Y.; Fukuda, M. Rabin8 regulates neurite outgrowth in both GEF activity-dependent and -independent manners. Mol. Biol. Cell 2016, 27, 2107–2118. [Google Scholar] [CrossRef]
- Blacque, O.E.; Scheidel, N.; Kuhns, S. Rab GTPases in cilium formation and function. Small GTPases 2018, 9, 76–94. [Google Scholar] [CrossRef]
- Du, F.; Edwards, K.; Shen, Z.; Sun, B.; De Lozanne, A.; Briggs, S.; Firtel, R.A. Regulation of contractile vacuole formation and activity in Dictyostelium. EMBO J. 2008, 27, 2064–2076. [Google Scholar] [CrossRef]
- Ralston, K.S. Chew on this: Amoebic trogocytosis and host cell killing by Entamoeba histolytica. Trends Parasitol. 2015, 31, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Santi-Rocca, J.; Rigothier, M.C.; Guillen, N. Host-microbe interactions and defense mechanisms in the development of amoebic liver abscesses. Clin. Microbiol. Rev. 2009, 22, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Nakada-Tsukui, K.; Saito-Nakano, Y.; Husain, A.; Nozaki, T. Conservation and function of Rab small GTPases in Entamoeba: Annotation of E. invadens Rab and its use for the understanding of Entamoeba biology. Exp. Parasitol. 2010, 126, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Nakada-Tsukui, K.; Tsuboi, K.; Furukawa, A.; Yamada, Y.; Nozaki, T. A novel class of cysteine protease receptors that mediate lysosomal transport. Cell. Microbiol. 2012, 14, 1299–1317. [Google Scholar] [CrossRef] [Green Version]
- Emmanuel, M.; Nakano, Y.S.; Nozaki, T.; Datta, S. Small GTPase Rab21 mediates fibronectin induced actin reorganization in Entamoeba histolytica: Implications in pathogen invasion. PLoS Pathog. 2015, 11, e1004666. [Google Scholar] [CrossRef] [PubMed]
- Hanadate, Y.; Saito-Nakano, Y.; Nakada-Tsukui, K.; Nozaki, T. Endoplasmic reticulum-resident Rab8A GTPase is involved in phagocytosis in the protozoan parasite Entamoeba histolytica. Cell. Microbiol. 2016, 18, 1358–1373. [Google Scholar] [CrossRef]
- Verma, K.; Datta, S. The Monomeric GTPase Rab35 Regulates Phagocytic Cup Formation and Phagosomal Maturation in Entamoeba histolytica. J. Biol. Chem. 2017, 292, 4960–4975. [Google Scholar] [CrossRef]
- Nakada-Tsukui, K.; Saito-Nakano, Y.; Ali, V.; Nozaki, T. A retromerlike complex is a novel Rab7 effector that is involved in the transport of the virulence factor cysteine protease in the enteric protozoan parasite Entamoeba histolytica. Mol. Biol. Cell 2005, 16, 5294–5303. [Google Scholar] [CrossRef]
- Rojas, R.; van Vlijmen, T.; Mardones, G.A.; Prabhu, Y.; Rojas, A.L.; Mohammed, S.; Heck, A.J.R.; Raposo, G.; van der Sluijs, P.; Bonifacino, J.S. Regulation of retromer recruitment to endosomes by sequential action of Rab5 and Rab7. J. Cell Biol. 2008, 183, 513–526. [Google Scholar] [CrossRef] [Green Version]
- Hattula, K.; Peranen, J. FIP-2, a coiled-coil protein, links Huntingtin to Rab8 and modulates cellular morphogenesis. Curr. Biol. 2000, 10, 1603–1606. [Google Scholar] [CrossRef]
- Hattula, K.; Furuhjelm, J.; Arffman, A.; Peranen, J. A Rab8-specific GDP/GTP exchange factor is involved in actin remodeling and polarized membrane transport. Mol. Biol. Cell 2002, 13, 3268–3280. [Google Scholar] [CrossRef] [PubMed]
- Bredeston, L.M.; Caffaro, C.E.; Samuelson, J.; Hirschberg, C.B. Golgi and endoplasmic reticulum functions take place in different subcellular compartments of Entamoeba histolytica. J. Biol. Chem. 2005, 280, 32168–32176. [Google Scholar] [CrossRef] [PubMed]
- Brandizzi, F.; Barlowe, C. Organization of the ER-Golgi interface for membrane traffic control. Nat. Rev. Mol. Cell Biol. 2013, 14, 382–392. [Google Scholar] [CrossRef]
- Costa, S.; Marek, M.; Axelsen, K.B.; Theorin, L.; Pomorski, T.G.; López-Marqués, R.L. Role of post-translational modifications at the beta-subunit ectodomain in complex association with a promiscuous plant P4-ATPase. Biochem. J. 2016, 473, 1605–1615. [Google Scholar] [CrossRef] [PubMed]
- Jacquot, A.; Montigny, C.; Hennrich, H.; Barry, R.; le Maire, M.; Jaxel, C.; Holthuis, J.; Champeil, P.; Lenoir, G. Phosphatidylserine stimulation of Drs2p/Cdc50p lipid translocase dephosphorylation is controlled by phosphatidylinositol-4-phosphate. J. Biol. Chem. 2012, 287, 13249–13261. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.A.; Molday, R.S. Critical role of the beta-subunit CDC50A in the stable expression, assembly, subcellular localization, and lipid transport activity of the P4-ATPase ATP8A2. J. Biol. Chem. 2011, 286, 17205–17216. [Google Scholar] [CrossRef]
- Muthusamy, B.P.; Natarajan, P.; Zhou, X.; Graham, T.R. Linking phospholipid flippases to vesicle-mediated protein transport. Biochim. Biophys. Acta 2009, 1791, 612–619. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Fujimura-Kamada, K.; Yamamoto, T. Functions of phospholipid flippases. J. Biochem. 2011, 149, 131–143. [Google Scholar] [CrossRef]
- Lopez-Marqués, R.L.; Poulsen, L.R.; Hanisch, S.; Meffert, K.; Buch-Pedersen, M.J.; Jakobsen, M.K.; Pomorski, T.G.; Palmgren, M.G. Intracellular targeting signals and lipid specificity determinants of the ALA/ALIS P4-ATPase complex reside in the catalytic ALA alpha-subunit. Mol. Biol. Cell 2010, 21, 791–801. [Google Scholar] [CrossRef]
- Saito, K.; Fujimura-Kamada, K.; Furuta, N.; Kato, U.; Umeda, M.; Tanaka, K. Cdc50p, a protein required for polarized growth, associates with the Drs2p P-type ATPase implicated in phospholipid translocation in Saccharomyces cerevisiae. Mol. Biol. Cell 2004, 15, 3418–3432. [Google Scholar] [CrossRef] [PubMed]
- Van der Velden, L.M.; Wichers, C.G.; van Breevoort, A.E.; Coleman, J.A.; Molday, R.S.; Berger, R.; Klomp, L.W.J.; van de Graaf, S.F.J. Heteromeric interactions required for abundance and subcellular localization of human CDC50 proteins and class 1 P4-ATPases. J. Biol. Chem. 2010, 285, 40088–40096. [Google Scholar] [CrossRef] [PubMed]
- Seifert, K.; Duchêne, M.; Wernsdorfer, W.H.; Kollaritsch, H.; Scheiner, O.; Wiedermann, G.; Hottkowitz, T.; Eibl, H. Effects of miltefosine and other alkylphosphocholines on human intestinal parasite Entamoeba histolytica. Antimicrob. Agents Chemother. 2001, 45, 1505–1510. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Victoria, J.M.; Cortés-Selva, F.; Parodi-Talice, A.; Bavchvarov, B.I.; Pérez-Victoria, F.J.; Muñoz-Martínez, F.; Maitrejean, M.; Costi, M.P.; Barron, D.; di Pietro, A.; et al. Combination of suboptimal doses of inhibitors targeting different domains of LtrMDR1 efficiently overcomes resistance of Leishmania spp. to Miltefosine by inhibiting drug efflux. Antimicrob. Agents Chemother. 2006, 50, 3102–3110. [Google Scholar] [CrossRef] [PubMed]
- Perez-Victoria, F.J.; Sanchez-Canete, M.P.; Castanys, S.; Gamarro, F. Phospholipid translocation and miltefosine potency require both L. donovani miltefosine transporter and the new protein LdRos3 in Leishmania parasites. J. Biol. Chem. 2006, 281, 23766–23775. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Victoria, F.J.; Sánchez-Cañete, M.P.; Seifert, K.; Croft, S.L.; Sundar, S.; Castanys, S.; Gamarro, F. Mechanisms of experimental resistance of Leishmania to miltefosine: Implications for clinical use. Drug Resist. Updates 2006, 9, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Diekmann, Y.; Seixas, E.; Gouw, M.; Tavares-Cadete, F.; Seabra, M.C.; Pereira-Leal, J.B. Thousands of rab GTPases for the cell biologist. PLoS Comput. Biol. 2011, 7, e1002217. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Leal, J.B.; Seabra, M.C. Evolution of the Rab family of small GTP-binding proteins. J. Mol. Biol. 2001, 313, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Kremer, K.; Kamin, D.; Rittweger, E.; Wilkes, J.; Flammer, H.; Mahler, S.; Heng, J.; Tonkin, C.J.; Langsley, G.; Hell, S.W.; et al. An overexpression screen of Toxoplasma gondii Rab-GTPases reveals distinct transport routes to the micronemes. PLoS Pathog. 2013, 9, e1003213. [Google Scholar] [CrossRef]
- Ackers, J.P.; Dhir, V.; Field, M.C. A bioinformatic analysis of the RAB genes of Trypanosoma brucei. Mol. Biochem. Parasitol. 2005, 141, 89–97. [Google Scholar] [CrossRef]
- Magnelli, P.; Cipollo, J.F.; Ratner, D.M.; Cui, J.; Kelleher, D.; Gilmore, R.; Costello, C.E.; Robbins, P.W.; Samuelson, J. Unique Asn-linked oligosaccharides of the human pathogen Entamoeba histolytica. J. Biol. Chem. 2008, 283, 18355–18364. [Google Scholar] [CrossRef] [PubMed]
- Graef, M.; Friedman, J.R.; Graham, C.; Babu, M.; Nunnari, J. ER exit sites are physical and functional core autophagosome biogenesis components. Mol. Biol. Cell 2013, 24, 2918–2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, T.; Takatsu, H.; Miyano, R.; Takada, N.; Nakayama, K.; Shin, H.W. Phospholipid Flippase ATP10A Translocates Phosphatidylcholine and Is Involved in Plasma Membrane Dynamics. J. Biol. Chem. 2015, 290, 15004–15017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, S.; Pratte, D.; Schwarz, H.; Pipkorn, R.; Singer-Kruger, B. Oligomeric Dop1p is part of the endosomal Neo1p-Ysl2p-Arl1p membrane remodeling complex. Traffic 2010, 11, 1092–1106. [Google Scholar] [CrossRef] [PubMed]
- Takatsu, H.; Baba, K.; Shima, T.; Umino, H.; Kato, U.; Umeda, M.; Nakayama, K.; Shin, H.-W. ATP9B, a P4-ATPase (a putative aminophospholipid translocase), localizes to the trans-Golgi network in a CDC50 protein-independent manner. J. Biol. Chem. 2011, 286, 38159–38167. [Google Scholar] [CrossRef] [PubMed]
- Geilen, C.C.; Wieder, T.; Reutter, W. Hexadecylphosphocholine inhibits translocation of CTP:choline-phosphate cytidylyltransferase in Madin-Darby canine kidney cells. J. Biol. Chem. 1992, 267, 6719–6724. [Google Scholar] [PubMed]
- Ramakrishnan, S.; Serricchio, M.; Striepen, B.; Butikofer, P. Lipid synthesis in protozoan parasites: A comparison between kinetoplastids and apicomplexans. Prog. Lipid Res. 2013, 52, 488–512. [Google Scholar] [CrossRef] [Green Version]
- Perez-Victoria, F.J.; Castanys, S.; Gamarro, F. Leishmania donovani resistance to miltefosine involves a defective inward translocation of the drug. Antimicrob. Agents Chemother. 2003, 47, 2397–2403. [Google Scholar] [CrossRef]
- Coelho, A.C.; Trinconi, C.T.; Costa, C.H.; Uliana, S.R. In vitro and in vivo miltefosine susceptibility of a Leishmania amazonensis isolate from a patient with diffuse cutaneous leishmaniasis. PLoS Negl. Trop. Dis. 2014, 8, e2999. [Google Scholar] [CrossRef]
- Coelho, A.C.; Boisvert, S.; Mukherjee, A.; Leprohon, P.; Corbeil, J.; Ouellette, M. Multiple mutations in heterogeneous miltefosine-resistant Leishmania major population as determined by whole genome sequencing. PLoS Negl. Trop. Dis. 2012, 6, e1512. [Google Scholar] [CrossRef]
- Huang, W.; Liao, G.; Baker, G.M.; Wang, Y.; Lau, R.; Paderu, P.; Perlin, D.S.; Xue, C. Lipid flippase subunit Cdc50 mediates drug resistance and virulence in Cryptococcus neoformans. MBio 2016, 7, e00478-16. [Google Scholar] [CrossRef] [PubMed]
- Jeelani, G.; Nozaki, T. Metabolomic analysis of Entamoeba: Applications and implications. Curr. Opin. Microbiol. 2014, 20, 118–124. [Google Scholar] [CrossRef]
- Diamond, L.S.; Harlow, D.R.; Cunnick, C.C. A new medium for the axenic cultivation of Entamoeba histolytica and other Entamoeba. Trans. R. Soc. Trop. Med. Hyg. 1978, 72, 431–432. [Google Scholar] [CrossRef]
- Loftus, B.; Anderson, I.; Davies, R.; Alsmark, U.C.M.; Samuelson, J.; Amedeo, P.; Roncaglia, P.; Berriman, M.; Hirt, R.P.; Mann, B.J.; et al. The genome of the protist parasite Entamoeba histolytica. Nature 2005, 433, 865–868. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Nozaki, T. Non-vesicular lipid transport machinery in Entamoeba histolytica. Front. Cell. Infect. Microbiol. 2018, 8, 315. [Google Scholar] [CrossRef] [PubMed]
- Pina-Vazquez, C.; Reyes-Lopez, M.; Mendoza-Hernandez, G.; Bermudez-Cruz, R.M.; de la Garza, M. Identification of phosphatidylcholine transfer protein-like in the parasite Entamoeba histolytica. Biochimie 2014, 107 Pt B, 223–234. [Google Scholar] [CrossRef]
- Diamond, L.S.; Mattern, C.F.; Bartgis, I.L. Viruses of Entamoeba histolytica. I. Identification of transmissible virus-like agents. J. Virol. 1972, 9, 326–341. [Google Scholar] [PubMed]
- Bracha, R.; Nuchamowitz, Y.; Anbar, M.; Mirelman, D. Transcriptional silencing of multiple genes in trophozoites of Entamoeba histolytica. PLoS Pathog. 2006, 2, e48. [Google Scholar] [CrossRef]
- Nozaki, T.; Asai, T.; Sanchez, L.B.; Kobayashi, S.; Nakazawa, M.; Takeuchi, T. Characterization of the gene encoding serine acetyltransferase, a regulated enzyme of cysteine biosynthesis from the protist parasites Entamoeba histolytica and Entamoeba dispar. Regulation and possible function of the cysteine biosynthetic pathway in Entamoeba. J. Biol. Chem. 1999, 274, 32445–32452. [Google Scholar]
- Ali IK, M.; Haque, R.; Siddique, A.; Kabir, M.; Sherman, N.E.; Gray, S.A.; Cangelosi, G.A.; Petri, W.A., Jr. Proteomic analysis of the cyst stage of Entamoeba histolytica. PLoS Negl. Trop. Dis. 2012, 6, e1643. [Google Scholar] [CrossRef]
- Makiuchi, T.; Mi-ichi, F.; Nakada-Tsukui, K.; Nozaki, T. Novel TPR-containing subunit of TOM complex functions as cytosolic receptor for Entamoeba mitosomal transport. Sci. Rep. 2013, 3, 1129. [Google Scholar] [CrossRef] [PubMed]
- Saito-Nakano, Y.; Yasuda, T.; Nakada-Tsukui, K.; Leippe, M.; Nozaki, T. Rab5-associated vacuoles play a unique role in phagocytosis of the enteric protozoan parasite Entamoeba histolytica. J. Biol. Chem. 2004, 279, 49497–49507. [Google Scholar] [CrossRef] [PubMed]
- Penuliar, G.M.; Nakada-Tsukui, K.; Nozaki, T. Phenotypic and transcriptional profiling in Entamoeba histolytica reveal costs to fitness and adaptive responses associated with metronidazole resistance. Front. Microbiol. 2015, 6, 354. [Google Scholar] [CrossRef] [PubMed]





| Annotation | Gene ID | Molecular Weight (kDa) | Normalized Relative Ratio against Common Peptides, mock/mycRab8A | Subcellular Localization in Other Organisms | E-Value (Species) |
|---|---|---|---|---|---|
| Cdc50 | EHI_142740 | 36 | 0/2.68 | trans-Golgi/endosome/PM/ER | 1.3 × 10−43 (Arabidopsis thaliana) |
| Nuclear pore protein | EHI_118780 | 38 | 2.80/9.36 | nucleus | 5.8 × 10−18 (Chaetomiu thermopholum) |
| Annotation | Gene ID | Molecular Weight (kDa) | Normalized Relative Ratio against Common Peptides, mock/mycRab8A | Subcellular Localization in Other Organisms | E-Value (Species) |
|---|---|---|---|---|---|
| Sphingomyelinase phosphodiesterase | EHI_100080 | 46 | 0/1.02 | Acid organella | 1.4 × 10−39 (Dictyostelium discoideum) |
| tldc domain-containing protein | EHI_134660 | 42 | 0/1.01 | cytosol | 4 × 10−7 (Heterostelium album) |
| Vacuolar ATP synthase subunit δ | EHI_106350 | 40 | 0/1.01 | lysosome membrane | 1.3 × 10−100 (Dictyostelium discoideum) |
| Sulfate adenylyltransferase | EHI_197160 | 48 | 0/1.01 | amoebic mitosome lumen | 8.8 × 10−141 (Desulfovibrio desulfuricans) |
| Glycerophosphodiester phosphodiesterase | EHI_068320 | 45 | 0.84/2.02 | ER | 4.6 × 10−25 (Bacillus subtilis) |
| C2 domain containing protein | EHI_069950 | 37 | 4.19/9.09 | cytosol | 5.1 × 10−7 (Arabidopsis thaliana) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanadate, Y.; Saito-Nakano, Y.; Nakada-Tsukui, K.; Nozaki, T. Identification and Characterization of the Entamoeba Histolytica Rab8a Binding Protein: A Cdc50 Homolog. Int. J. Mol. Sci. 2018, 19, 3831. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19123831
Hanadate Y, Saito-Nakano Y, Nakada-Tsukui K, Nozaki T. Identification and Characterization of the Entamoeba Histolytica Rab8a Binding Protein: A Cdc50 Homolog. International Journal of Molecular Sciences. 2018; 19(12):3831. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19123831
Chicago/Turabian StyleHanadate, Yuki, Yumiko Saito-Nakano, Kumiko Nakada-Tsukui, and Tomoyoshi Nozaki. 2018. "Identification and Characterization of the Entamoeba Histolytica Rab8a Binding Protein: A Cdc50 Homolog" International Journal of Molecular Sciences 19, no. 12: 3831. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19123831