Interaction of the GTPase Elongation Factor Like-1 with the Shwachman-Diamond Syndrome Protein and Its Missense Mutations

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
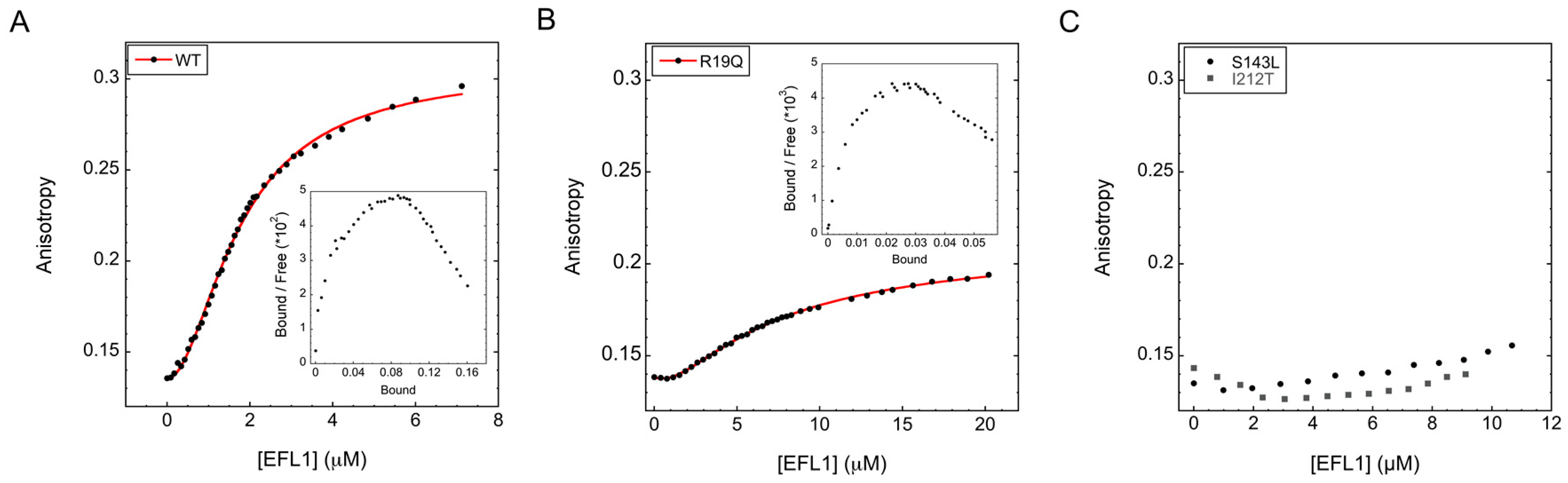
2.1. Interaction between Human EFL1 and SBDS Missense Variants
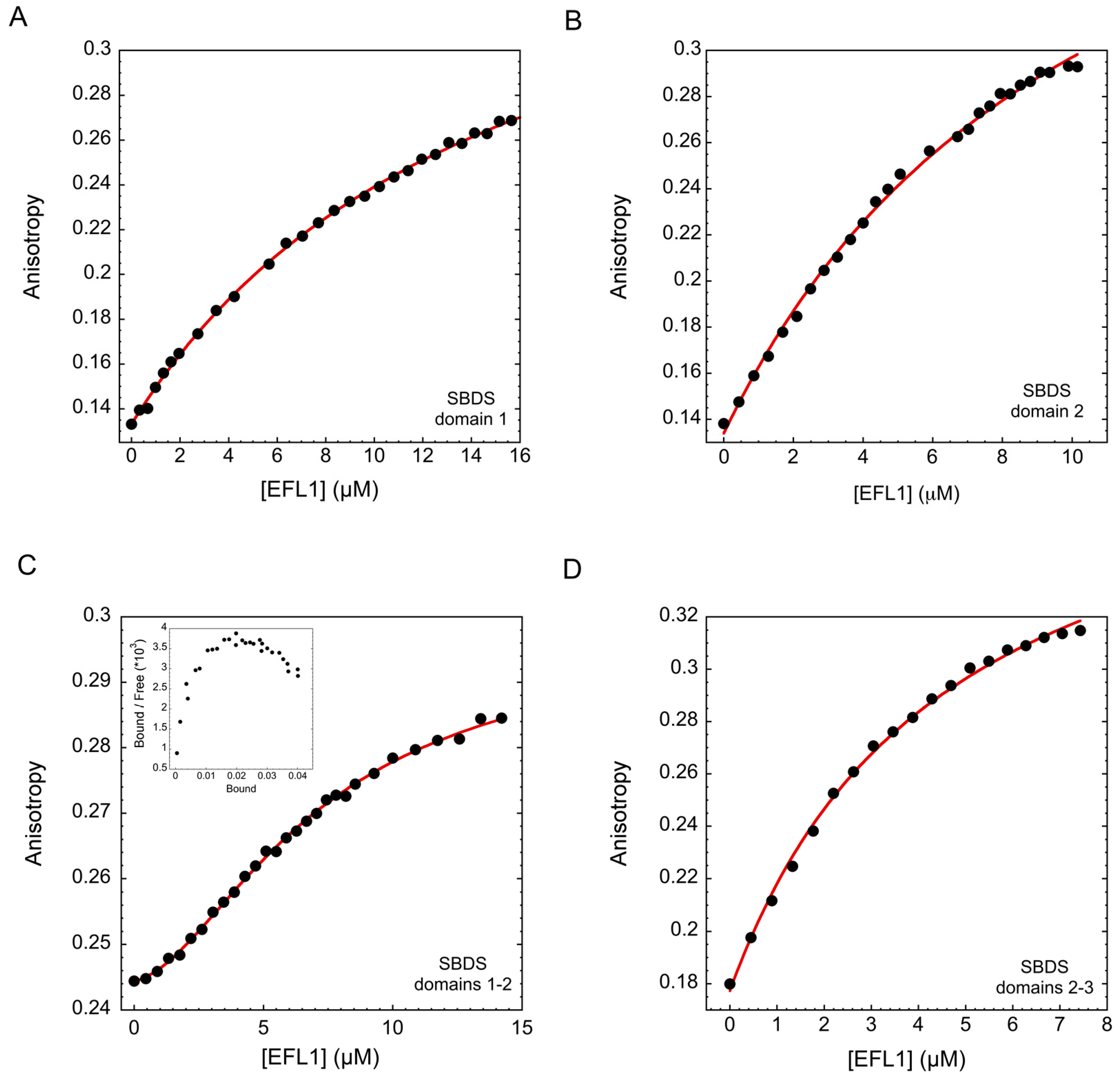
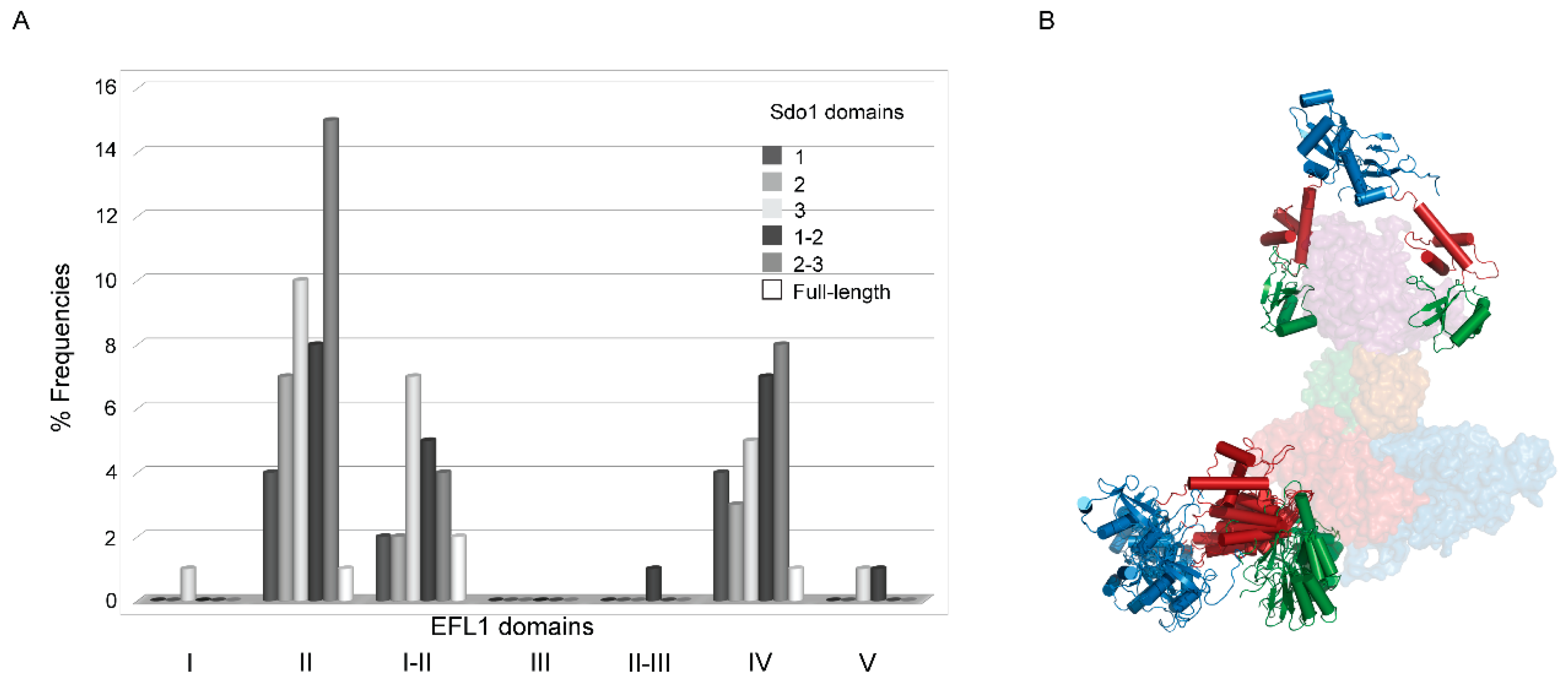
2.2. Interaction between Human EFL1 and the Different Domains of SBDS
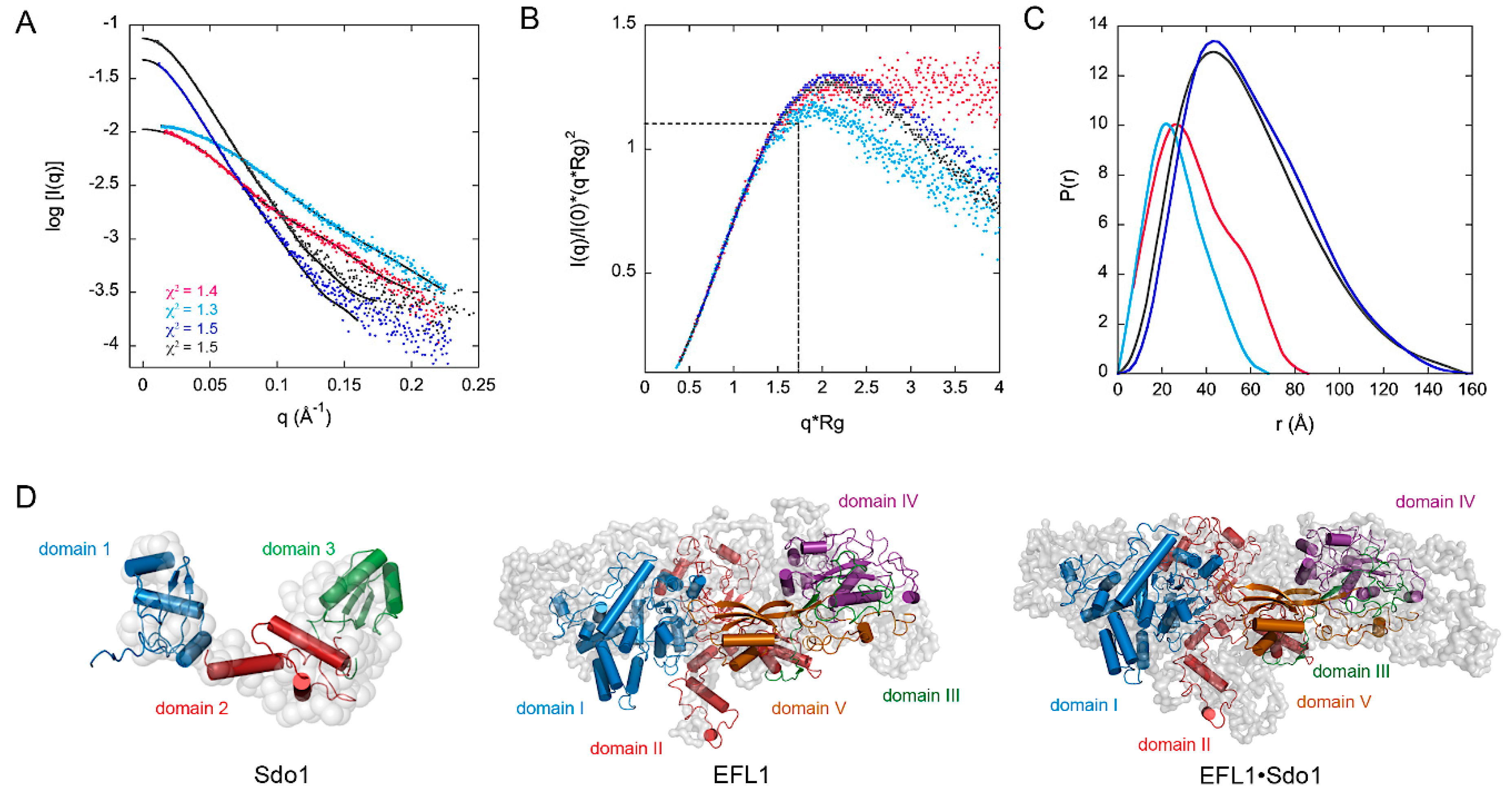
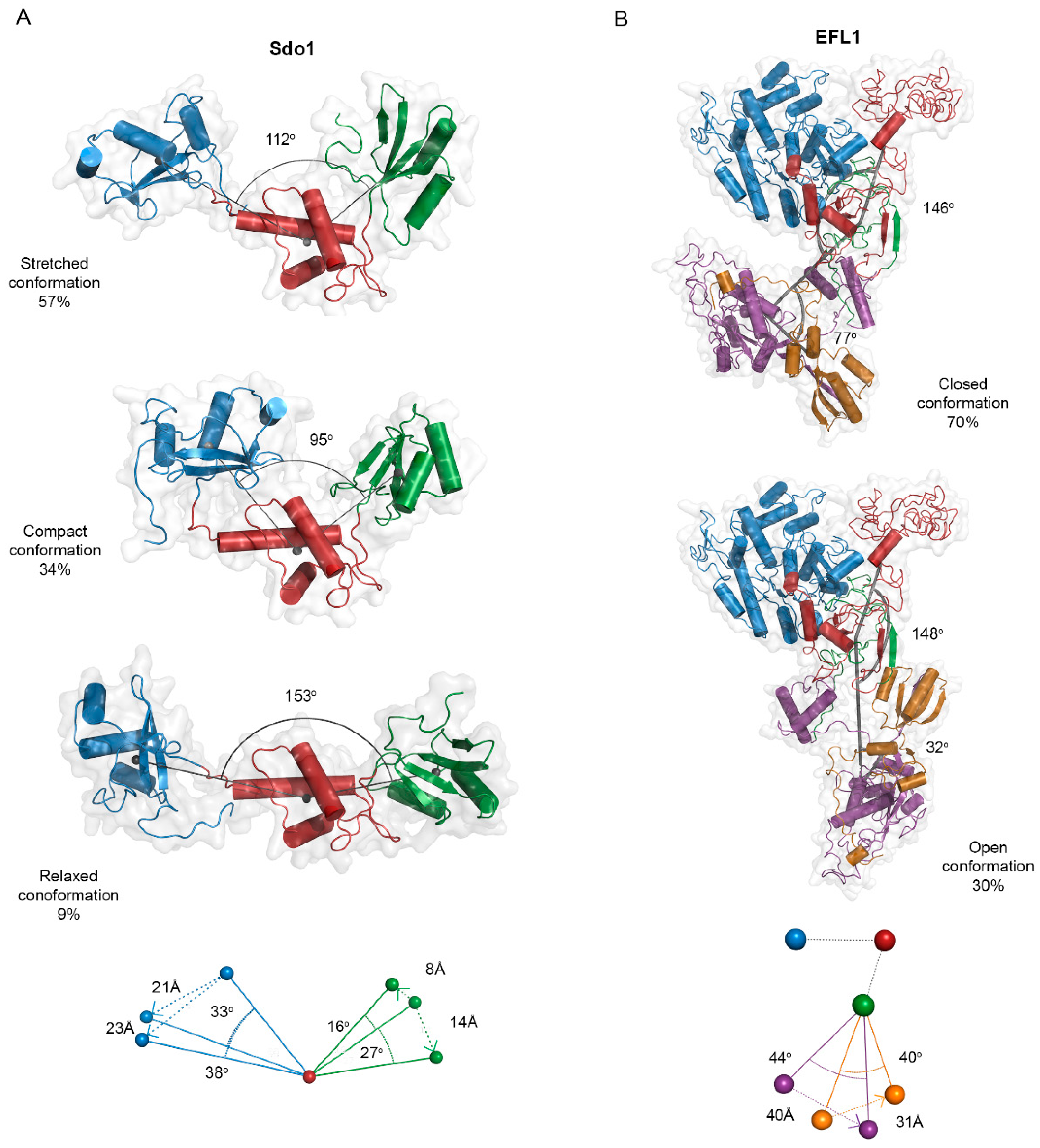
2.3. Small Angle X-ray Scattering and Flexibility Analysis of the Yeast SBDS, EFL1 and the EFL1•SBDS Complex
3. Discussion
3.1. Molecular Function of the SBDS Missense Mutations on SDS
3.2. Interaction Mechanism between EFL1 and SBDS
4. Materials and Methods
4.1. Protein Expression and Purification
4.2. Fluorescence Anisotropy Experiments
4.3. Small Angle X-ray Scattering (SAXS) Experiments
4.4. SAXS Data Modelling and Flexibility Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
Abbreviations
| EFL1 | Elongation Factor like-1 protein |
| SBDS | Shwachman-Bodian-Diamond Syndrome protein |
| SDS | Shwachman-Diamond Syndrome |
| eIF6 | eukaryotic Initiation Factor 6 |
| SAXS | Small-Angle X-ray Scattering |
References
- Dror, Y.; Donadieu, J.; Koglmeier, J.; Dodge, J.; Toiviainen-Salo, S.; Makitie, O.; Kerr, E.; Zeidler, C.; Shimamura, A.; Shah, N.; et al. Draft consensus guidelines for diagnosis and treatment of Shwachman-Diamond syndrome. Ann. N. Y. Acad. Sci. 2011, 1242, 40–55. [Google Scholar] [CrossRef] [Green Version]
- Donadieu, J.; Leblanc, T.; Bader Meunier, B.; Barkaoui, M.; Fenneteau, O.; Bertrand, Y.; Maier-Redelsperger, M.; Micheau, M.; Stephan, J.L.; Phillipe, N.; et al. Analysis of risk factors for myelodysplasias, leukemias and death from infection among patients with congenital neutropenia. Experience of the French Severe Chronic Neutropenia Study Group. Haematologica 2005, 90, 45–53. [Google Scholar]
- Boocock, G.R.; Morrison, J.A.; Popovic, M.; Richards, N.; Ellis, L.; Durie, P.R.; Rommens, J.M. Mutations in SBDS are associated with Shwachman-Diamond syndrome. Nat. Genet. 2003, 33, 97–101. [Google Scholar] [CrossRef]
- Costa, E.; Duque, F.; Oliveira, J.; Garcia, P.; Goncalves, I.; Diogo, L.; Santos, R. Identification of a novel AluSx-mediated deletion of exon 3 in the SBDS gene in a patient with Shwachman-Diamond syndrome. Blood Cells Mol. Dis. 2007, 39, 96–101. [Google Scholar] [CrossRef]
- Costa, E.; Santos, R. Hematologically important mutations: Shwachman-Diamond syndrome. Blood Cells Mol. Dis. 2008, 40, 183–184. [Google Scholar] [CrossRef]
- Nakashima, E.; Mabuchi, A.; Makita, Y.; Masuno, M.; Ohashi, H.; Nishimura, G.; Ikegawa, S. Novel SBDS mutations caused by gene conversion in Japanese patients with Shwachman-Diamond syndrome. Hum. Genet. 2004, 114, 345–348. [Google Scholar]
- Nicolis, E.; Bonizzato, A.; Assael, B.M.; Cipolli, M. Identification of novel mutations in patients with Shwachman-Diamond syndrome. Hum. Mutat. 2005, 25, 410. [Google Scholar] [CrossRef]
- Finch, A.J.; Hilcenko, C.; Basse, N.; Drynan, L.F.; Goyenechea, B.; Menne, T.F.; Gonzalez Fernandez, A.; Simpson, P.; D’Santos, C.S.; Arends, M.J.; et al. Uncoupling of GTP hydrolysis from eIF6 release on the ribosome causes Shwachman-Diamond syndrome. Genes Dev. 2011, 25, 917–929. [Google Scholar] [CrossRef] [Green Version]
- Ng, C.L.; Waterman, D.G.; Koonin, E.V.; Walters, A.D.; Chong, J.P.J.; Isupov, M.N.; Lebedev, A.A.; Bunka, D.H.J.; Stockley, P.G.; Ortiz-Lombardía, M.; et al. Conformational flexibility and molecular interactions of an archaeal homologue of the Shwachman-Bodian-Diamond syndrome protein. BMC Struct. Biol. 2009, 9, 32. [Google Scholar] [CrossRef]
- Savchenko, A.; Krogan, N.; Cort, J.R.; Evdokimova, E.; Lew, J.M.; Yee, A.A.; Sanchez-Pulido, L.; Andrade, M.A.; Bochkarev, A.; Watson, J.D.; et al. The Shwachman-Bodian-Diamond syndrome protein family is involved in RNA metabolism. J. Biol. Chem. 2005, 280, 19213–19220. [Google Scholar] [CrossRef]
- Dhanraj, S.; Matveev, A.; Li, H.; Lauhasurayotin, S.; Jardine, L.; Cada, M.; Zlateska, B.; Tailor, C.S.; Zhou, J.; Mendoza-Londono, R.; et al. Biallelic mutations in DNAJC21 cause Shwachman-Diamond syndrome. Blood 2017, 129, 1557–1562. [Google Scholar] [CrossRef] [PubMed]
- Tummala, H.; Walne, A.J.; Williams, M.; Bockett, N.; Collopy, L.; Cardoso, S.; Ellison, A.; Wynn, R.; Leblanc, T.; Fitzgibbon, J.; et al. DNAJC21 Mutations Link a Cancer-Prone Bone Marrow Failure Syndrome to Corruption in 60S Ribosome Subunit Maturation. Am. J. Hum. Genet. 2016, 99, 115–124. [Google Scholar] [CrossRef]
- Carapito, R.; Konantz, M.; Paillard, C.; Miao, Z.; Pichot, A.; Leduc, M.S.; Yang, Y.; Bergstrom, K.L.; Mahoney, D.H.; Shardy, D.L.; et al. Mutations in signal recognition particle SRP54 cause syndromic neutropenia with Shwachman-Diamond-like features. J. Clin. Investig. 2017, 127, 4090–4103. [Google Scholar] [CrossRef] [PubMed]
- Stepensky, P.; Chacon-Flores, M.; Kim, K.H.; Abuzaitoun, O.; Bautista-Santos, A.; Simanovsky, N.; Siliqi, D.; Altamura, D.; Mendez-Godoy, A.; Gijsbers, A.; et al. Mutations in EFL1, an SBDS partner, are associated with infantile pancytopenia, exocrine pancreatic insufficiency and skeletal anomalies in a Shwachman-Diamond like syndrome. J. Med. Genet. 2017, 54, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.K.; Cope, H.; Spillmann, R.C.; Stong, N.; Jiang, Y.H.; McDonald, M.T.; Rothman, J.A.; Butler, M.W.; Frush, D.P.; Lachman, R.S.; et al. Further evidence for the involvement of EFL1 in a Shwachman-Diamond-like syndrome and expansion of the phenotypic features. Cold Spring Harb. Mol. Case Stud. 2018, 4, a003046. [Google Scholar] [CrossRef] [PubMed]
- Menne, T.F.; Goyenechea, B.; Sanchez-Puig, N.; Wong, C.C.; Tonkin, L.M.; Ancliff, P.J.; Brost, R.L.; Costanzo, M.; Boone, C.; Warren, A.J. The Shwachman-Bodian-Diamond syndrome protein mediates translational activation of ribosomes in yeast. Nat. Genet. 2007, 39, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Ceci, M.; Gaviraghi, C.; Gorrini, C.; Sala, L.A.; Offenhauser, N.; Marchisio, P.C.; Biffo, S. Release of eIF6 (p27BBP) from the 60S subunit allows 80S ribosome assembly. Nature 2003, 426, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Marquez, A.; Gijsbers, A.; de la Mora, E.; Sanchez-Puig, N. Defective Guanine Nucleotide Exchange in the Elongation Factor-like 1 (EFL1) GTPase by Mutations in the Shwachman-Diamond Syndrome Protein. J. Biol. Chem. 2015, 290, 17669–17678. [Google Scholar] [CrossRef] [PubMed]
- Gijsbers, A.; Garcia-Marquez, A.; Luviano, A.; Sanchez-Puig, N. Guanine nucleotide exchange in the ribosomal GTPase EFL1 is modulated by the protein mutated in the Shwachman-Diamond Syndrome. Biochem. Biophys. Res. Commun. 2013, 437, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Calamita, P.; Miluzio, A.; Russo, A.; Pesce, E.; Ricciardi, S.; Khanim, F.; Cheroni, C.; Alfieri, R.; Mancino, M.; Gorrini, C.; et al. SBDS-Deficient Cells Have an Altered Homeostatic Equilibrium due to Translational Inefficiency Which Explains their Reduced Fitness and Provides a Logical Framework for Intervention. PLoS Genet. 2017, 13, e1006552. [Google Scholar] [CrossRef] [PubMed]
- Gijsbers, A.; Nishigaki, T.; Sanchez-Puig, N. Fluorescence Anisotropy as a Tool to Study Protein-protein Interactions. J. Vis. Exp. 2016. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, J.F.; Sforca, M.L.; Blumenschein, T.M.A.; Goldfeder, M.B.; Guimaraes, B.G.; Oliveira, C.C.; Zanchin, N.I.T.; Zeri, A.-C. Structure, dynamics, and RNA interaction analysis of the human SBDS protein. J. Mol. Biol. 2010, 396, 1053–1069. [Google Scholar] [CrossRef] [PubMed]
- Siliqi, D.; Foadi, J.; Mazzorana, M.; Altamura, D.; Méndez-Godoy, A.; Sánchez-Puig, N. Conformational flexibility of proteins involved in ribosome biogenesis: Investogations via small angle X-ray scattering (SAXS). Crystals 2018, 8, 109. [Google Scholar] [CrossRef]
- Jorgensen, R.; Ortiz, P.A.; Carr-Schmid, A.; Nissen, P.; Kinzy, T.G.; Andersen, G.R. Two crystal structures demonstrate large conformational changes in the eukaryotic ribosomal translocase. Nat. Struct. Biol. 2003, 10, 379–385. [Google Scholar] [CrossRef]
- Weis, F.; Giudice, E.; Churcher, M.; Jin, L.; Hilcenko, C.; Wong, C.C.; Traynor, D.; Kay, R.R.; Warren, A.J. Mechanism of eIF6 release from the nascent 60S ribosomal subunit. Nat. Struct. Mol. Biol. 2015, 22, 914–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneidman-Duhovny, D.; Hammel, M.; Tainer, J.A.; Sali, A. FoXS, FoXSDock and MultiFoXS: Single-state and multi-state structural modeling of proteins and their complexes based on SAXS profiles. Nucleic Acids Res. 2016, 44, W424–W429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez-Garcia, B.; Pons, C.; Svergun, D.I.; Bernado, P.; Fernandez-Recio, J. pyDockSAXS: Protein-protein complex structure by SAXS and computational docking. Nucleic Acids Res. 2015, 43, W356–W361. [Google Scholar] [CrossRef]
- Woloszynek, J.R.; Rothbaum, R.J.; Rawls, A.S.; Minx, P.J.; Wilson, R.K.; Mason, P.J.; Bessler, M.; Link, D.C. Mutations of the SBDS gene are present in most patients with Shwachman-Diamond syndrome. Blood 2004, 104, 3588–3590. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, C.M.; Zuccherato, L.W.; Williams, C.L.; Neill, N.J.; Murdock, D.R.; Bainbridge, M.; Jhangiani, S.N.; Muzny, D.M.; Gibbs, R.A.; Ip, W.; et al. Structural variation and missense mutation in SBDS associated with Shwachman-Diamond syndrome. BMC Med. Genet. 2014, 15, 64. [Google Scholar] [CrossRef]
- Erdos, M.; Alapi, K.; Balogh, I.; Oroszlan, G.; Rakoczi, E.; Sumegi, J.; Marodi, L. Severe Shwachman-Diamond syndrome phenotype caused by compound heterozygous missense mutations in the SBDS gene. Exp. Hematol. 2006, 34, 1517–1521. [Google Scholar] [CrossRef]
- Holding, A.N. XL-MS: Protein cross-linking coupled with mass spectrometry. Methods 2015, 89, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Austin, K.M.; Leary, R.J.; Shimamura, A. The Shwachman-Diamond SBDS protein localizes to the nucleolus. Blood 2005, 106, 1253–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luz, J.S.; Georg, R.C.; Gomes, C.H.; Machado-Santelli, G.M.; Oliveira, C.C. Sdo1p, the yeast orthologue of Shwachman-Bodian-Diamond syndrome protein, binds RNA and interacts with nuclear rRNA-processing factors. Yeast 2009, 26, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Gandin, V.; Miluzio, A.; Barbieri, A.M.; Beugnet, A.; Kiyokawa, H.; Marchisio, P.C.; Biffo, S. Eukaryotic initiation factor 6 is rate-limiting in translation, growth and transformation. Nature 2008, 455, 684–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Record, M.T., Jr.; Anderson, C.F.; Mills, P.; Mossing, M.; Roe, J.H. Ions as regulators of protein-nucleic acid interactions in vitro and in vivo. Adv. Biophys. 1985, 20, 109–135. [Google Scholar] [CrossRef]
- Record, M.T., Jr.; Ha, J.H.; Fisher, M.A. Analysis of equilibrium and kinetic measurements to determine thermodynamic origins of stability and specificity and mechanism of formation of site-specific complexes between proteins and helical DNA. Methods Enzymol. 1991, 208, 291–343. [Google Scholar] [PubMed]
- Mitkevich, V.A.; Shyp, V.; Petrushanko, I.Y.; Soosaar, A.; Atkinson, G.C.; Tenson, T.; Makarov, A.A.; Hauryliuk, V. GTPases IF2 and EF-G bind GDP and the SRL RNA in a mutually exclusive manner. Sci. Rep. 2012, 2, 843. [Google Scholar] [CrossRef] [PubMed]
- Whitty, A. Cooperativity and biological complexity. Nat. Chem. Biol. 2008, 4, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Asano, N.; Atsuumi, H.; Nakamura, A.; Tanaka, Y.; Tanaka, I.; Yao, M. Direct interaction between EFL1 and SBDS is mediated by an intrinsically disordered insertion domain. Biochem. Biophys. Res. Commun. 2014, 443, 1251–1256. [Google Scholar] [CrossRef] [Green Version]
- Blanchet, C.E.; Spilotros, A.; Schwemmer, F.; Graewert, M.A.; Kikhney, A.; Jeffries, C.M.; Franke, D.; Mark, D.; Zengerle, R.; Cipriani, F.; et al. Versatile sample environments and automation for biological solution X-ray scattering experiments at the P12 beamline (PETRA III, DESY). J. Appl. Crystallogr. 2015, 48, 431–443. [Google Scholar] [CrossRef]
- Graewert, M.A.; Franke, D.; Jeffries, C.M.; Blanchet, C.E.; Ruskule, D.; Kuhle, K.; Flieger, A.; Schafer, B.; Tartsch, B.; Meijers, R.; et al. Automated pipeline for purification, biophysical and x-ray analysis of biomacromolecular solutions. Sci. Rep. 2015, 5, 10734. [Google Scholar] [CrossRef]
- Franke, D.; Kikhney, A.G.; Svergun, D.I. Automated acquisition and analysis of small angle X-ray scattering data. Nucl. Instrum. Methods Phys. Res. Sect. A 2012, 689, 52–59. [Google Scholar] [CrossRef]
- Guinier, A. La diffraction des rayons X aux très petits angles: Application à l’étude de phénomènes ultramicroscopiques. Ann. Phys. 1939, 11, 161–237. [Google Scholar] [CrossRef]
- Glatter, O. A new method for the evaluation of small-angle scattering data. J. Appl. Crystallogr. 1977, 10, 415–421. [Google Scholar] [CrossRef] [Green Version]
- Svergun, D.I.; Petoukhov, M.V.; Koch, M.H. Determination of domain structure of proteins from X-ray solution scattering. Biophys. J. 2001, 80, 2946–2953. [Google Scholar] [CrossRef]
- Volkov, V.V.; Svergun, D.I. Uniqueness of ab initio shape determination in small-angle scattering. J. Appl. Crystallogr. 2003, 36, 860–864. [Google Scholar] [CrossRef]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 40. [Google Scholar] [CrossRef]
- Emekli, U.; Schneidman-Duhovny, D.; Wolfson, H.J.; Nussinov, R.; Haliloglu, T. HingeProt: Automated prediction of hinges in protein structures. Proteins 2008, 70, 1219–1227. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SBDS Missense Variants | |||
| SBDS | Kd1 (µM) | Kd2 (µM) | Hill coefficient |
Wild type | 5.4 ± 1 | 0.4 ± 0.04 | 1.8 ± 0.02 |
R19Q | 4.2 ± 0.5 | 5.2 ± 0.3 | 1.5 ± 0.02 |
K67E | 7.2 ± 2 | 3 ± 1 | 1.8 ± 0.07 |
R126T | 9.3 ± 1.7 | 2.5 ± 0.2 | 1.7 ± 0.04 |
S143L | ND | ND | ND |
I167T | 10 ± 3 | 1.4 ± 0.2 | 1.5 ± 0.04 |
R175W | 12 ± 4 | 3.3 ± 0.6 | 1.3 ± 0.03 |
I212T | ND | ND | ND |
| SBDS Constructs | |||
| SBDS | Kd1 (µM) | Kd2 (µM) | Hill coefficient |
 | 15.2 ± 0.72 | - | - |
Domain 2 * | 10.8 ± 0.9 | - | - |
 | 11.3 ± 1.8 | 3.9 ± 0.2 | 1.4 ± 0.02 |
 | 4.6 ± 0.3 | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gijsbers, A.; Montagut, D.C.; Méndez-Godoy, A.; Altamura, D.; Saviano, M.; Siliqi, D.; Sánchez-Puig, N. Interaction of the GTPase Elongation Factor Like-1 with the Shwachman-Diamond Syndrome Protein and Its Missense Mutations. Int. J. Mol. Sci. 2018, 19, 4012. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124012
Gijsbers A, Montagut DC, Méndez-Godoy A, Altamura D, Saviano M, Siliqi D, Sánchez-Puig N. Interaction of the GTPase Elongation Factor Like-1 with the Shwachman-Diamond Syndrome Protein and Its Missense Mutations. International Journal of Molecular Sciences. 2018; 19(12):4012. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124012
Chicago/Turabian StyleGijsbers, Abril, Diana Carolina Montagut, Alfonso Méndez-Godoy, Davide Altamura, Michele Saviano, Dritan Siliqi, and Nuria Sánchez-Puig. 2018. "Interaction of the GTPase Elongation Factor Like-1 with the Shwachman-Diamond Syndrome Protein and Its Missense Mutations" International Journal of Molecular Sciences 19, no. 12: 4012. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124012