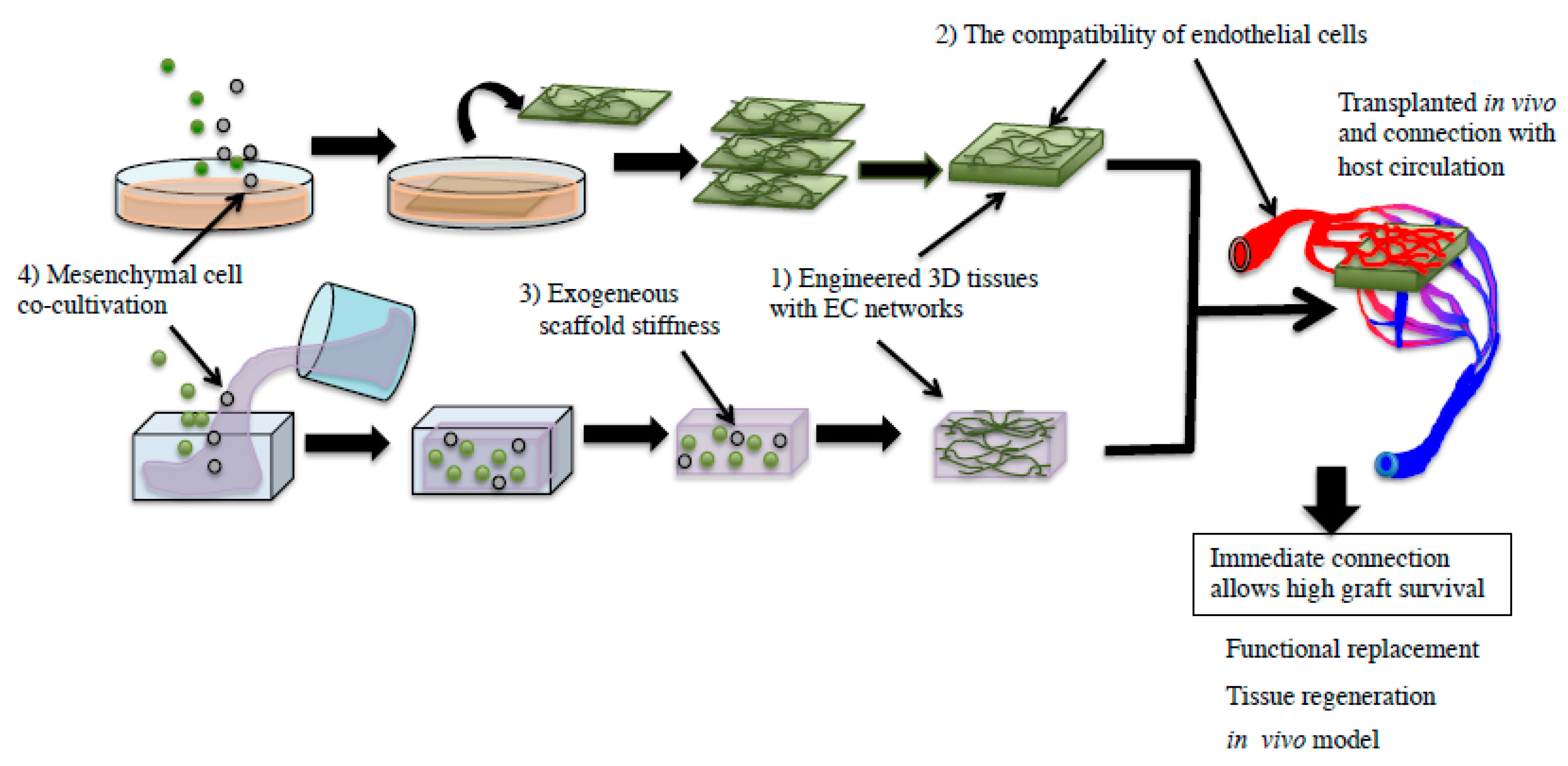
Pathological Process of Prompt Connection between Host and Donor Tissue Vasculature Causing Rapid Perfusion of the Engineered Donor Tissue after Transplantation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Factors Affecting Host–Donor Vasculature Connection Causing Donor Tissue Perfusion
3. Establishment of Blood Perfusion in Engineered Tissues after Transplantation
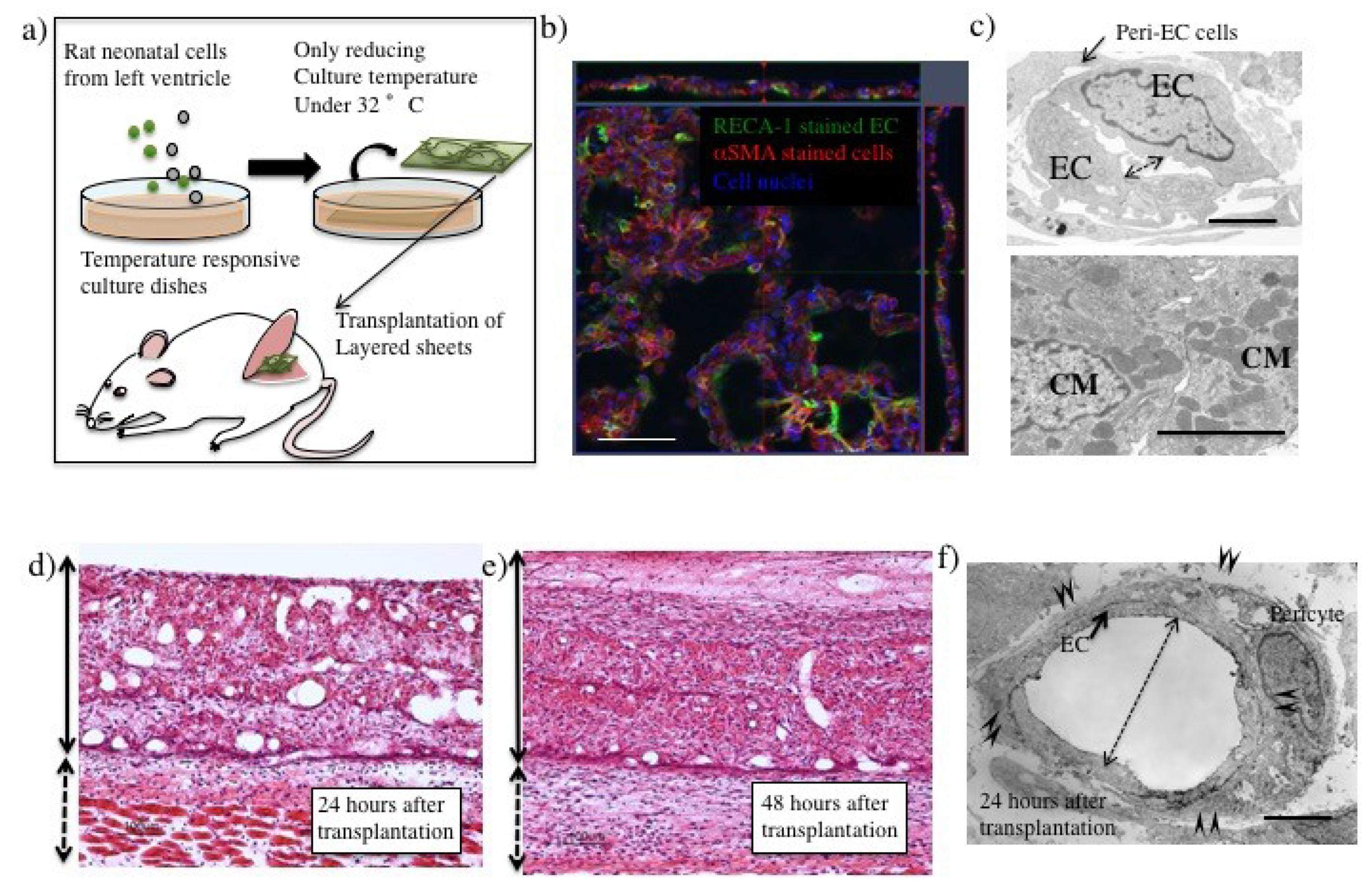
3.1. Tissue Engineering Technology Including Cell Sheet Technology
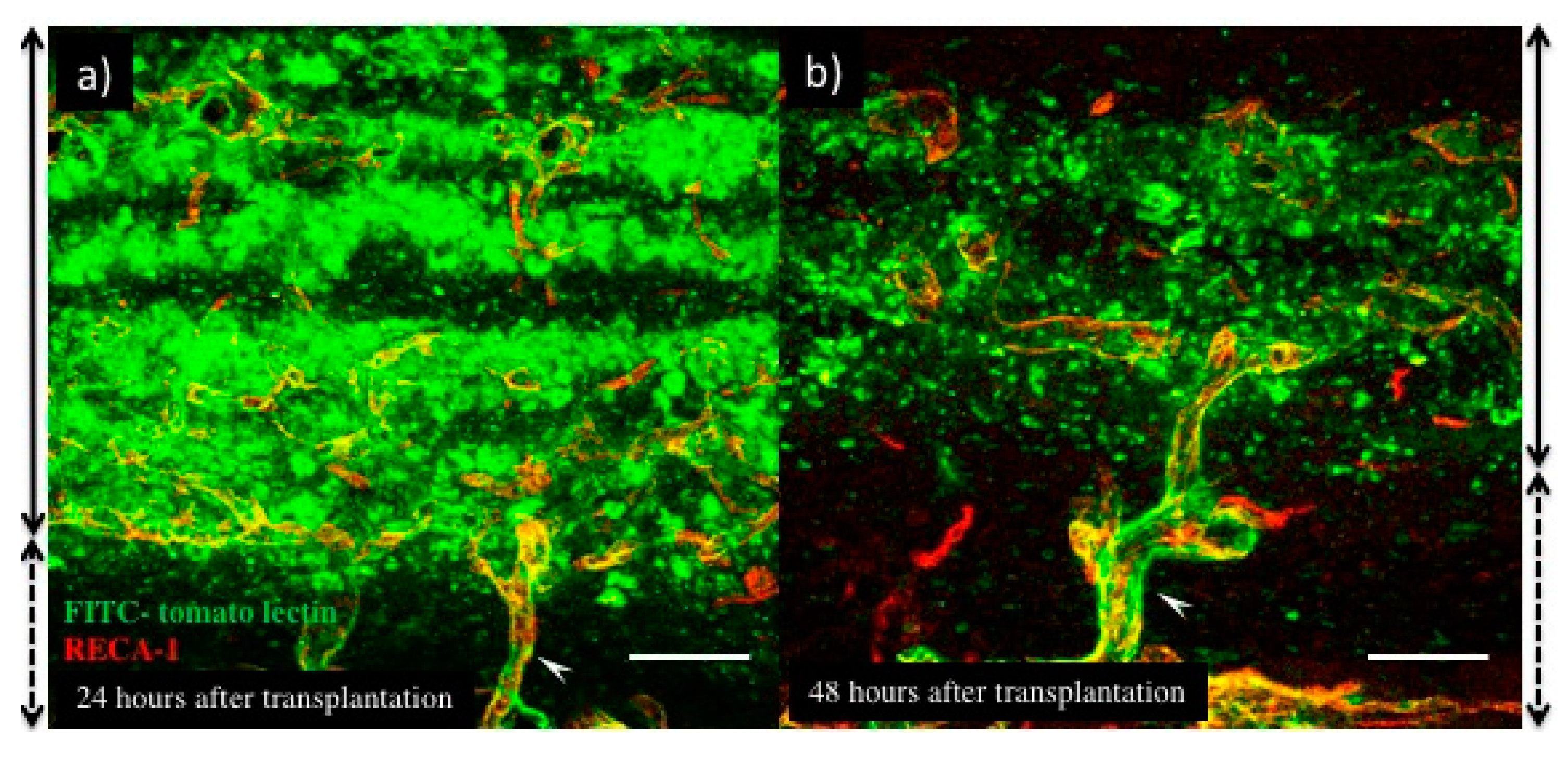
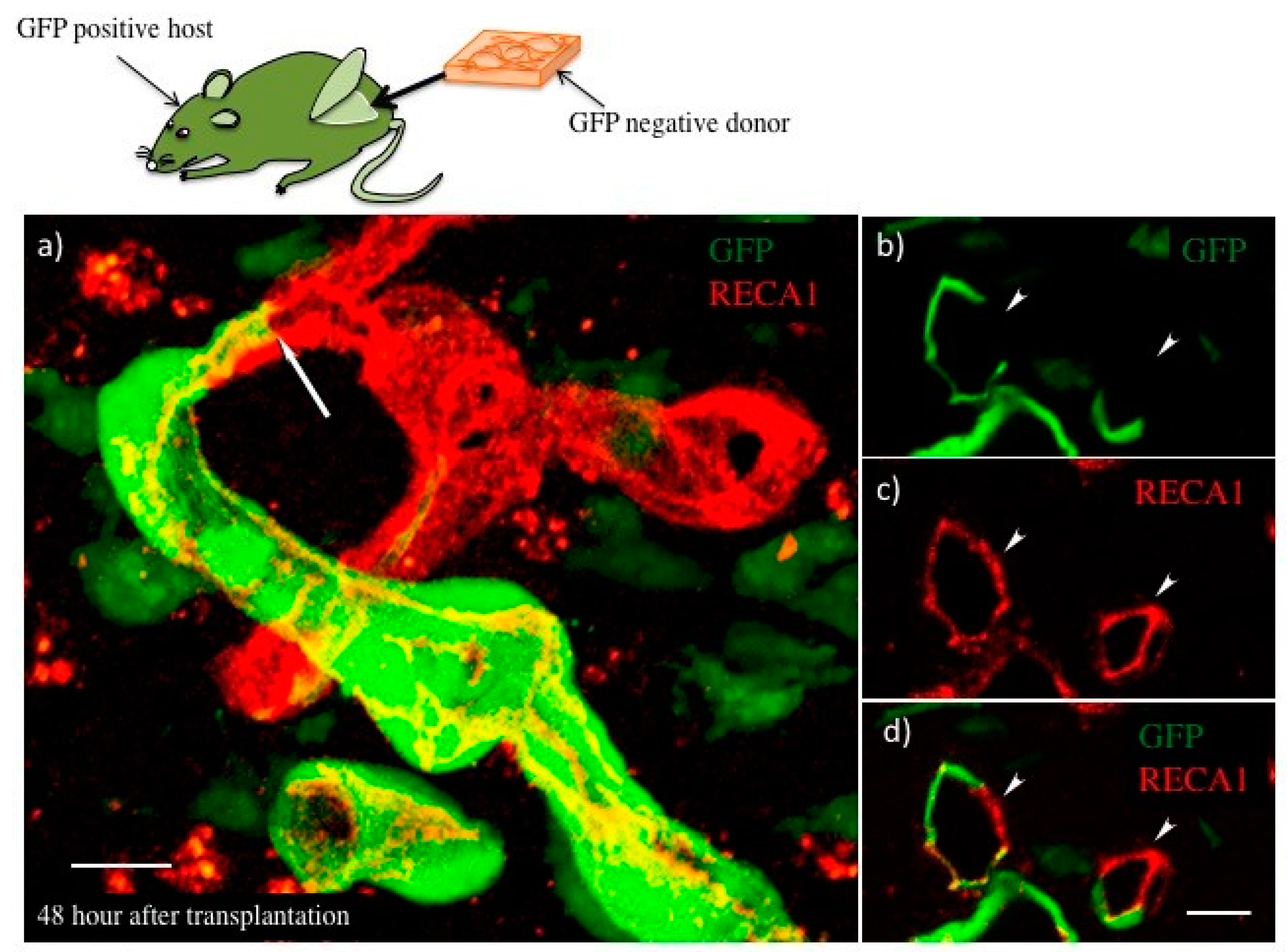
3.2. Blood Perfusion throughout Engineered Tissue after Transplantation
3.3. The Process of Host–Donor Vasculature Connection after Transplantation
4. Conclusions
5. Materials and Methods
5.1. Preparation of Myocardial Cell Sheet
5.2. Transplantation of Fabricated Tissues into Rats
5.3. Vessel Permeability Analysis (FITC-Conjugated Tomato Lectin Perfusion)
5.4. Tissue Preparation for Histological Analysis
5.5. Immunohistochemistry
5.6. Transmission Electron Microscopy
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| VEGF | vascular endothelial growth factor |
| NG2 | neural/glial antigen 2 |
| ECM | extracellular matrix |
| GFP | green fluorescent protein |
| PDGFR | platelet-derived growth factor receptor |
| FITC | fluorescein isothiocyanate |
| RECA-1 | rat endothelial cell antigen-1 |
| NG2+ PDGFRβ+ | NG2 positive and PDGF-β positive |
References
- Reich, H.J.; Kobashigawa, J.A.; Aintablian, T.; Ramzy, D.; Kittleson, M.M.; Esmailian, F. Effects of older donor age and cold ischemic time on long-term outcomes of heart transplantation. Tex. Heart Instit. J. 2018, 45, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Postalcioglu, M.; Kaze, A.D.; Byun, B.C.; Siedlecki, A.; Tullius, S.G.; Milford, E.L.; Paik, J.M.; Abdi, R. Association of cold ischemia time with acute renal transplant rejection. Transplantation 2018, 102, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Sato, H.; Kato-Itoh, M.; Goto, T.; Hara, H.; Sanbo, M.; Mizuno, N.; Kobayashi, T.; Yanagida, A.; Umino, A.; et al. Interspecies organogenesis generates autologous functional islets. Nature 2017, 542, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Takebe, T.; Sekine, K.; Enomura, M.; Koike, H.; Kimura, M.; Ogaeri, T.; Zhang, R.R.; Ueno, Y.; Zheng, Y.W.; Koike, N.; et al. Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature 2013, 499, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Song, J.J.; Guyette, J.P.; Gilpin, S.E.; Gonzalez, G.; Vacanti, J.P.; Ott, H.C. Regeneration and experimental orthotopic transplantation of a bioengineered kidney. Nat. Med. 2013, 19, 646–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekiya, S.; Shimizu, T. Introduction of vasculature in engineered three-dimensional tissue. Inflamm. Regen. 2017, 37, 25. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Liao, S.; Kit Wong, H.; Lacorre, D.A.; di Tomaso, E.; Au, P.; Fukumura, D.; Jain, R.K.; Munn, L.L. Engineered blood vessel networks connect to host vasculature via wrapping-and-tapping anastomosis. Blood 2011, 118, 4740–4749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekiya, S.; Shimizu, T.; Yamato, M.; Kikuchi, A.; Okano, T. Bioengineered cardiac cell sheet grafts have intrinsic angiogenic potential. Biochem. Biophys. Res. Commun. 2006, 341, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Sasagawa, T.; Shimizu, T.; Sekiya, S.; Yamato, M.; Okano, T. Comparison of angiogenic potential between prevascular and non-prevascular layered adipose-derived stem cell-sheets in early post-transplanted period. J. Biomed Mater. Res. A 2014, 102, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Asano, Y.; Shimoda, H.; Okano, D.; Matsusaki, M.; Akashi, M. Transplantation of three-dimensional artificial human vascular tissues fabricated using an extracellular matrix nanofilm-based cell-accumulation technique. J. Tissue Eng. Regen. Med. 2017, 11, 1303–1307. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, M.D.; Piristine, H.; Hogrebe, N.J.; Nocera, T.M.; Boehm, M.W.; Reen, R.K.; Koelling, K.W.; Agarwal, G.; Sarang-Sieminski, A.L.; Gooch, K.J. A self-assembling peptide matrix used to control stiffness and binding site density supports the formation of microvascular networks in three dimensions. Acta Biomater. 2013, 9, 7651–7661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shkumatov, A.; Thompson, M.; Choi, K.M.; Sicard, D.; Baek, K.; Kim, D.H.; Tschumperlin, D.J.; Prakash, Y.S.; Kong, H. Matrix stiffness-modulated proliferation and secretory function of the airway smooth muscle cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 308, L1125–L1135. [Google Scholar] [CrossRef] [PubMed]
- Koike, N.; Fukumura, D.; Gralla, O.; Au, P.; Schechner, J.S.; Jain, R.K. Tissue engineering: Creation of long-lasting blood vessels. Nature 2004, 428, 138–139. [Google Scholar] [CrossRef] [PubMed]
- Au, P.; Tam, J.; Fukumura, D.; Jain, R.K. Bone marrow-derived mesenchymal stem cells facilitate engineering of long-lasting functional vasculature. Blood 2008, 111, 4551–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, T. Cell sheet-based tissue engineering for fabricating 3-dimensional heart tissues. Circ. J. 2014, 78, 2594–2603. [Google Scholar] [CrossRef] [PubMed]
- Matsusaki, M.; Case, C.P.; Akashi, M. Three-dimensional cell culture technique and pathophysiology. Adv. Drug Deliv. Rev. 2014, 74, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekiya, S.; Shimizu, T.; Okano, T. Vascularization in 3D tissue using cell sheet technology. Regen. Med. 2013, 8, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Ohki, T.; Yamato, M.; Ota, M.; Takagi, R.; Kondo, M.; Kanai, N.; Okano, T.; Yamamoto, M. Application of regenerative medical technology using tissue-engineered cell sheets for endoscopic submucosal dissection of esophageal neoplasms. Dig. Endosc. 2015, 27, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Ilan, N.; Mahooti, S.; Madri, J.A. Distinct signal transduction pathways are utilized during the tube formation and survival phases of in vitro angiogenesis. J. Cell Sci. 1998, 111, 3621–3631. [Google Scholar] [PubMed]
- Montesano, R.; Pepper, M.S.; Orci, L. Paracrine induction of angiogenesis in vitro by swiss 3T3 fibroblasts. J. Cell Sci. 1993, 105, 1013–1024. [Google Scholar] [PubMed]
- Asakawa, N.; Shimizu, T.; Tsuda, Y.; Sekiya, S.; Sasagawa, T.; Yamato, M.; Fukai, F.; Okano, T. Pre-vascularization of in vitro three-dimensional tissues created by cell sheet engineering. Biomaterials 2010, 31, 3903–3909. [Google Scholar] [CrossRef] [PubMed]
- Burri, P.H.; Hlushchuk, R.; Djonov, V. Intussusceptive angiogenesis: Its emergence, its characteristics, and its significance. Dev. Dyn. 2004, 231, 474–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groppa, E.; Brkic, S.; Uccelli, A.; Wirth, G.; Korpisalo-Pirinen, P.; Filippova, M.; Dasen, B.; Sacchi, V.; Muraro, M.G.; Trani, M.; et al. EphrinB2/EphB4 signaling regulates non-sprouting angiogenesis by VEGF. EMBO Rep. 2018, 19, e45054. [Google Scholar] [CrossRef] [PubMed]
- Gianni-Barrera, R.; Trani, M.; Fontanellaz, C.; Heberer, M.; Djonov, V.; Hlushchuk, R.; Banfi, A. VEGF over-expression in skeletal muscle induces angiogenesis by intussusception rather than sprouting. Angiogenesis 2013, 16, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Tsoporis, J.N.; Izhar, S.; Proteau, G.; Slaughter, G.; Parker, T.G. S100B-RAGE dependent VEGF secretion by cardiac myocytes induces myofibroblast proliferation. J. Mol. Cell. Cardiol. 2012, 52, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, S.; Muraoka, M.; Sasagawa, T.; Shimizu, T.; Yamato, M.; Okano, T. Three-dimensional cell-dense constructs containing endothelial cell-networks are an effective tool for in vivo and in vitro vascular biology research. Microvasc. Res. 2010, 80, 549–551. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, S.; Ezaki, T. Phenotypic changes and possible angiogenic roles of pericytes during wound healing in the mouse skin. Histol. Histopathol. 2011, 26, 979–995. [Google Scholar] [PubMed]
- Morikawa, S.; Baluk, P.; Kaidoh, T.; Haskell, A.; Jain, R.K.; McDonald, D.M. Abnormalities in pericytes on blood vessels and endothelial sprouts in tumors. Am. J. Pathol. 2002, 160, 985–1000. [Google Scholar] [CrossRef]
- Ruan, J.; Luo, M.; Wang, C.; Fan, L.; Yang, S.N.; Cardenas, M.; Geng, H.; Leonard, J.P.; Melnick, A.; Cerchietti, L.; et al. Imatinib disrupts lymphoma angiogenesis by targeting vascular pericytes. Blood 2013, 121, 5192–5202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, T.; Yamato, M.; Isoi, Y.; Akutsu, T.; Setomaru, T.; Abe, K.; Kikuchi, A.; Umezu, M.; Okano, T. Fabrication of pulsatile cardiac tissue grafts using a novel 3-dimensional cell sheet manipulation technique and temperature-responsive cell culture surfaces. Circ. Res. 2002, 90, e40. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sekiya, S.; Morikawa, S.; Ezaki, T.; Shimizu, T. Pathological Process of Prompt Connection between Host and Donor Tissue Vasculature Causing Rapid Perfusion of the Engineered Donor Tissue after Transplantation. Int. J. Mol. Sci. 2018, 19, 4102. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124102
Sekiya S, Morikawa S, Ezaki T, Shimizu T. Pathological Process of Prompt Connection between Host and Donor Tissue Vasculature Causing Rapid Perfusion of the Engineered Donor Tissue after Transplantation. International Journal of Molecular Sciences. 2018; 19(12):4102. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124102
Chicago/Turabian StyleSekiya, Sachiko, Shunichi Morikawa, Taichi Ezaki, and Tatsuya Shimizu. 2018. "Pathological Process of Prompt Connection between Host and Donor Tissue Vasculature Causing Rapid Perfusion of the Engineered Donor Tissue after Transplantation" International Journal of Molecular Sciences 19, no. 12: 4102. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124102