The Activity of Matrix Metalloproteinases (MMP-2, MMP-9) and Their Tissue Inhibitors (TIMP-1, TIMP-3) in the Cerebral Cortex and Hippocampus in Experimental Acanthamoebiasis

 , , , , and
, , , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
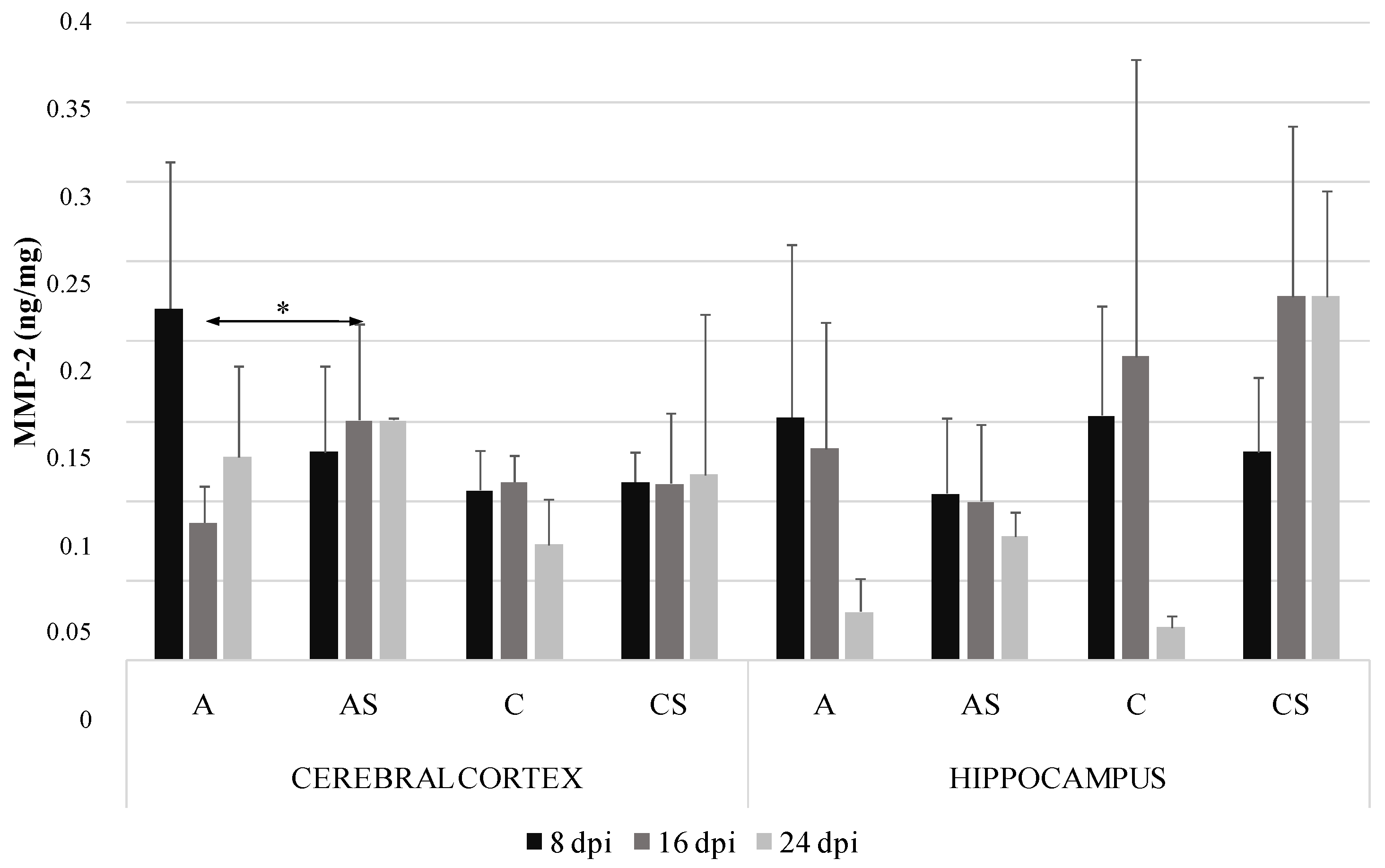
2.1. MMP-2 in the Cerebral Cortex and Hippocampus during Acanthamoebiasis
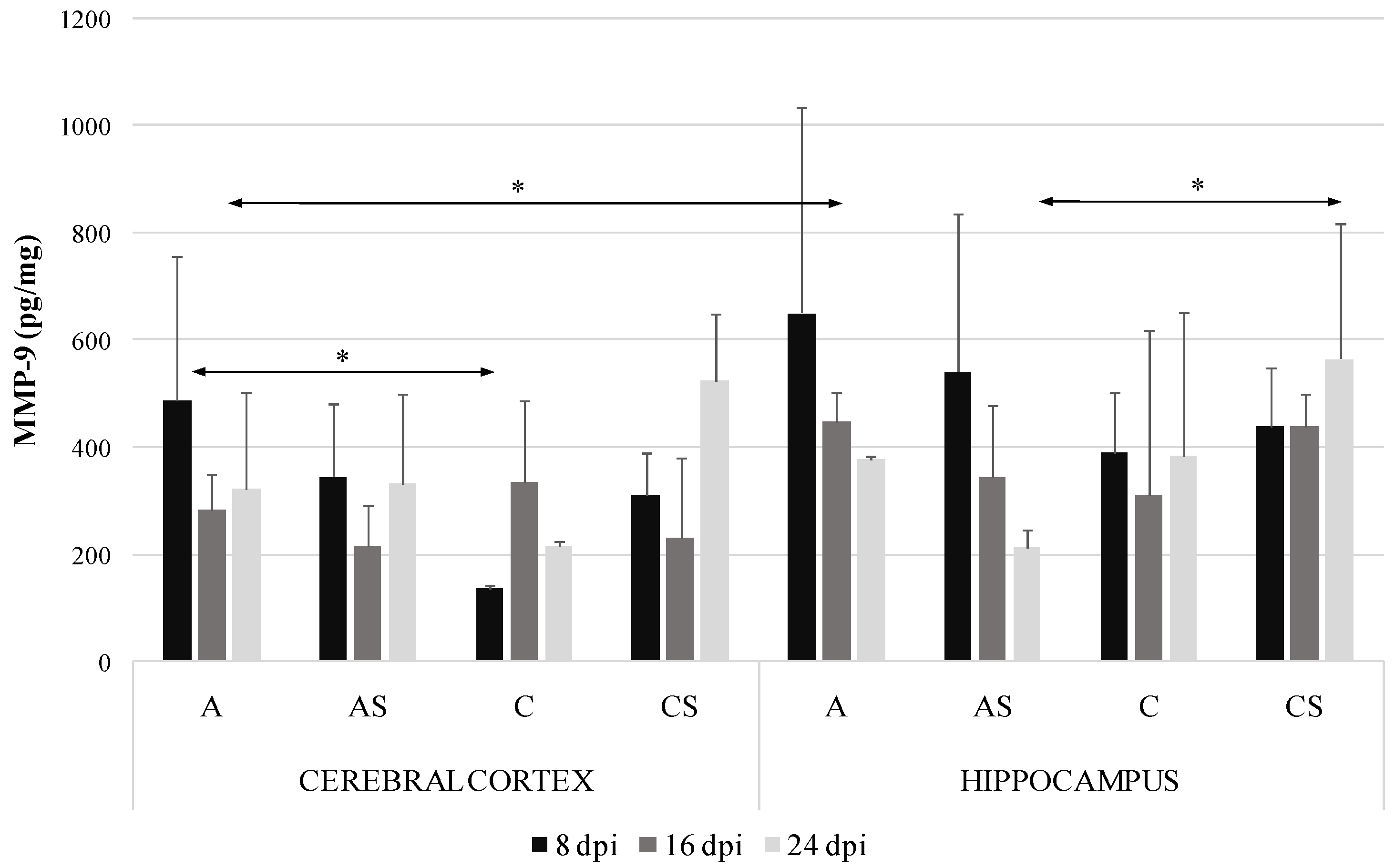
2.2. MMP-9 in the Cerebral Cortex and Hippocampus during Acanthamoebiasis
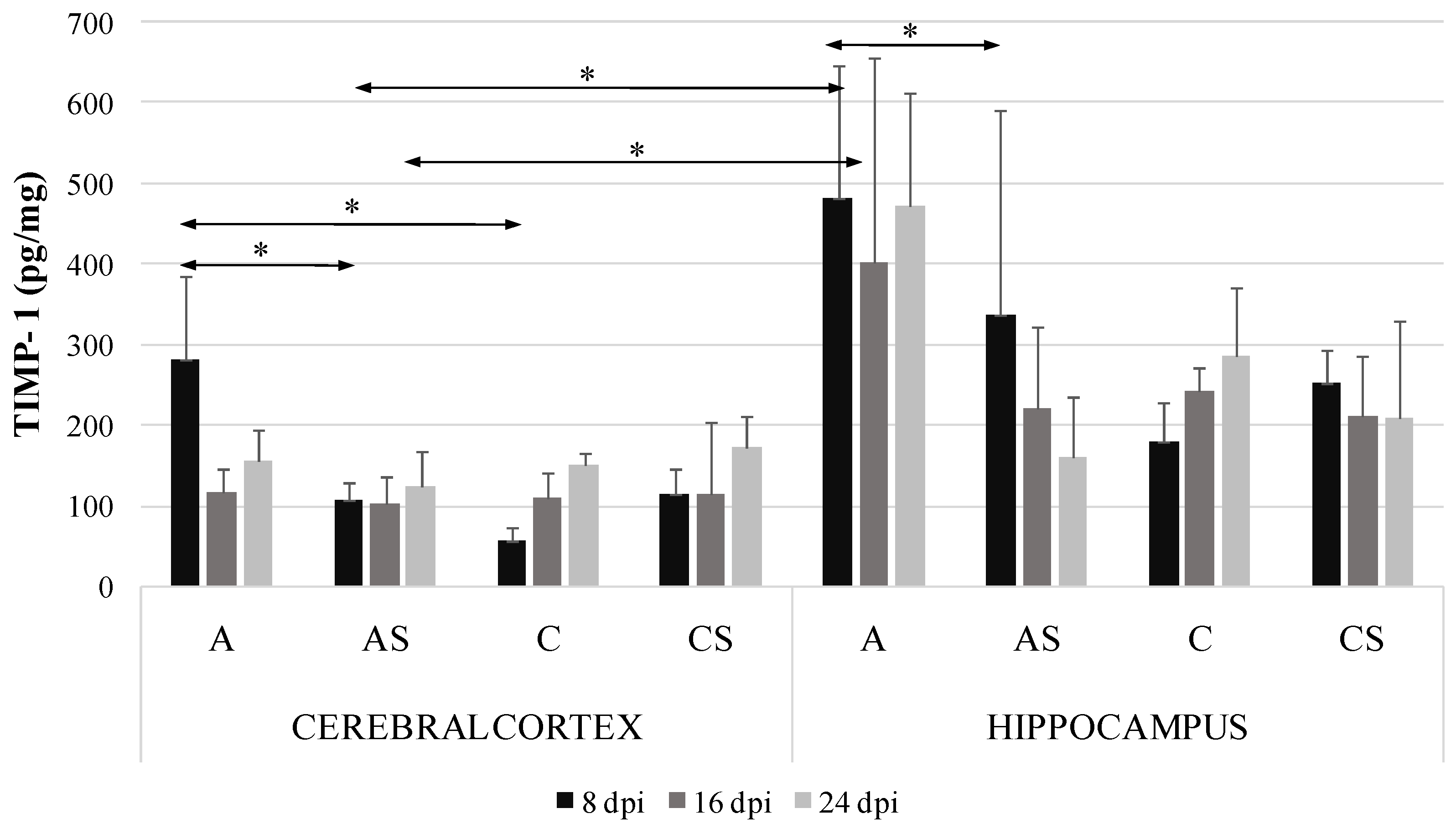
2.3. TIMP-1 in the Cerebral Cortex and Hippocampus during Acanthamoebiasis
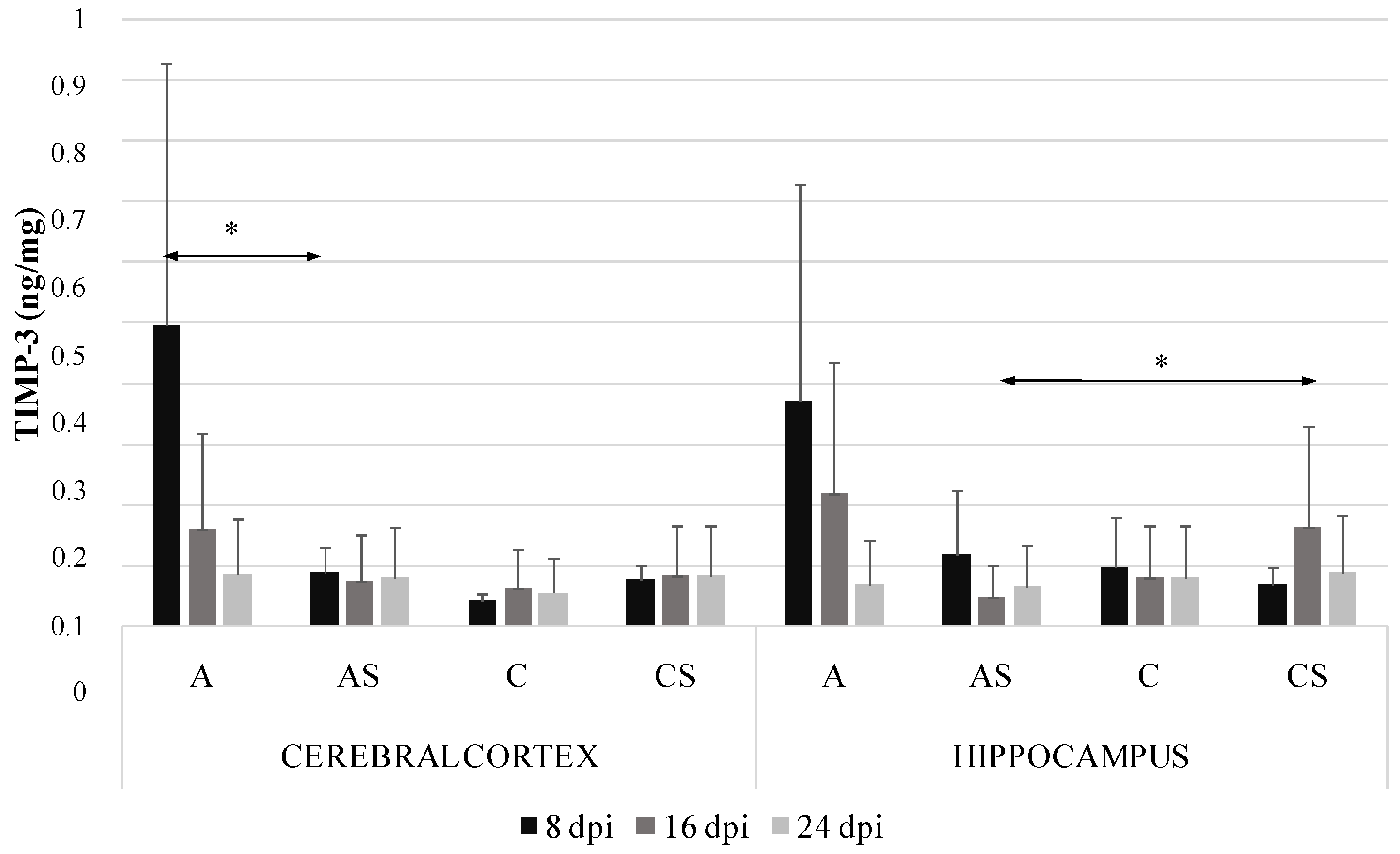
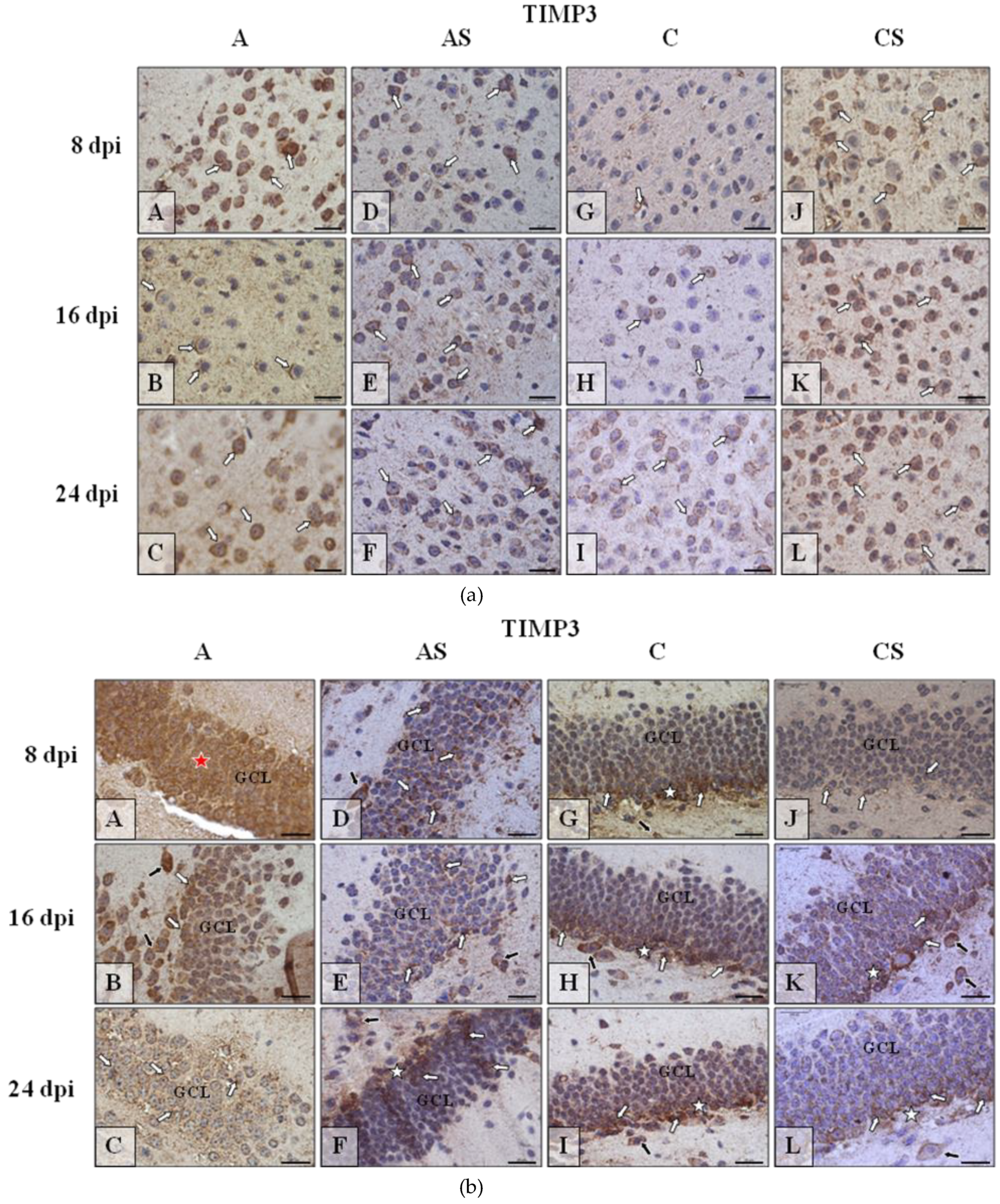
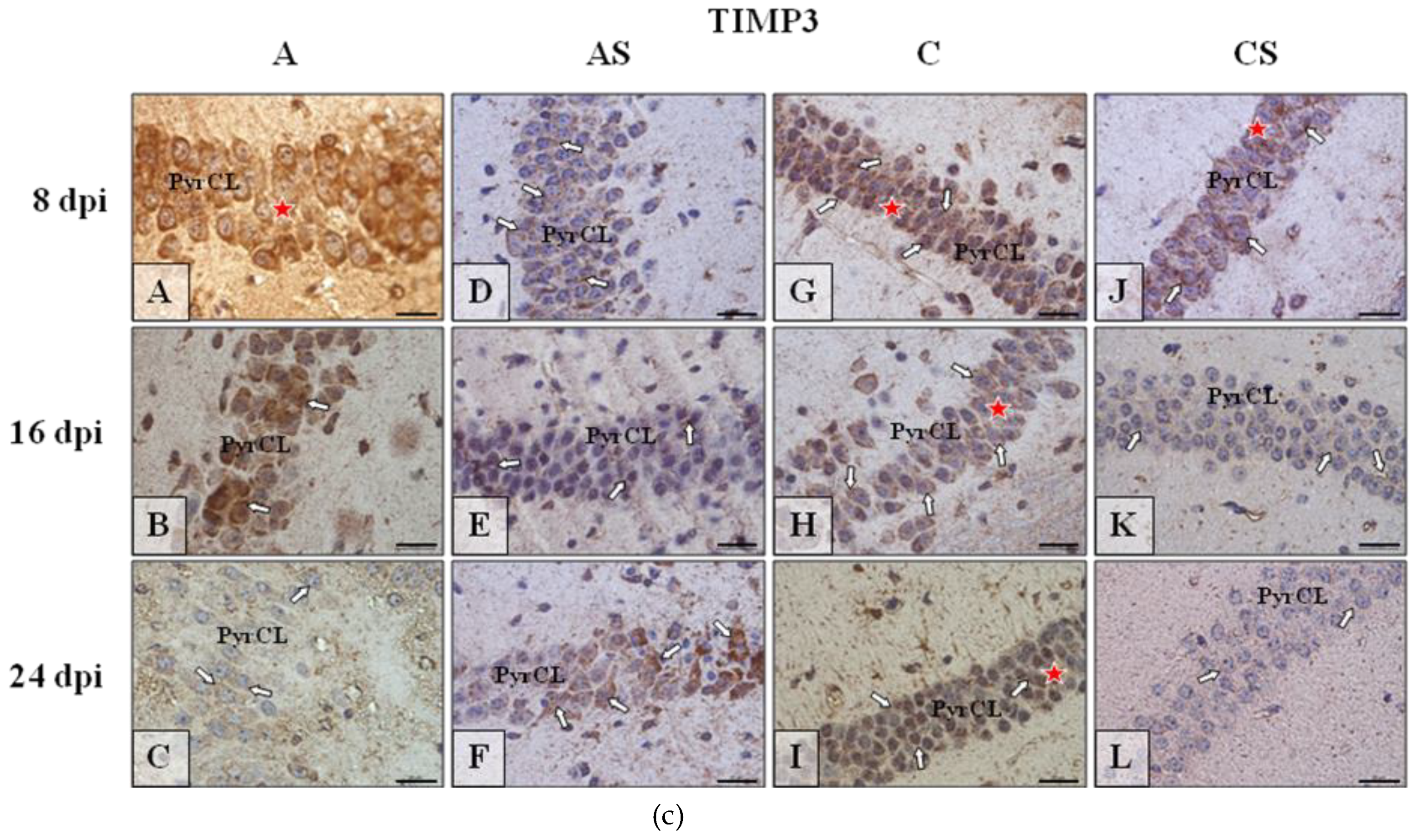
2.4. TIMP-3 in the Cerebral Cortex and Hippocampus during Acanthamoebiasis
2.5. MMP-9/TIMP-1 and MMP-2/TIMP-3 Ratios in the Brain Structures
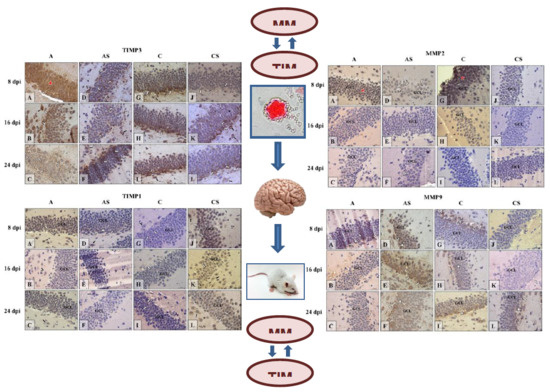
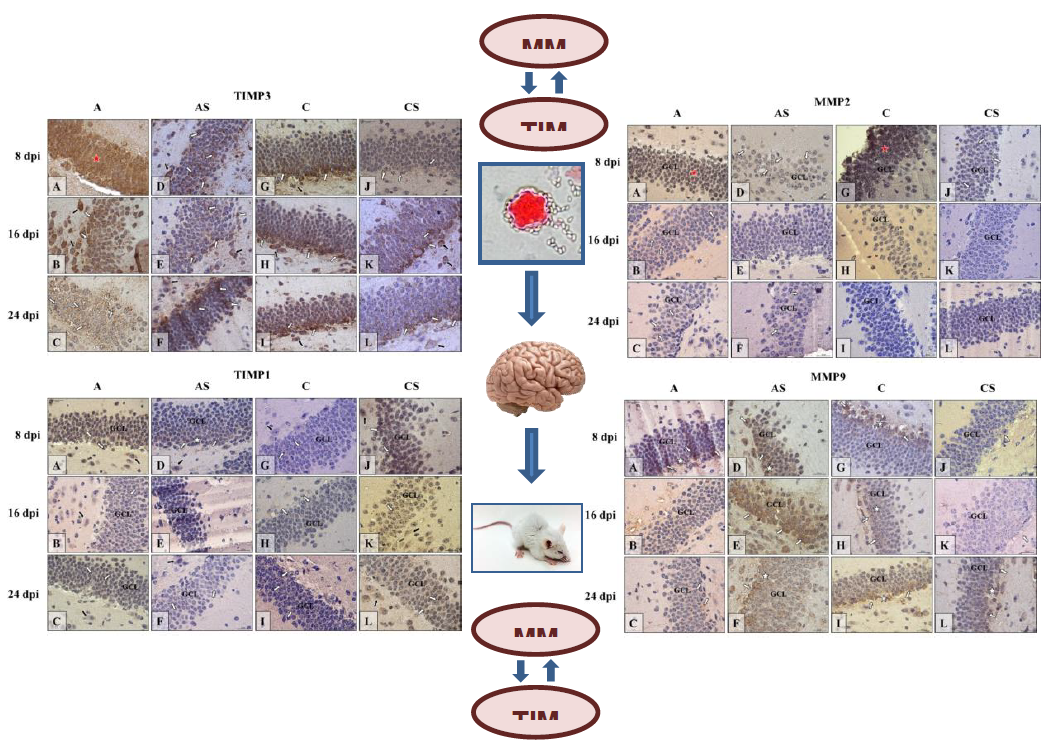
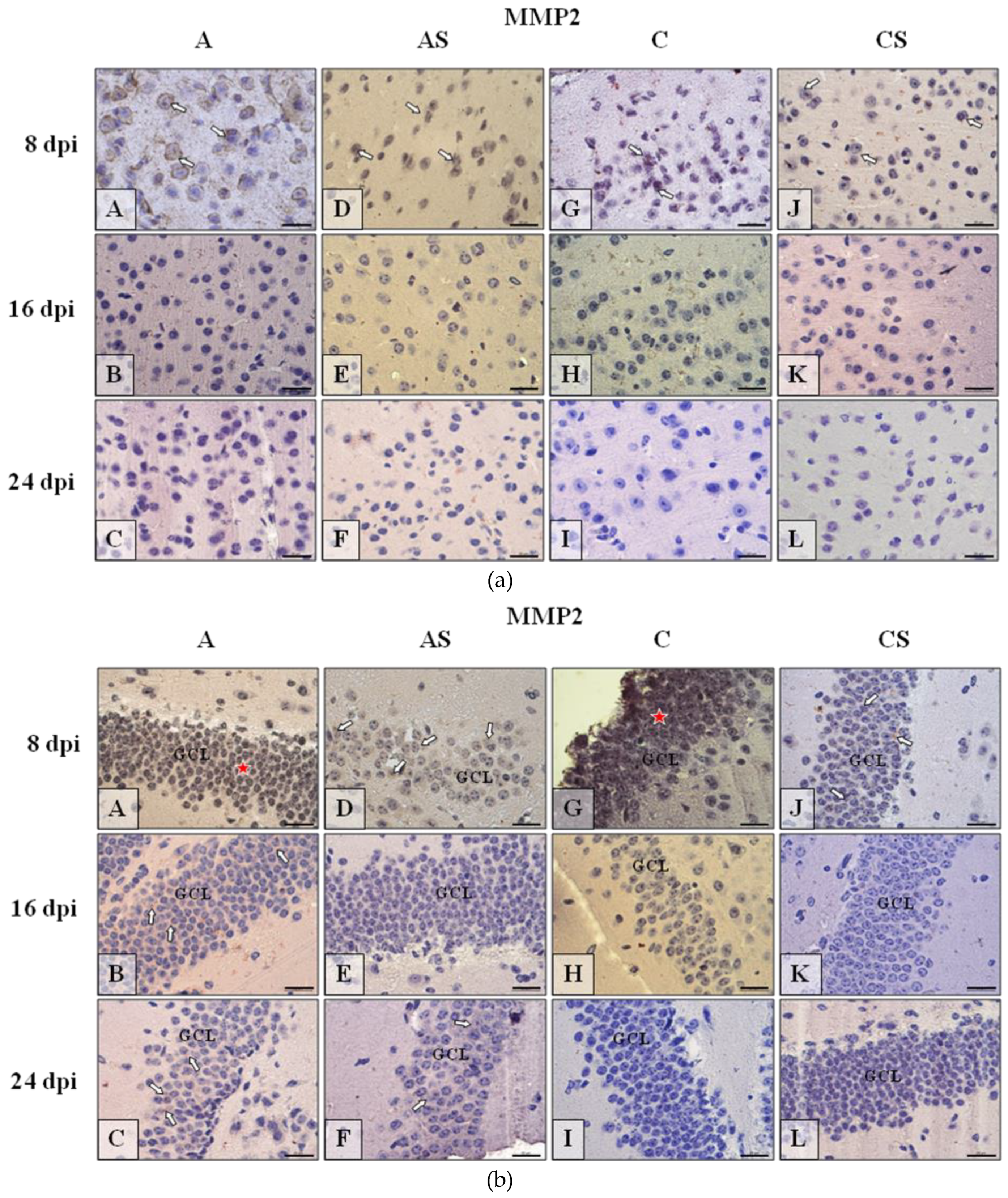
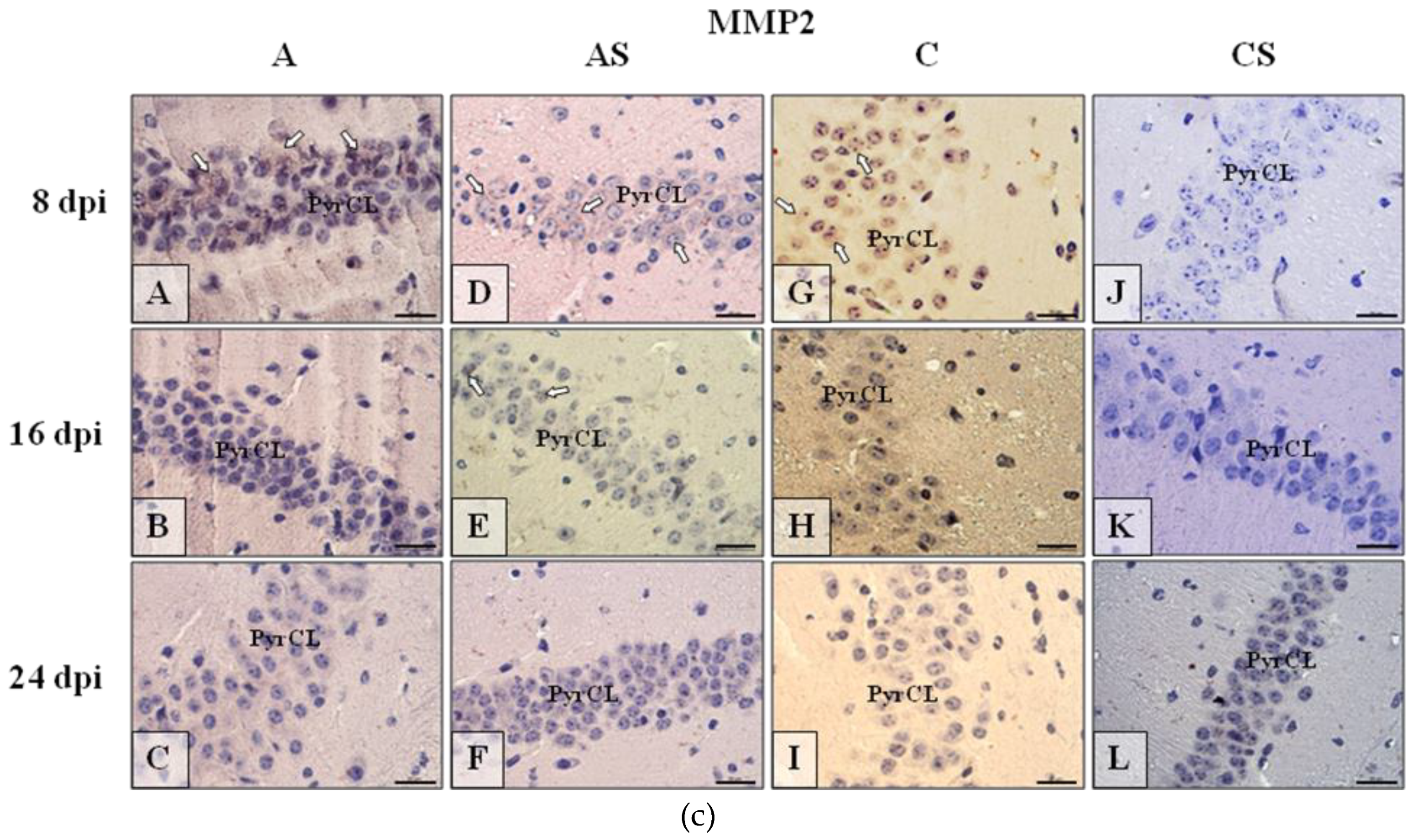
2.6. Immunohistochemistry
2.6.1. MMP-2
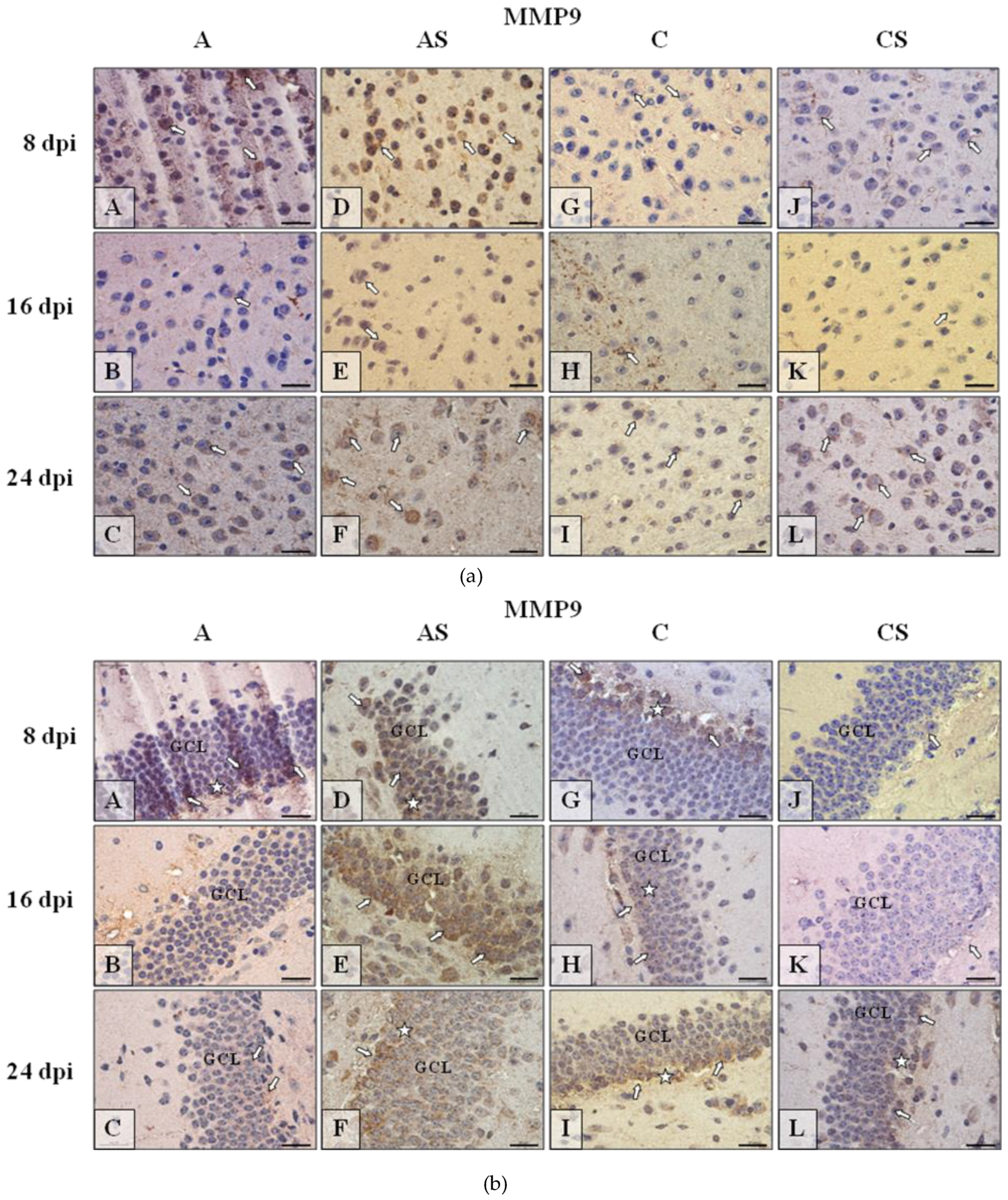
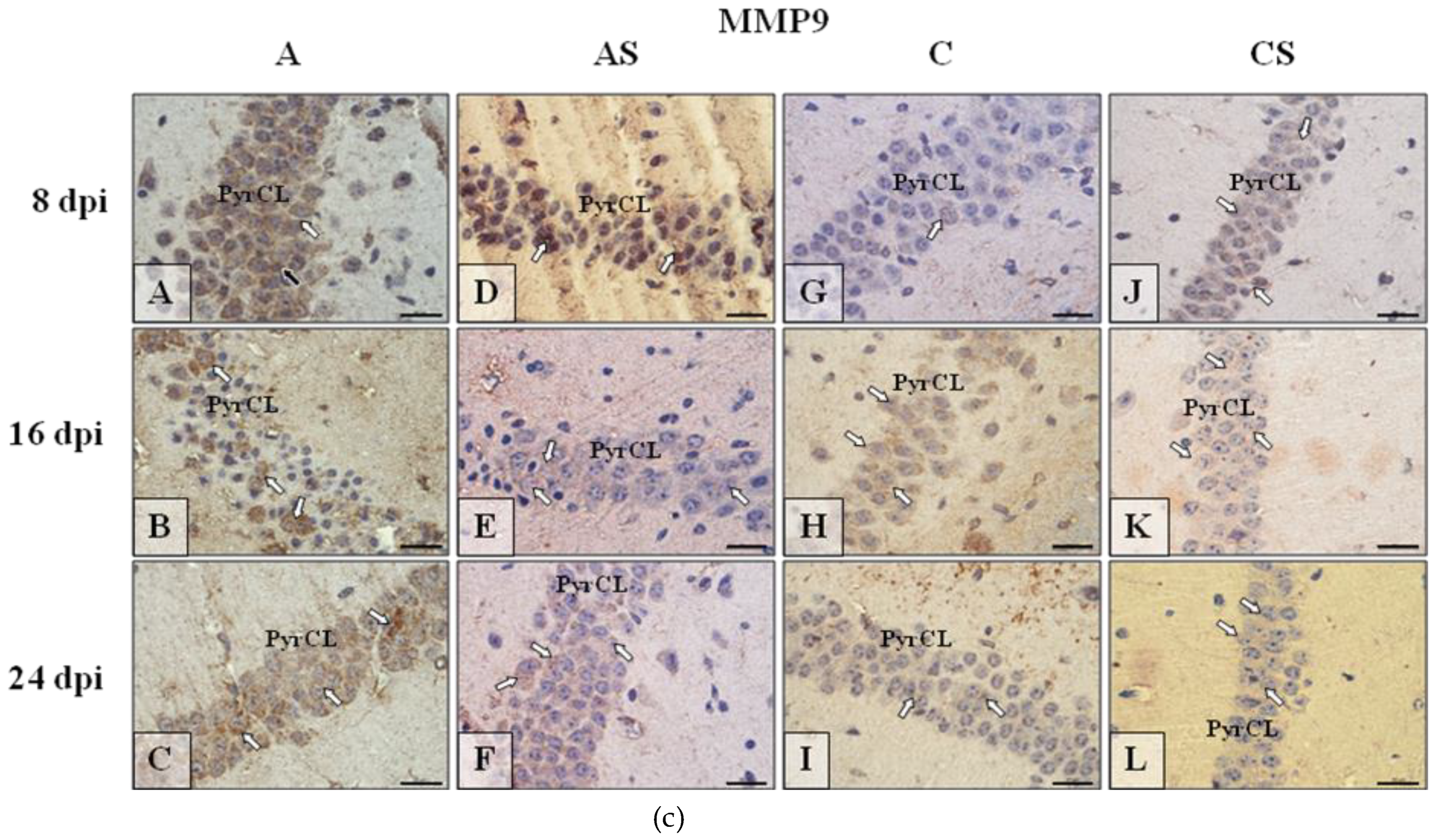
2.6.2. MMP-9
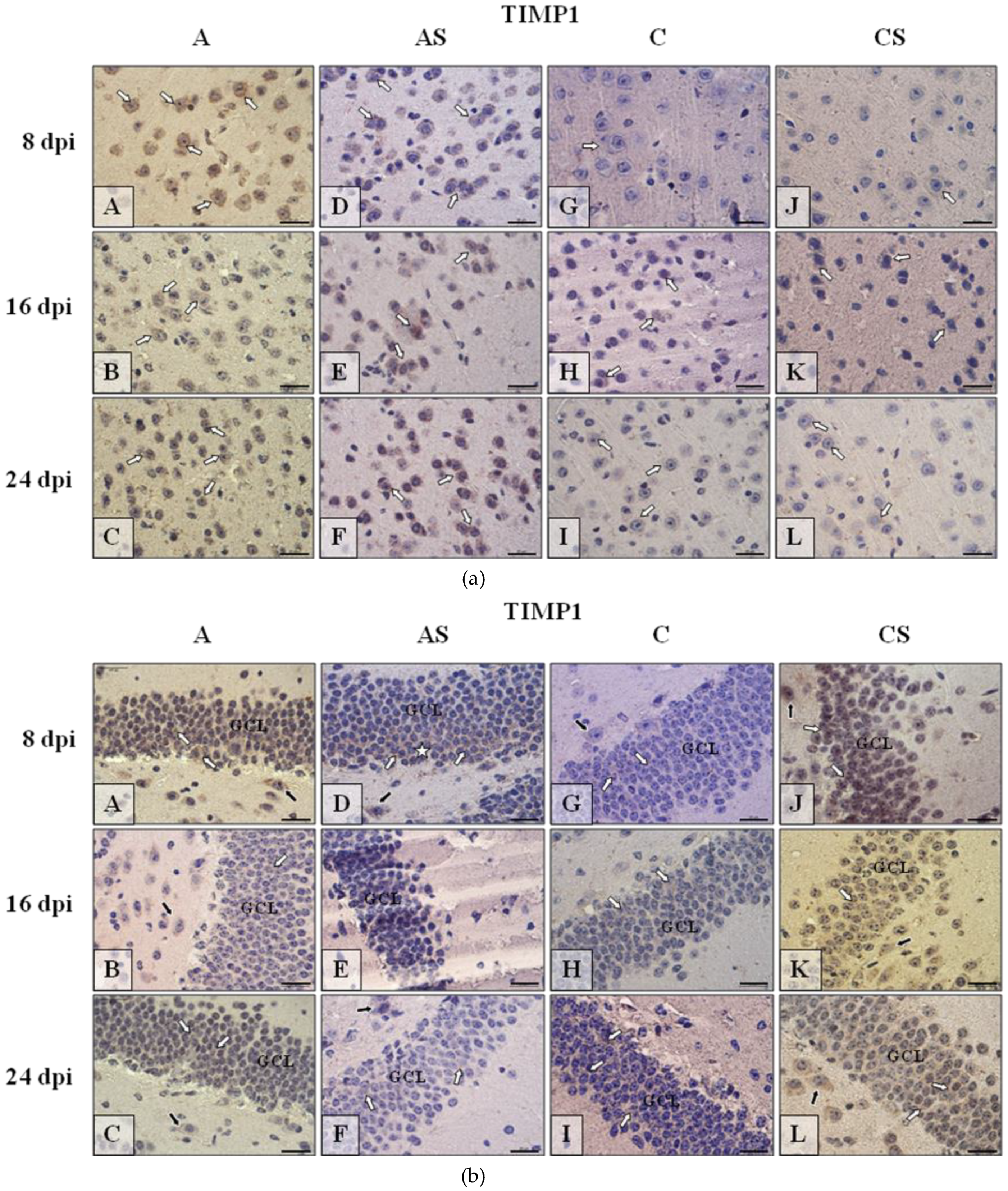
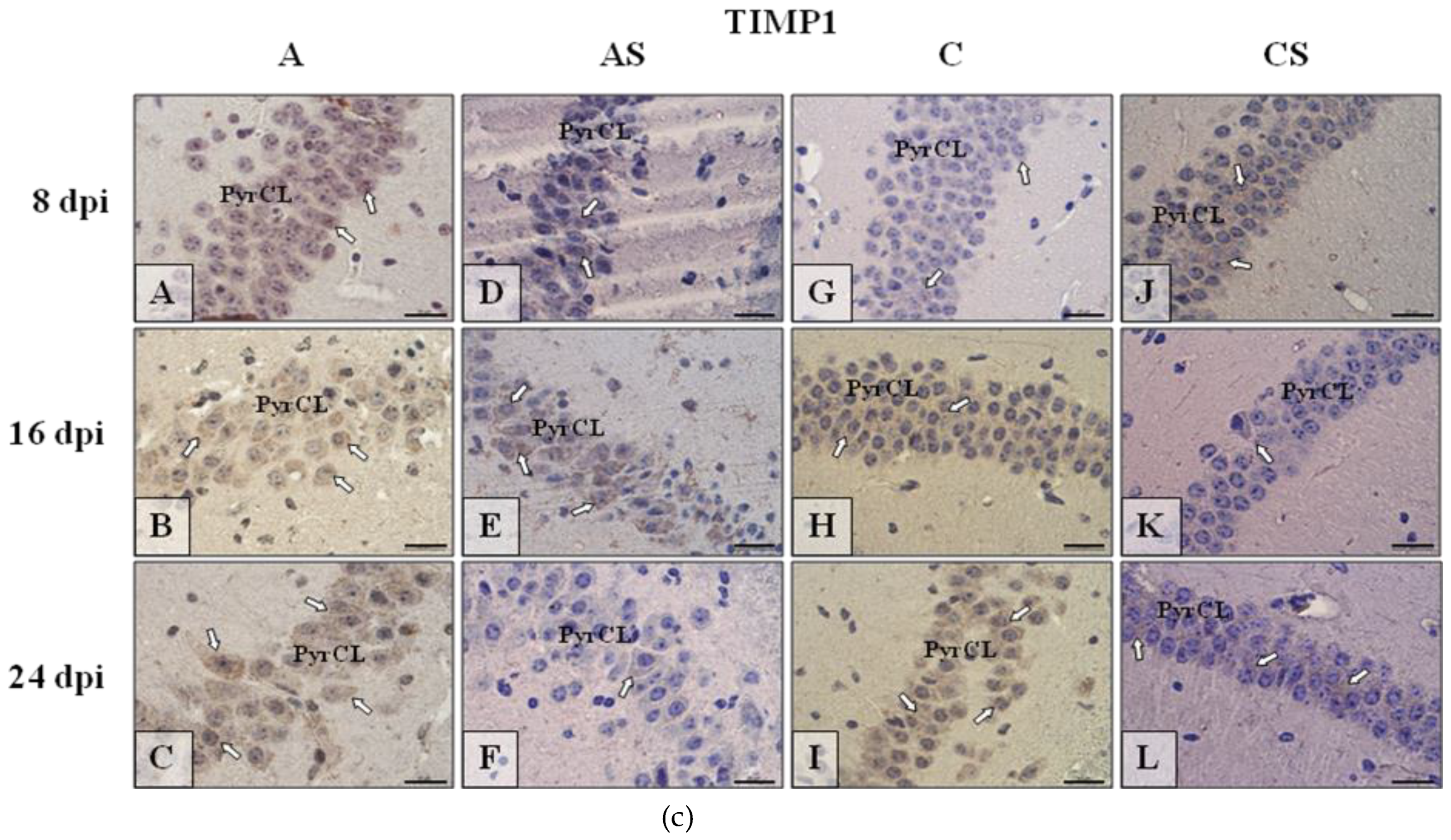
2.6.3. TIMP-1
2.6.4. TIMP-3
3. Discussion
4. Materials and Methods
4.1. Experimental Animal Model
- immunocompetent control group uninfected mice (C, n = 18),
- immunocompetent Acanthamoeba spp. infected mice (A, n = 30),
- immunosuppressed Acanthamoeba spp. infected mice (AS, n = 30),
- immunosuppressed uninfected mice (CS, n = 18).
4.2. Determination of MMP-2, MMP-9, TIMP-3, TIMP-1 in the Brain
4.3. Measurement of Total Protein Concentration by BCA
4.4. I.mmunolocation of MMP-2, MMP-9, TIMP-1 and TIMP-3 by Immunohistochemical Methods
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| A | immunocompetent Acanthamoeba spp. infected mice |
| ADAM | adamalysin-like proteinases |
| AM 22 | amoebic strain no. 22 |
| AML | acute myeloid leukemia |
| AS | immunosuppressed Acanthamoeba spp. infected mice |
| BBB | blood brain barrier |
| BCA | bicinchoninic acid |
| C | immunocompetent uninfected control group mice |
| cAMP | cyclic adenosine monophosphate |
| CNS | central nervous system |
| COX-2 | cyclooxygenase-2 |
| CS | immunosuppressed uninfected mice |
| CSF | cerebrospinal fluid |
| Dpi | day post infection |
| GAE | granulomatous amebic encephalitis |
| GCL | granular cell layer |
| HIV | human immunodeficiency virus |
| IHC | Immunohistochemistry |
| IL-1 | interleukin 1 |
| IL-6 | interleukin 6 |
| IL-23 | interleukin 23 |
| LTP | long term potentiation |
| MMPs | matrix metalloproteinases |
| MMP-2 | matrix metalloproteinase-2 |
| MMP-9 | matrix metalloproteinase-9 |
| MPS | methylprednisolone sodium succinate |
| NK | natural killers |
| PBS | phosphate-buffered saline |
| PDGF | platelet-derived growth factor |
| PGE2 | Prostaglandin E2 |
| PKDL | Post-kala-azar dermal leishmaniasis |
| PyrCL | pyramidal cell layer |
| SD | standard deviation |
| TACE/ADAM17 | ADAM metallopeptidase domain 17 |
| TGF-β | transforming growth factor—β |
| TIMP | tissue inhibitor of metalloproteinase |
| TIMP-1 | tissue inhibitor of metalloproteinase-1 |
| TIMP-3 | tissue inhibitor of metalloproteinase-3 |
| TNF | tumor necrosis factor |
| TNF-α | tumor necrosis factor—α |
References
- Memari, F.; Niyyati, M.; Joneidi, Z. Pathogenic Acanthamoeba T4 genotype isolated from mucosal tissue of a patient with HIV infection: A Case Report. Iran. J. Parasitol. 2017, 12, 143–147. [Google Scholar]
- Matsui, T.; Maeda, T.; Kusakabe, S.; Arita, H.; Yagita, K.; Morii, E.; Kanakura, Y. A case report of granulomatous amoebic encephalitis by Group 1 Acanthamoeba genotype T18 diagnosed by the combination of morphological examination and genetic analysis. Diagn. Pathol. 2018, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Ong, T.Y.Y.; Khan, N.A.; Siddiqui, R. Brain-eating amoebae: Predilection sites in the brain and disease outcome. J. Clin. Microbiol. 2017, 55, 1989–1997. [Google Scholar] [CrossRef]
- Siddiqui, R.; Khan, N.A. Biology and pathogenesis of Acanthamoeba. Parasit Vectors 2012, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A. Acanthamoeba and the blood-brain barrier: The breakthrough. J. Med. Microbiol. 2008, 57, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Morales, J.; Khan, N.A.; Walochnik, J. An update on Acanthamoeba keratitis: Diagnosis, pathogenesis and treatment. Parasite 2015, 22, 10. [Google Scholar] [CrossRef]
- Elsheikha, H.M.; Khan, N.A. Protozoa traversal of the blood-brain barrier to invade the central nervous system. FEMS Microbiol. Rev. 2010, 34, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Marciano-Cabral, F.; Cabral, G. Acanthamoeba spp. as agents of disease in humans. Clin. Microbiol. Rev. 2003, 16, 273–307. [Google Scholar] [CrossRef]
- Masocha, W.; Kristensson, K. Passage of parasites across the blood-brain barrier. Virulence 2012, 3, 202–212. [Google Scholar] [CrossRef] [Green Version]
- Lakhan, S.E.; Kirchgessner, A.; Tepper, D.; Leonard, A. Matrix metalloproteinases and blood-brain barrier disruption in acute ischemic stroke. Front. Neurol. 2013, 4, 32. [Google Scholar] [CrossRef]
- Mukherjee, A.; Swarnakar, S. Implication of matrix metalloproteinases in regulating neuronal disorder. Mol. Biol. Rep. 2015, 42, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rempe, R.G.; Hartz, A.M.; Bauer, B. Matrix metalloproteinases in the brain and blood-brain barrier: Versatile breakers and makers. J. Cereb. Blood Flow Metab. 2016, 36, 1481–1507. [Google Scholar] [CrossRef] [PubMed]
- Ricci, S.; Grandgirard, D.; Wenzel, M.; Braccini, T.; Salvatore, P.; Oggioni, M.R.; Leib, S.L.; Koedel, U. Inhibition of matrix metalloproteinases attenuates brain damage in experimental meningococcal meningitis. BMC Infect. Dis. 2014, 14, 726. [Google Scholar] [CrossRef] [PubMed]
- Bruschi, F.; Pinto, B. The significance of matrix metalloproteinases in parasitic infections involving the central nervous system. Pathogens 2013, 2, 105–129. [Google Scholar] [CrossRef] [PubMed]
- Geurts, N.; Opdenakker, G.; Van den Steen, P.E. Matrix metalloproteinases as therapeutic targets in protozoan parasitic infections. Pharmacol. Ther. 2012, 133, 257–279. [Google Scholar] [CrossRef]
- Lam, C.; Jamerson, M.; Cabral, G.; Carlesso, A.M.; Marciano-Cabral, F. Expression of matrix metalloproteinases in Naegleria fowleri and their role in invasion of the central nervous system. Microbiology 2017, 163, 1436–1444. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Marten, N.W.; Bergmann, C.C.; Macklin, W.B.; Hinton, D.R.; Stohlman, S.A. Expression of matrix metalloproteinases and their tissue inhibitor during viral encephalitis. J. Virol. 2005, 79, 4764–4773. [Google Scholar] [CrossRef]
- Shukla, V.; Shakya, A.K.; Shukla, M.; Kumari, N.; Krishnani, N.; Dhole, T.N.; Misra, U.K. Circulating levels of matrix metalloproteinases and tissue inhibitors of matrix metalloproteinases during Japanese encephalitis virus infection. Virusdisease 2016, 27, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Visvesvara, G.S. Infections with free-living amebae. Handb. Clin. Neurol. 2013, 114, 153–168. [Google Scholar] [CrossRef]
- Doan, N.; Rozansky, G.; Nguyen, H.S.; Gelsomino, M.; Shabani, S.; Mueller, W.; Johnson, V. Granulomatous amebic encephalitis following hematopoietic stem cell transplantation. Surg. Neurol. Int. 2015, 6, S459–S462. [Google Scholar] [CrossRef]
- Yu, C.F.; Chen, F.H.; Lu, M.H.; Hong, J.H.; Chiang, C.S. Dual roles of tumour cells-derived matrix metalloproteinase 2 on brain tumour growth and invasion. Br. J. Cancer 2017, 117, 1828–1836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, P.; Yang, L.; Zhou, H.Y.; Sun, R.P. Matrix metalloproteinase-9 may be a potential therapeutic target in epilepsy. Med. Hypotheses 2011, 76, 184–186. [Google Scholar] [CrossRef]
- Nagy, V.; Bozdagi, O.; Matynia, A.; Balcerzyk, M.; Okulski, P.; Dzwonek, J.; Costa, R.M.; Silva, A.J.; Kaczmarek, L.; Huntley, G.W. Matrix metalloproteinase-9 is required for hippocampal late-phase long-term potentiation and memory. J. Neurosci. 2006, 26, 1923–1934. [Google Scholar] [CrossRef] [PubMed]
- Jacintho, A.P.P.; Melo, G.D.; Machado, G.F.; Bertolo, P.H.L.; Moreira, P.R.R.; Momo, C.; Souza, T.A.; Vasconcelos, R.O. Expression of matrix metalloproteinase-2 and metalloproteinase-9 in the skin of dogs with visceral leishmaniasis. Parasitol. Res. 2018, 117, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Fares, R.C.; Gomes, J.A.; Garzoni, L.R.; Waghabi, M.C.; Saraiva, R.M.; Medeiros, N.I.; Oliveira-Prado, R.; Sangenis, L.H.; Chambela Mda, C.; de Araújo, F.F.; et al. Matrix metalloproteinases 2 and 9 are differentially expressed in patients with indeterminate and cardiac clinical forms of Chagas disease. Infect. Immun. 2013, 81, 3600–3608. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, F.R.; Lalu, M.M.; Mariano, F.S.; Milanezi, C.M.; Cena, J.; Gerlach, R.F.; Santos, J.E.; Torres-Dueñas, D.; Cunha, F.Q.; Schulz, R.; et al. Increased activities of cardiac matrix metalloproteinases matrix metalloproteinase (MMP)-2 and MMP-9 are associated with mortality during the acute phase of experimental Trypanosoma cruzi infection. J. Infect. Dis. 2008, 197, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.T.; Nance, J.P.; Noor, S.; Wilson, E.H. T cell production of matrix metalloproteases and inhibition of parasite clearance by TIMP-1 during chronic toxoplasma infection in the brain. ASN Neurol. 2010, 3, 1–12. [Google Scholar]
- Rosenberg, G.A.; Dencoff, J.E.; Correa, N., Jr.; Reiners, M.; Ford, C.C. Effect of steroids on CSF matrix metalloproteinases in multiple sclerosis: Relation to blood-brain barrier injury. Neurology 1996, 46, 1626–1632. [Google Scholar] [CrossRef] [PubMed]
- Green, J.A.; Tran, C.T.; Farrar, J.J.; Nguyen, M.T.; Nguyen, P.H.; Dinh, S.X.; Ho, N.D.; Ly, C.V.; Tran, H.T.; Friedland, J.S.; et al. Dexamethasone, cerebrospinal fluid matrix metalloproteinase concentrations and clinical outcomes in tuberculous meningitis. PLoS ONE 2009, 4, e7277. [Google Scholar] [CrossRef]
- Costanzo, R.M.; Perrino, L.A. Peak in matrix metaloproteinases-2 levels observed during recovery from olfactory nerve injury. Neuroreport 2008, 19, 327–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, C.S.; Crocker, S.J. An alternate perspective on the roles of TIMPs and MMPs in pathology. Am. J. Pathol. 2012, 180, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Acar, B.A.; Öztekin, Z.N.; Öztekin, M.F.; Acar, T. Serum MMP-2, MMP-9, TIMP-1 and TIMP-2 levels in multiple sclerosis clinical subtypes and their diagnostic value in the progressive disease course. Biomed. Res. 2014, 25, 343–350. [Google Scholar]
- Yu, W.H.; Yu, S.; Meng, Q.; Brew, K.; Woessner, J.F.J. TIMP-3 binds to sulfated glycosaminoglycans of the extracellular matrix. J. Biol. Chem. 2000, 275, 31226–31232. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.H.; Edwards, D.R.; Murphy, G. Metalloproteinase inhibitors: Biological actions and therapeutic opportunities. J. Cell Sci. 2002, 115, 3719–3727. [Google Scholar] [CrossRef] [PubMed]
- Amour, A.; Slocombe, P.M.; Webster, A.; Butler, M.; Knight, C.G.; Smith, B.J.; Stephens, P.E.; Shelley, C.; Hutton, M.; Knäuper, V.; et al. TNF-alpha converting enzyme (TACE) is inhibited by TIMP-3. FEBS Lett. 1998, 435, 39–44. [Google Scholar] [CrossRef]
- Tsai, C.L.; Chen, W.C.; Hsieh, H.L.; Chi, P.L.; Hsiao, L.D.; Yang, C.M. TNF-α induces matrix metalloproteinase-9-dependent soluble intercellular adhesion molecule-1 release via TRAF2-mediated MAPKs and NF-κB activation in osteoblast-like MC3T3-E1 cells. J. Biomed. Sci. 2014, 21, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, N.A.; Katara, G.K.; Ramesh, V.; Salotra, P. Evidence for involvement of TNFR1 and TIMPs in pathogenesis of post-kala-azar dermal leishmaniasis. Clin. Exp. Immunol. 2008, 154, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Shepherd, N.; Lan, J.; Li, W.; Rane, S.; Gupta, S.K.; Zhang, S.; Dong, J.; Yu, Q. MMPs/TIMPs imbalances in the peripheral blood and cerebrospinal fluid are associated with the pathogenesis of HIV-1-associated neurocognitive disorders. Brain Behav. Immun. 2017, 65, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Lichtinghagen, R.; Seifert, T.; Kracke, A.; Marckmann, S.; Wurster, U.; Heidenreich, F. Expression of matrix metalloproteinase-9 and its inhibitors in mononuclear blood cells of patients with multiple sclerosis. J. Neuroimmunol. 1999, 99, 19–26. [Google Scholar] [CrossRef]
- Łanocha-Arendarczyk, N.; Baranowska-Bosiacka, I.; Kot, K.; Gutowska, I.; Kolasa-Wołosiuk, A.; Chlubek, D.; Kosik-Bogacka, D. Expression and activity of COX-1 and COX-2 in Acanthamoeba sp. infected lungs according to the host immunological status. Int. J. Mol. Sci. 2018, 19, 121. [Google Scholar] [CrossRef] [PubMed]
- Łanocha-Arendarczyk, N.; Baranowska-Bosiacka, I.; Kot, K.; Pilarczyk, B.; Tomza-Marciniak, A.; Kabat-Koperska, J.; Kosik-Bogacka, D. Biochemical profile, liver and kidney selenium (Se) status during acanthamoebiasis in a mouse model. Folia Biol. 2018, 1, 33–40. [Google Scholar] [CrossRef]
- Lanocha, N.; Kosik-Bogacka, D.; Maciejewska, A.; Sawczuk, M.; Wilk, A.; Kuźna-Grygiel, W. The occurrence Acanthamoeba (free living amoeba) in environmental and respiratory samples in Poland. Acta Protozool. 2009, 48, 271–279. [Google Scholar]
- Culbertson, C.G.; Ensminger, P.W.; Overton, W.M. The isolation of additional strains ff pathogenic Hartmanella sp. (Acanthamoeba): Proposed culture method for application to biological material. Am. J. Clin. Pathol. 1965, 43, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Markowitz, S.M.; Sobieski, T.; Martinez, A.J.; Duma, R.J. Experimental Acanthamoeba infections in mice pretreated with methylprednisolone or tetracycline. Am. J. Pathol. 1978, 92, 733–744. [Google Scholar]












© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Łanocha-Arendarczyk, N.; Baranowska-Bosiacka, I.; Gutowska, I.; Kolasa-Wołosiuk, A.; Kot, K.; Łanocha, A.; Metryka, E.; Wiszniewska, B.; Chlubek, D.; Kosik-Bogacka, D. The Activity of Matrix Metalloproteinases (MMP-2, MMP-9) and Their Tissue Inhibitors (TIMP-1, TIMP-3) in the Cerebral Cortex and Hippocampus in Experimental Acanthamoebiasis. Int. J. Mol. Sci. 2018, 19, 4128. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124128
Łanocha-Arendarczyk N, Baranowska-Bosiacka I, Gutowska I, Kolasa-Wołosiuk A, Kot K, Łanocha A, Metryka E, Wiszniewska B, Chlubek D, Kosik-Bogacka D. The Activity of Matrix Metalloproteinases (MMP-2, MMP-9) and Their Tissue Inhibitors (TIMP-1, TIMP-3) in the Cerebral Cortex and Hippocampus in Experimental Acanthamoebiasis. International Journal of Molecular Sciences. 2018; 19(12):4128. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124128
Chicago/Turabian StyleŁanocha-Arendarczyk, Natalia, Irena Baranowska-Bosiacka, Izabela Gutowska, Agnieszka Kolasa-Wołosiuk, Karolina Kot, Aleksandra Łanocha, Emilia Metryka, Barbara Wiszniewska, Dariusz Chlubek, and Danuta Kosik-Bogacka. 2018. "The Activity of Matrix Metalloproteinases (MMP-2, MMP-9) and Their Tissue Inhibitors (TIMP-1, TIMP-3) in the Cerebral Cortex and Hippocampus in Experimental Acanthamoebiasis" International Journal of Molecular Sciences 19, no. 12: 4128. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124128