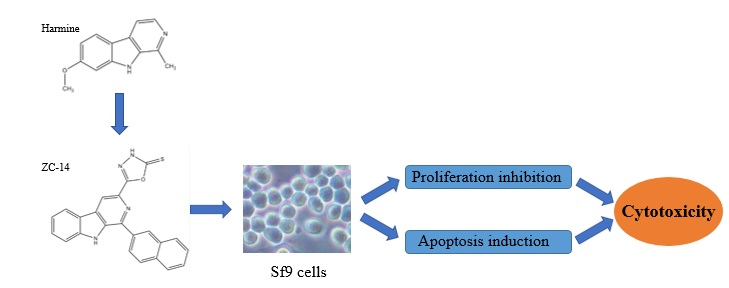
Cytotoxic and Apoptotic Activity of the Novel Harmine Derivative ZC-14 in Sf9 Cells

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
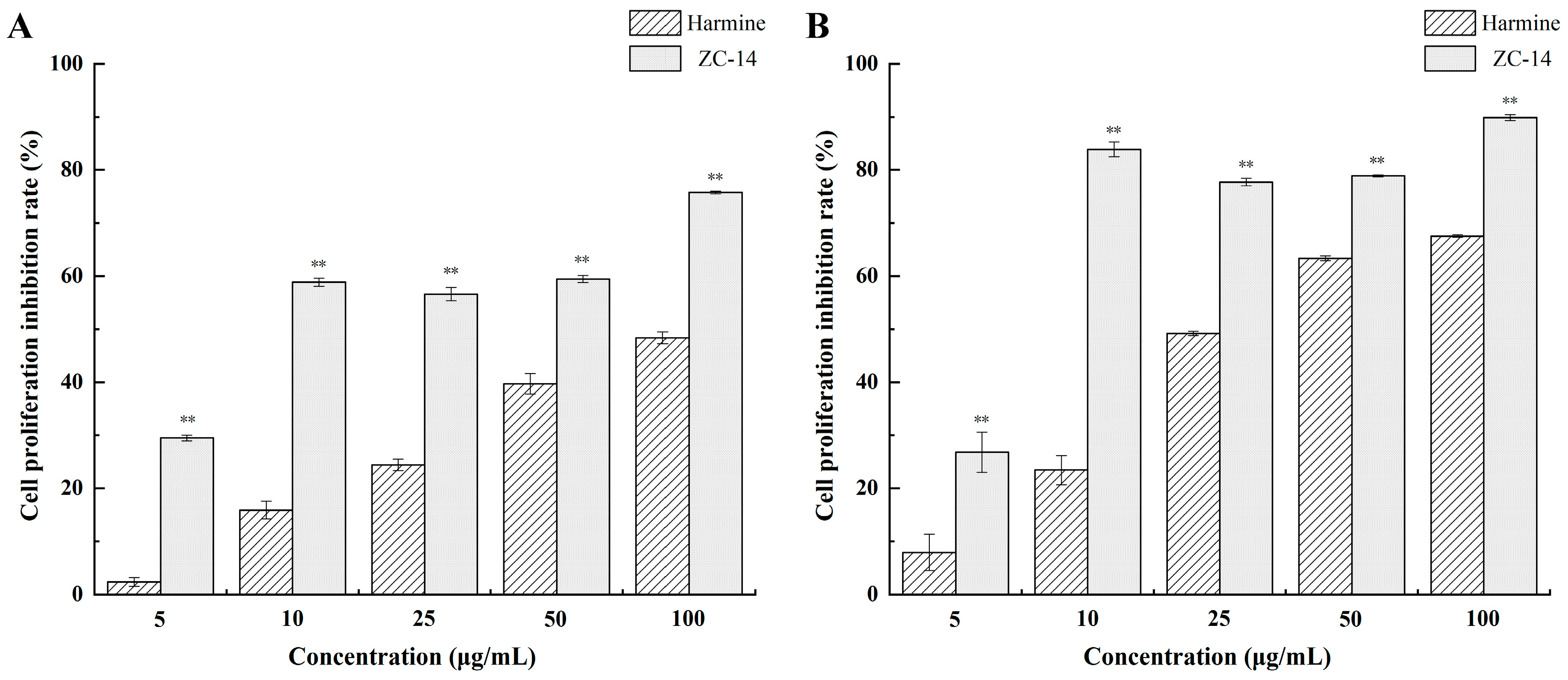
2.1. Harmine and ZC-14 Inhibited the Proliferation of Sf9 Cells
2.2. ZC-14 Induced Morphological Changes in Sf9 Cells
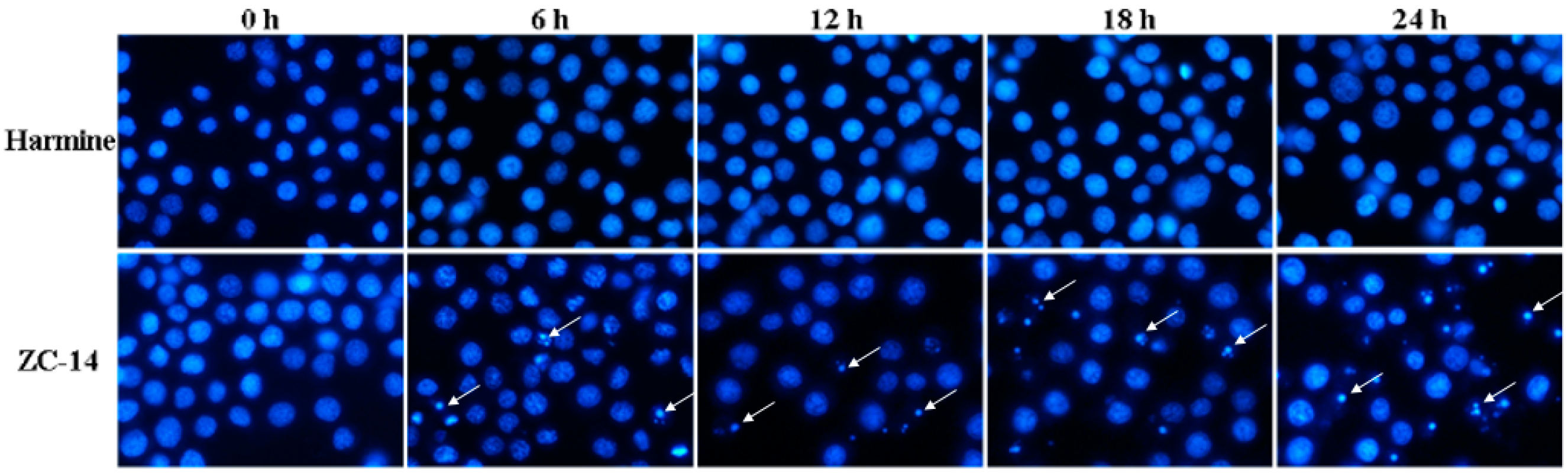
2.3. Nuclear Condensation/Fragmentation Appeared in Sf9 Cells after ZC-14 Treatments
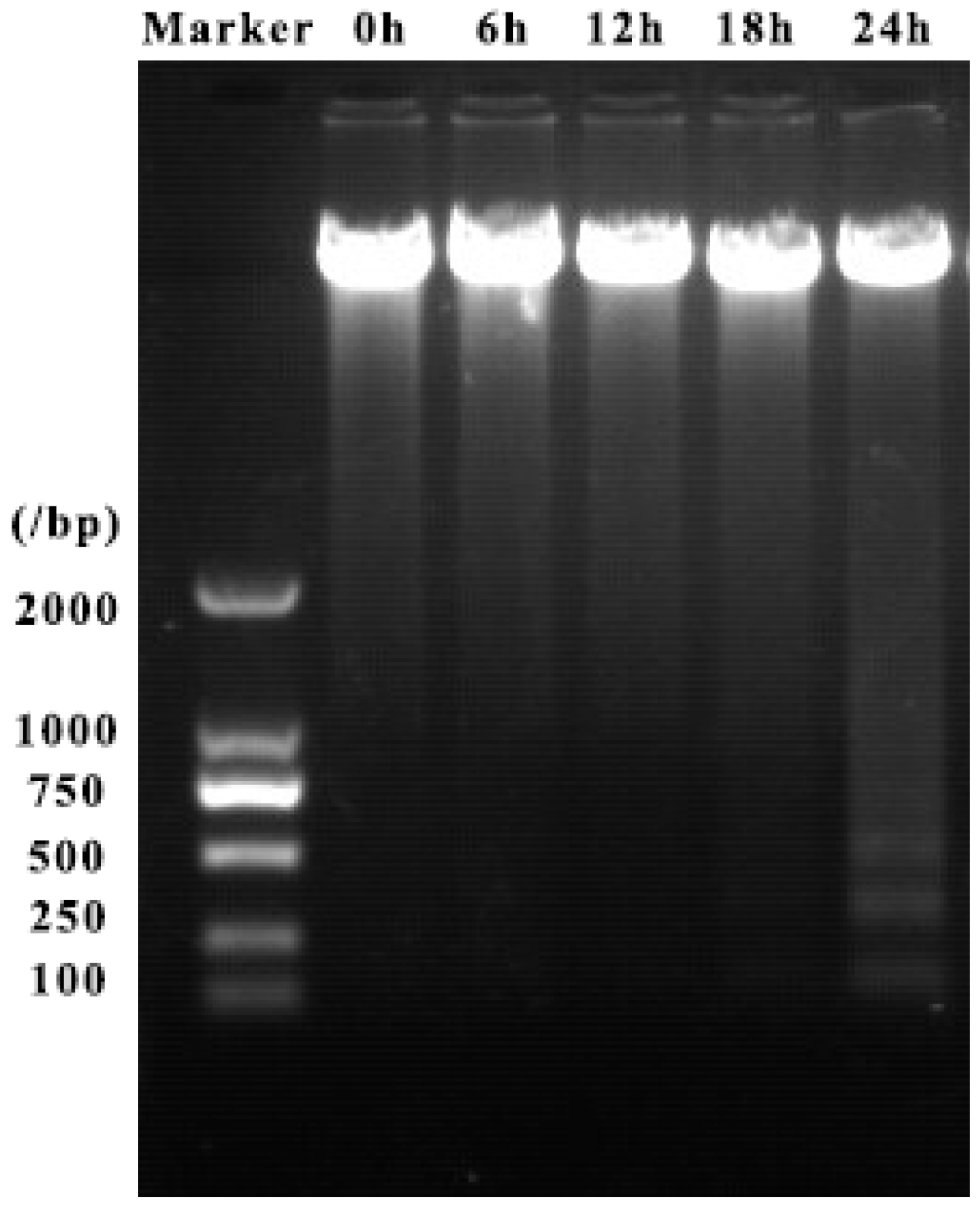
2.4. ZC-14 Treatments Induced DNA Fragmentation in Sf9 Cells
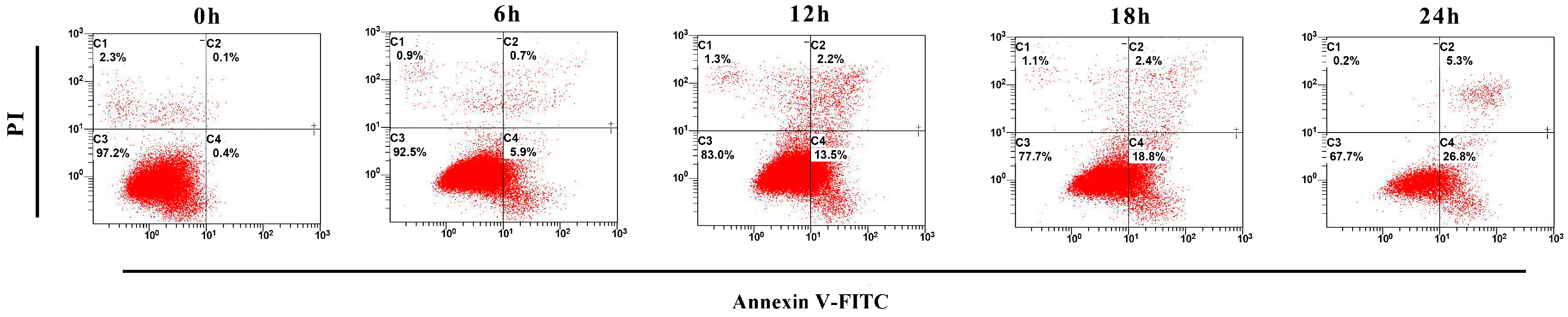
2.5. Apoptotic Rate Induced by ZC-14 Treatments in Sf9 Cells
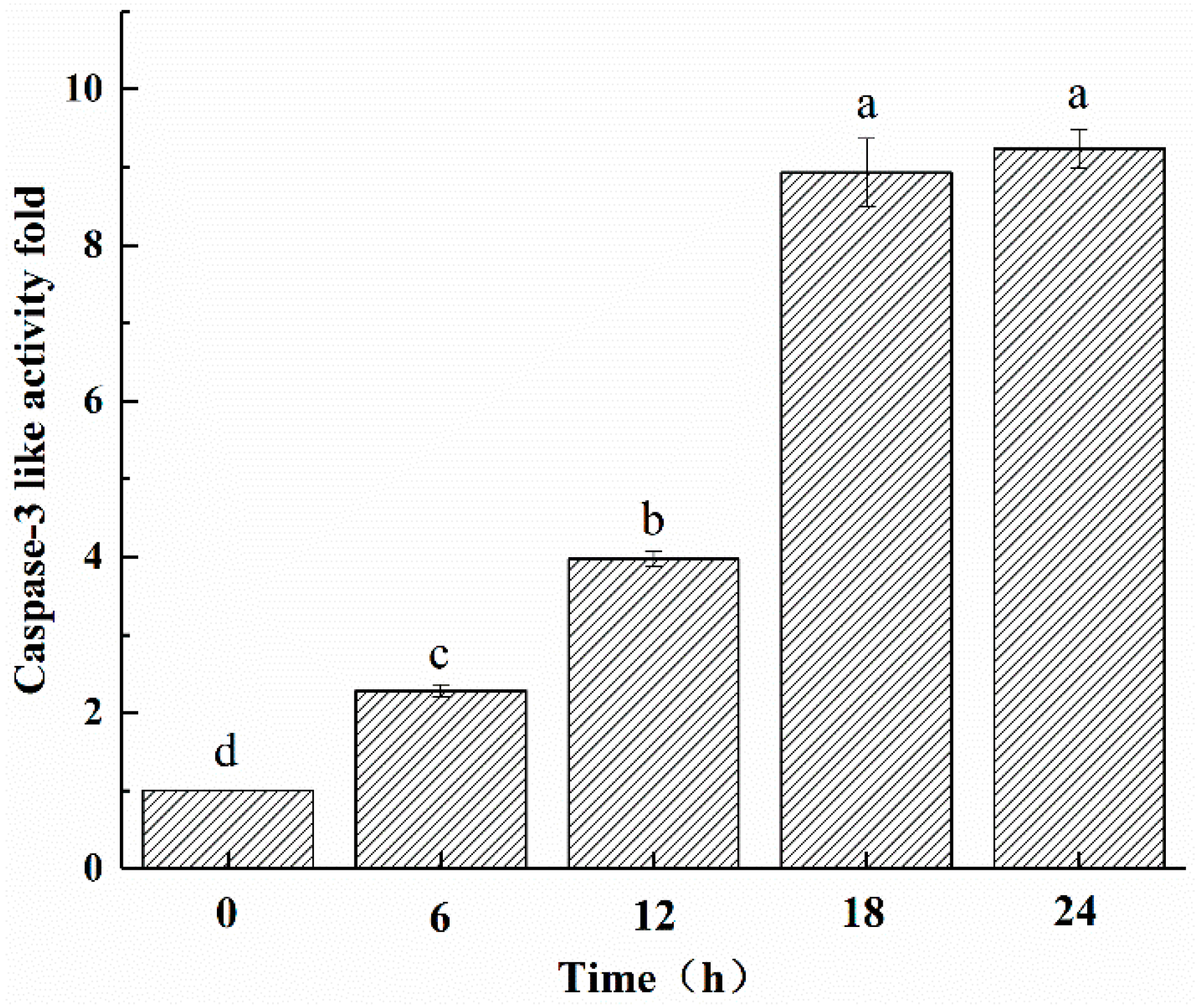
2.6. ZC-14 Treatments Increased the Caspase-3 Like Activity in Sf9 Cells
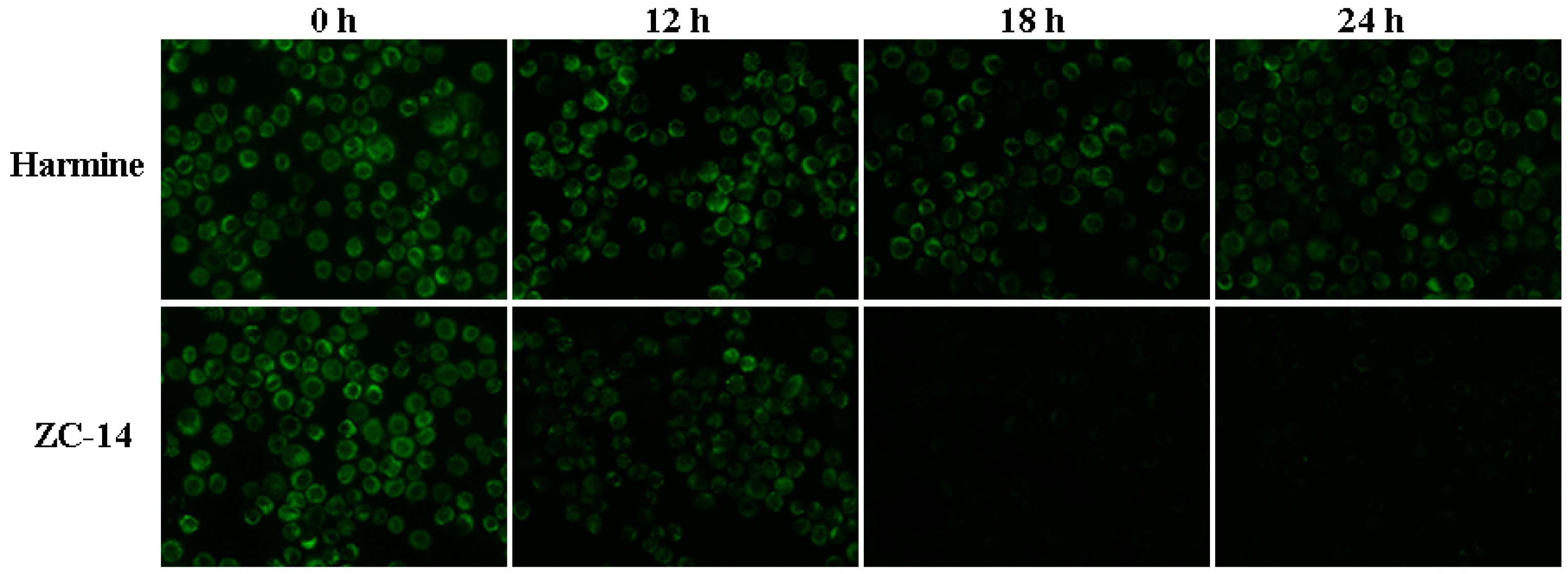
2.7. Loss of Mitochondrial Membrane Potential (MMP) Induced by ZC-14 Treatments in Sf9 Cells
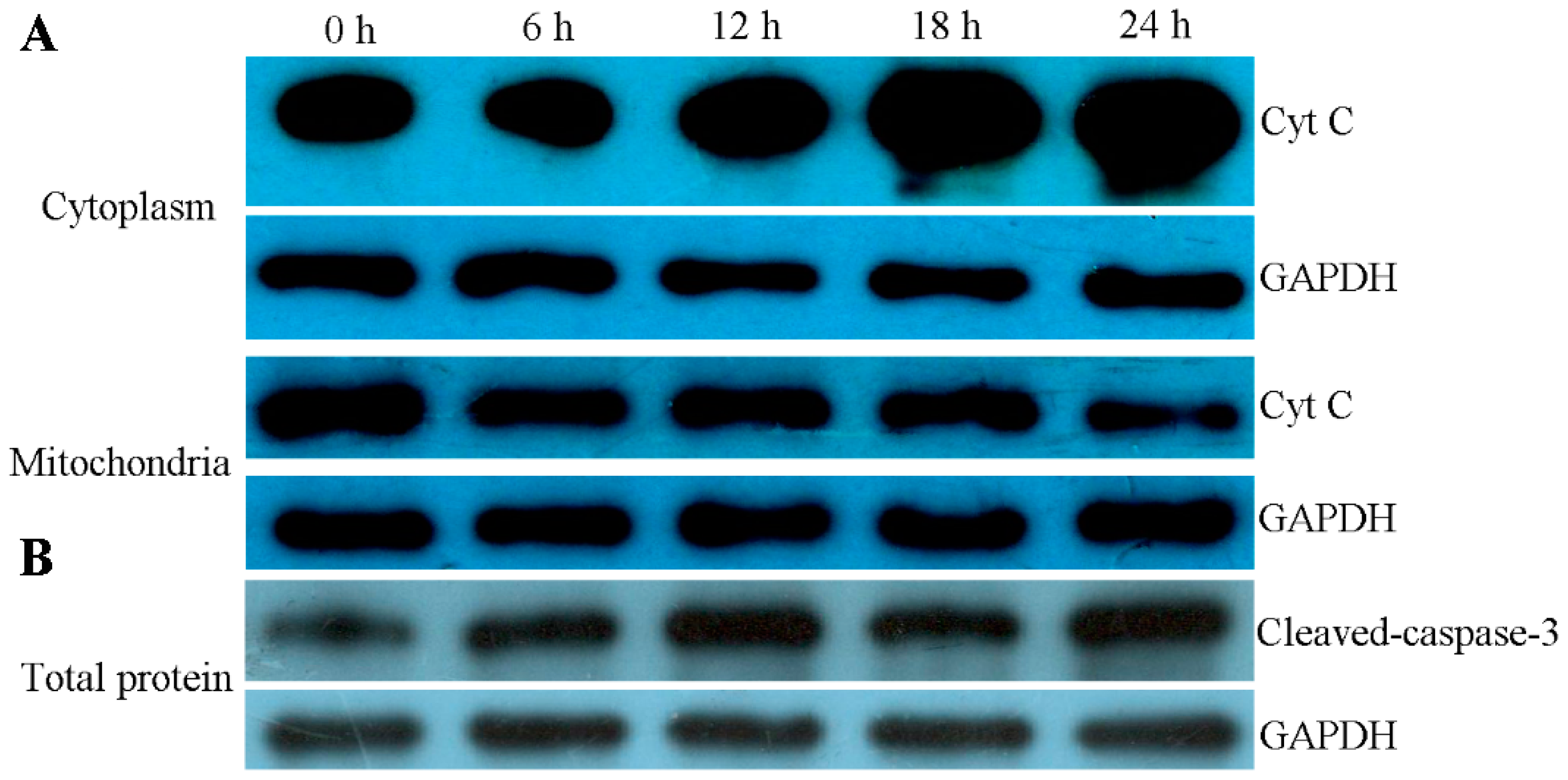
2.8. ZC-14 Treatments Activated Apoptotic Proteins in Sf9 Cells
3. Discussion
4. Materials and Methods
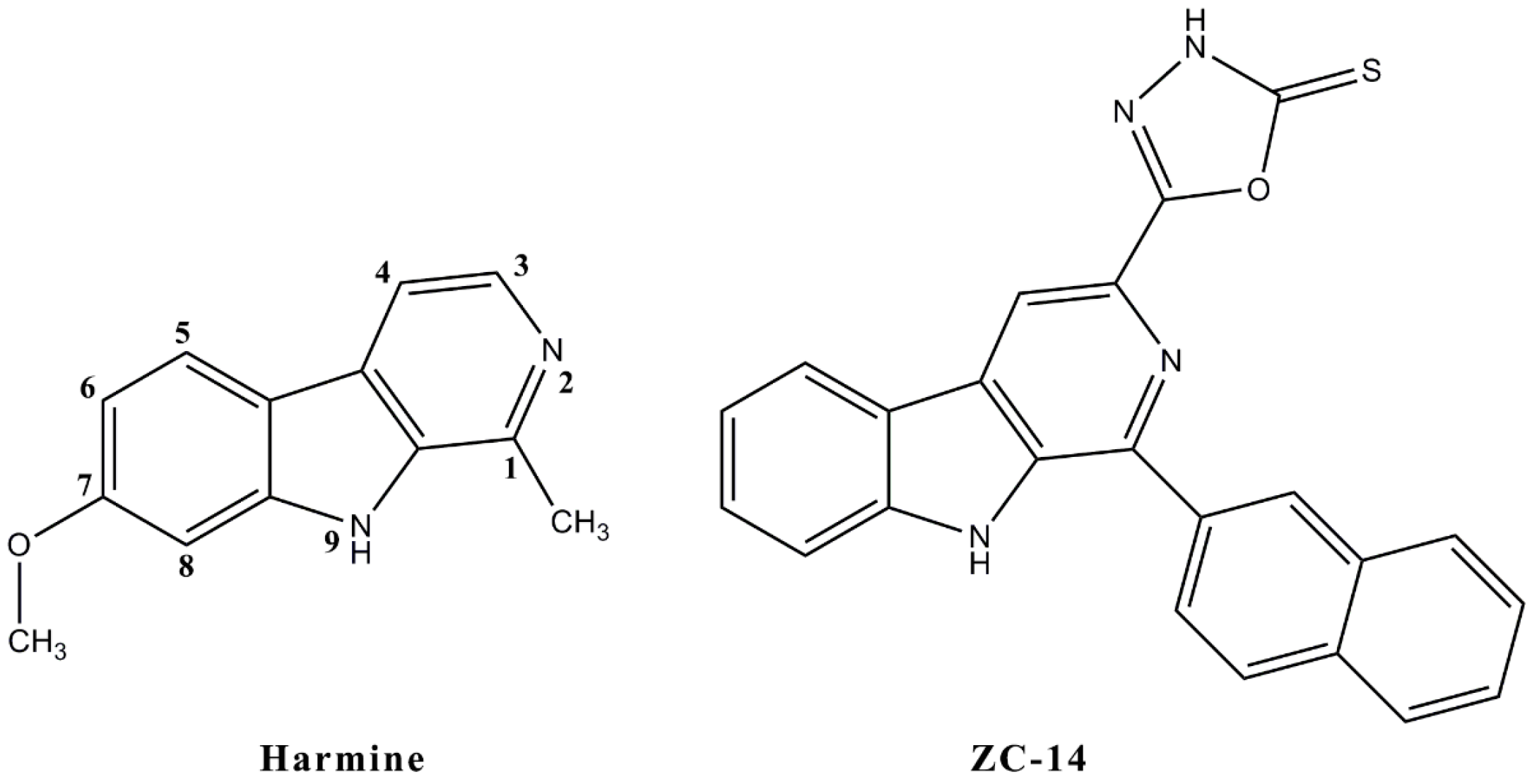
4.1. Chemicals
4.2. Cell Culture and Reagents
4.3. Cell Proliferation Assay
4.4. Cell Morphological Observation
4.5. Hoechst 33258 Staining Analysis
4.6. DNA Fragmentation Assay
4.7. Flow Cytometry Assay
4.8. Caspase-3 Like Activity Assay
4.9. Rhodamine 123 Staining
4.10. Western Blot
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflict of interest
References
- Li, C.; Wang, Y.; Wang, C.; Yi, X.; Li, M.; He, X. Anticancer activities of harmine by inducing a pro-death autophagy and apoptosis in human gastric cancer cells. Phytomedicine 2017, 28, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Frost, D.; Meechoovet, B.; Wang, T.; Gately, S.; Giorgetti, M.; Shcherbakova, I.; Dunckley, T. β-Carboline compounds, including harmine, inhibit DYRK1A and tau phosphorylation at multiple Alzheimer’s disease-related sites. PLoS ONE 2011, 6, e19264. [Google Scholar] [CrossRef] [PubMed]
- Moloudizargari, M.; Mikaili, P.; Aghajanshakeri, S.; Asghari, M.H.; Shayegh, J. Pharmacological and therapeutic effects of Peganum harmala and its main alkaloids. Pharmacogn. Rev. 2013, 7, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Bouayad, N.; Rharrabe, K.; Ghailani, N.N.; Jbilou, R.; Castanera, P.; Ortego, F. Insecticidal effects of Moroccan plant extracts on development, energy reserves and enzymatic activities of Plodia interpunctella. Span. J. Agric. Res. 2013, 11, 189–198. [Google Scholar] [CrossRef]
- Ortego, F.; Rodríguez, B.; Castañera, P. Effects of neo-clerodane diterpenes from Teucrium on feeding behavior of Colorado potato beetle larvae. J. Chem. Ecol. 1995, 21, 1375–1386. [Google Scholar] [CrossRef] [PubMed]
- Bouayad, N.; Rharrabe, K.; Lamhamdi, M.; Nourouti, N.G.; Sayah, F. Dietary effects of harmine, a beta-carboline alkaloid, on development, energy reserves and α-amylase activity of Plodia interpunctella Hubner (Lepidoptera: Pyralidae). Saudi J. Biol. Sci. 2012, 19, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhao, T.; Cheng, X.M.; Wang, C.H.; Wang, Z.T. Characterization and determination of trace alkaloids in seeds extracts from Peganum harmala Linn. Using LC-ESI-MS and HPLC. Acta Chromatogr. 2013, 25, 221–240. [Google Scholar] [CrossRef]
- Liu, X.; Li, M.; Tan, S.; Wang, C.; Fan, S.; Huang, C. Harmine is an inflammatory inhibitor through the suppression of NF-kB signaling. Biochem. Biophysi. Res. Commu. 2017, 489, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Nafisi, S.; Bonsaii, M.; Maali, P.; Khalilzadeh, M.A.; Manoucheri, F. β-Carboline alkaloids bind DNA. J. Photochem. Photobiol. B 2010, 100, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Sobhani, A.M.; Ebrahimi, S.A.; Mahmoudian, M. An in vitro evaluation of human DNA topoisomerase I inhibition by Peganum harmala L. seeds extract and its acarboline alkaloids. J. Pharm. Pharm. Sci. 2002, 5, 19–23. [Google Scholar] [PubMed]
- Yang, Y.D.; Cheng, X.M.; Liu, W.; Chou, G.; Wang, Z.; Wang, C. Potent AChE and BChE inhibitors isolated from seeds of Peganum harmala Linn by a bioassay-guided fractionation. J. Ethnopharmacol. 2015, 168, 279–286. [Google Scholar] [CrossRef] [PubMed]
- He, D.D.; Wu, H.; Wei, Y.; Liu, W.; Huang, F.; Shi, H.; Zhang, B.; Wu, X.; Wang, C. Effects of harmine, an acetylcholinesterase inhibitor, on spatial learning and memory of APP/PS1 transgenic mice and scopolamine-induced memory impairment mice. Eur. J. Pharmacol. 2015, 768, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Lia, S.; Cheng, X.; Wang, C. A review on traditional uses, phytochemistry, pharmacology, pharmacokinetics and toxicology of the genus Peganum. J. Ethnopharmacol. 2017, 203, 127–162. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y.; Suzuki, T.; Ishii, H.; Ogata, A.; Nakae, D. Mitochondrial dysfunction and biotransformation of β-carboline alkaloids, harmine and harmaline, on isolated rat hepatocytes. Chem. Biol. Interact. 2010, 188, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Rommelspacher, H.; Nanz, C.; Borbe, H.O.; Fehske, K.J.; Müller, W.E.; Wollert, U. 1-Methyl-β-carboline (harmane), a potent endogenous inhibitor of benzodiazepine receptor binding. Naunyn Schmiedebergs Arch. Pharmacol. 1980, 314, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Li, S.P.; Liu, W.; Teng, L.; Cheng, X.; Wang, Z.; Wang, C. Metabolites identification of harmane in vitro/in vivo in rats by ultra-performance liquid chromatography combined with electrospray ionization quadrupole time-of-flight tandem mass spectrometry. J. Pharm. Biomed. Anal. 2014, 92, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Miralles, A.; Esteban, S.; Sastre-Coll, A.; Moranta, D.; Asensio, V.J.; García-Sevilla, J.A. High-affinity binding of, β-carbolines to imidazoline I-2B receptors and MAO-A in rat tissues: Norharman blocks the effect of morphine withdrawal on DOPA/noradrenaline synthesis in the brain. Eur. J. Pharmacol. 2005, 518, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Zhang, Y.; Weng, Q.; Hu, M.; Zhong, G. Cytotoxic and insecticidal activities of derivatives of harmine, a natural insecticidal component isolated from Peganum harmala. Molecules 2010, 15, 7775–7791. [Google Scholar] [CrossRef] [PubMed]
- Nenaah, G. Toxicity and growth inhibitory activities of methanol extract and the β-carboline alkaloids of Peganum harmala L. against two coleopteran stored-grain pests. J. Stored Prod. Res. 2011, 47, 255–261. [Google Scholar] [CrossRef]
- Zhong, G.; Cui, G.; Yi, X.; Sun, R.; Zhang, J. Insecticide cytotoxicology in China: Current status and challenges. Pestic. Biochem. Phys. 2016, 132, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Ruan, S.; Jia, F.; Li, J. Potential antitumor effect of harmine in the treatment of Thyroid Cancer. Evid. Based Complement. Altern. 2017, 2017, 9402615. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, Q.; Liu, Z.; Lin, L.; Zhang, X.; Cao, M.; Jiang, J. Harmine induces cell cycle arrest and mitochondrial pathway-mediated cellular apoptosis in SW620 cells via inhibition of the Akt and ERK signaling pathways. Oncol. Rep. 2016, 35, 3363–3370. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.O.V.; Mishra, S.; Tantak, M.P.; Nikhil, K.; Sadana, R.; Shah, K.; Kumar, D. Design, synthesis and in vitro cytotoxicity studies of novel β-carbolinium bromides. Bioorg. Med. Chem. Lett. 2017, 27, 1379–1384. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Jia, J.; Luo, J.; Weng, Q.; Zhong, G. Modes of action of six different botanical alkaloids against Spodoptera litura cultured cell line SL-1. J. Huazhong Agric. Univ. 2010, 29, 31–36. [Google Scholar]
- Cao, M.R.; Li, Q.; Liu, Z.L.; Liu, H.H.; Wang, W.; Liao, X.L.; Pan, Y.L.; Jiang, J.W. Harmine induces apoptosis in HepG2 cells via mitochondrial signaling pathway. Hepatobiliary Pancreat. Dis. Int. 2011, 10, 599–604. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, C.; Jiang, C.; Zeng, H.; He, X. Novel mechanism of harmaline on inducing G2/M cell cycle arrest and apoptosis by up-regulating Fas/FasL in SGC-7901 cells. Sci. Rep. 2015, 5, 18613. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Sun, R.; Jia, Y.; Chen, Q.; Tu, R.F.; Li, K.; Zhang, X.D.; Du, R.L.; Cao, R. Synthesis and mechanisms of action of novel harmine derivatives as potential antitumor agents. Sci. Rep. 2016, 6, 33204. [Google Scholar] [CrossRef] [PubMed]
- Lamchouri, F.; Zemzami, M.; Jossang, A.; Abdellatif, A.; Israili, Z.H.; Lyoussi, B. Cytotoxicity of alkaloids isolated from Peganum harmala seeds. Pak. J. Pharm. Sci. 2013, 26, 699–706. [Google Scholar] [PubMed]
- Zhang, L.; Zhang, F.; Zhang, W.; Chen, L.; Gao, N.; Men, Y.; Xu, X.; Jiang, Y. Harmine suppresses homologous recombination repair and inhibits proliferation of hepatoma cells. Cancer Biol. Ther. 2015, 16, 1585–1592. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Kesuma, D.; Wang, J.; Deng, Y.; Duan, J.; Wang, J.H.; Qi, R.Z. Specific inhibition of cyclin-dependent kinases and cell proliferation by harmine. Biochem. Biophys. Res. Commun. 2004, 317, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Hamsa, T.P.; Kuttan, G. Harmine inhibits tumour specific neo-vessel formation by regulating VEGF, MMP, TIMP and pro-inflammatory mediator both in vivo and in vitro. Eur. J. Pharmacol. 2010, 649, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Al-Saikhan, F.I.; Ansari, M.N. Evaluation of the diuretic and urinary electrolyte effects of methanolic extract of Peganum harmala L. in Wistar albino rats. Saudi J. Biol. Sci. 2016, 23, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Brierley, D.I.; Davidson, C. Developments in harmine pharmacology-Implications for ayahuasca use and drug-dependence treatment. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 39, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Ayoob, I.; Hazari, Y.M.; Lone, S.H.; Rehman, S.; Khuroo, M.A.; Fazili, K.M.; Bhat, K.A. Phytochemical and cytotoxic evaluation of Peganum Harmala: Structure activity relationship studies of harmine. ChemistrySelect 2017, 2, 2965–2968. [Google Scholar] [CrossRef]
- Xu, Q.B.; Chen, X.F.; Feng, J.; Miao, J.F.; Liu, J.; Liu, F.T.; Niu, B.X.; Cai, J.Y.; Huang, C.; Zhang, Y.; et al. Design, synthesis and biological evaluation of hybrids of β-carboline and salicylic acid as potential anticancer and apoptosis inducing agents. Sci. Rep. 2016, 6, 36238. [Google Scholar] [CrossRef] [PubMed]
- Shankaraiah, N.; Jadala, C.; Nekkanti, S.; Senwar, K.R.; Agesh, N.; Shrivastava, S.; Naidu, V.G.; Sathish, M.; Kamal, A. Design and synthesis of C3-tethered 1,2,3-triazolo-β-carboline derivatives: Anticancer activity, DNA-binding ability, viscosity and molecular modeling studies. Bioorg. Chem. 2016, 64, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Kumar, B.A.; Singh, J.; Maiti, B.C.; Maity, T.K. Synthesis of some novel 3,5-disubstituted 1,3,4-oxadiazole derivatives and anticancer activity on EAC animal model. Med. Chem. Res. 2011, 20, 1206–1213. [Google Scholar] [CrossRef]
- Xu, W.M.; Han, F.F.; He, M.; Hu, D.Y.; He, J.; Yang, S.; Song, B.A. Inhibition of Tobacco Bacterial Wilt with Sulfone Derivatives Containing an 1,3,4-Oxadiazole Moiety. J. Agric. Food Chem. 2012, 60, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Formagio, A.S.N.; Tonin, L.T.D.; Foglio, M.A.; Madjarof, C.; de Carvalho, J.E.; da Costa, W.F.; Cardoso, F.P.; Sarragiotto, M.H. Synthesis and antitumoral activity of novel 3-(2-substituted-1,3,4-oxadiazol-5-yl) and 3-(5-substituted-1,2,4-triazol-3-yl) β-carboline derivatives. Bioorg. Med. Chem. 2008, 16, 9660–9667. [Google Scholar] [CrossRef] [PubMed]
- Boyapati, S.; Kulandaivelu, U.; Sangu, S.; Vanga, M.R. Synthesis, Antimicrobial Evaluation, and Docking Studies of Novel 4-Substituted Quinazoline Derivatives as DNA-Gyrase Inhibitors. Arch. Pharm. Chem. Life Sci. 2010, 10, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.J.; Zhang, J.J.; Jiang, Z.Y.; Zhong, G.H. Design, Synthesis and Bioactivity Evaluation of Novel β-carboline 1,3,4-oxadiazole Derivatives. Molecules 2017, 22, 1811. [Google Scholar] [CrossRef] [PubMed]
- Suganuma1, I.; Ushiyama, T.; Yamada, H.; Iwamoto, A.; Kobayashi, M.; Ikeda, M. Cloning and characterization of a dronc homologue in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2011, 41, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Civciristov, S.; Hawkins, C.J.; Clem, R.J. SfDronc, an initiator caspase involved in apoptosis in the fall armyworm Spodoptera frugiperda. Insect Biochem. Mol. Biol. 2013, 43, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Shu, B.; Wang, W.; Hu, Q.; Huang, J.; Hu, M.; Zhong, G. A comprehensive study on apoptosis induction by azadirachtin in Spodoptera frugiperda cultured cell line Sf9. Arch. Insect Biochem. Physiol. 2015, 89, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.X.; Zhong, G.H.; Hu, M.Y.; Huang, J.F.; Ge, C.C. Induced apoptosis of camptothecin in Spodoptera frugiperda Sf9 cells. Acta Entomol. Sin. 2011, 54, 894–901. [Google Scholar]
- Yang, M.; Wang, B.; Gao, J.; Zhang, Y.; Xu, W.; Tao, L. Spinosad induces programmed cell death involves mitochondrial dysfunction and cytochrome C release in Spodoptera frugiperda Sf9 cells. Chemosphere 2017, 169, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.F.; Tian, M.; Lv, C.J.; Li, H.Y.; Muhammad, R.; Zhong, G.H. Preliminary studies on induction of apoptosis by abamectin in Spodoptera frugiperda (Sf9) cell line. Pestic. Biochem. Physiol. 2011, 100, 256–263. [Google Scholar] [CrossRef]
- Huang, J.; Lv, C.; Hu, M.; Zhong, G. The mitochondria-mediate apoptosis of Lepidopteran cells induced by Azadirachtin. PLoS ONE 2013, 8, e58499. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Cheng, X.; Meng, Q.; Wang, P.; Shu, B.; Hu, Q.; Hu, M.; Zhong, G. Azadirachtin-induced apoptosis involves lysosomal membrane permeabilization and cathepsin L release in Spodoptera frugiperda Sf9 cells. Int. J. Biochem. Cell Biol. 2015, 64, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Adrain, C.; Martin, S.J. The mitochondrial apoptosome: A killer unleashed by the cytochrome seas. Trends Biochem. Sci. 2001, 26, 390–397. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, Y.; Hui, T.; Wu, H.; Wu, Y.; Wang, W. Reaper homologue IBM1 in silkworm Bombyx mori induces apoptosis upon baculovirus infection. FEBS Lett. 2013, 587, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Sivalingam, K.S.; Paramasivan, P.; Weng, C.F.; Viswanadha, V.P. Neferine potentiates the antitumor effect of cisplatin in human lung adenocarcinoma cells via a mitochondria-mediated apoptosis pathway. J. Cell Biochem. 2017, 118, 2865–2876. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, C.; De Oliveira, F.; Ronot, X.; Mousseau, M.; Leverve, X.; Fontaine, E. Rotenone inhibits the mitochondrial permeability transit on induced cell death in U937 and KB cells. J. Biol. Chem. 2001, 276, 41394–41398. [Google Scholar] [CrossRef] [PubMed]
- Martinou, J.C.; Youle, R.J. Mitochondria in apoptosis: Bcl-2 family members and mitochondrial dynamics. Dev. Cell 2011, 21, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.; Hu, Y.F.; Song, J.; Yi, H.S.; Wang, L.; Yang, Y.Y.; Wang, Y.P.; Zhang, M.; Pan, M.H.; Lu, C. Effects of 10-hydroxycamptothecin on intrinsic mitochondrial pathway in silkworm BmN-SWU1 cells. Pestic. Biochem. Physiol. 2016, 127, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Shu, B.; Zhang, J.; Sethuraman, V.; Cui, G.; Yi, X.; Zhong, G. Transcriptome analysis of Spodoptera frugiperda Sf9 cells reveals putative apoptosis-related genes and a preliminary apoptosis mechanism induced by azadirachtin. Sci. Rep. 2017, 7, 13231. [Google Scholar] [CrossRef] [PubMed]
- Dorstyn, L.; Kumar, S. A cytochrome c-free fly apoptosome. Cell Death Differ. 2006, 13, 1049–1051. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Pan, M.H.; Sun, Z.Y.; Huang, S.J.; Yu, Z.S.; Liu, D.; Zhao, D.H.; Lu, C. The genomic underpinnings of apoptosis in the silkworm, Bombyx mori. BMC Genom. 2010, 11, 611. [Google Scholar] [CrossRef] [PubMed]









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Zhang, Z.; Shu, B.; Cui, G.; Zhong, G. Cytotoxic and Apoptotic Activity of the Novel Harmine Derivative ZC-14 in Sf9 Cells. Int. J. Mol. Sci. 2018, 19, 811. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030811
Zhang J, Zhang Z, Shu B, Cui G, Zhong G. Cytotoxic and Apoptotic Activity of the Novel Harmine Derivative ZC-14 in Sf9 Cells. International Journal of Molecular Sciences. 2018; 19(3):811. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030811
Chicago/Turabian StyleZhang, Jingjing, Zhijun Zhang, Benshui Shu, Gaofeng Cui, and Guohua Zhong. 2018. "Cytotoxic and Apoptotic Activity of the Novel Harmine Derivative ZC-14 in Sf9 Cells" International Journal of Molecular Sciences 19, no. 3: 811. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030811