Impact of Cationic and Neutral Gemini Surfactants on Conidia and Hyphal Forms of Aspergillus brasiliensis


Abstract
:1. Introduction
2. Results
2.1. Minimal Inhibitory Concentration
2.2. Viability of A. brasiliensis Conidia in the Presence of Microbiocides
2.3. Dry Weight and Ergosterol in Mycelium after GS Treatment
2.4. Ergosterol Binding Assay
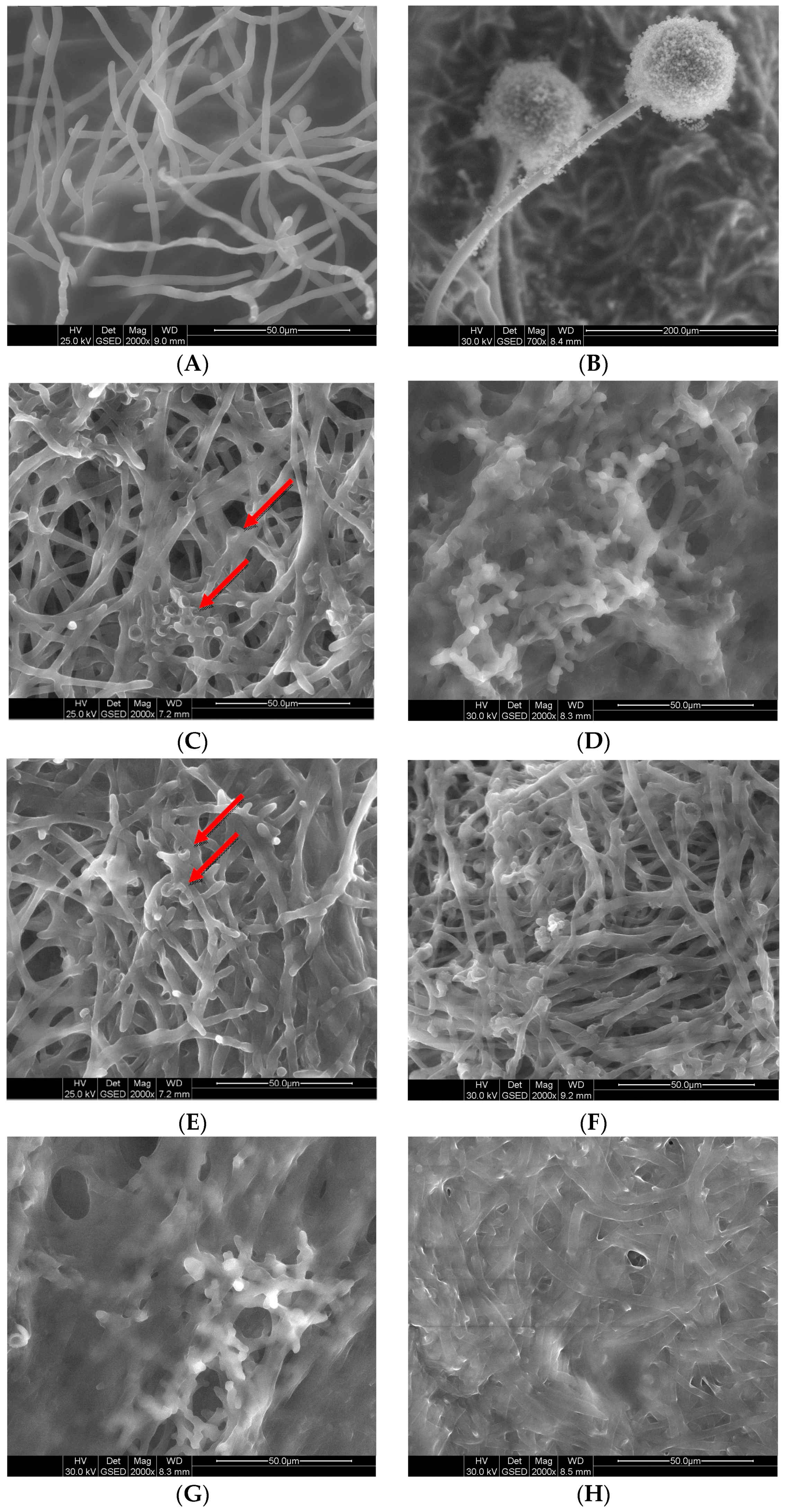
2.5. Effects of GS on Fungal Morphology by Scanning Electron Microscopy
3. Discussion
4. Materials and Methods
4.1. Strain and Growth Condition
4.2. Gemini Surfactants
4.3. Minimal Inhibitory Concentration (MIC)
4.4. Viability of A. brasiliensis Conidia in the Presence of Microbiocides
4.5. Dry Weight of Mycelium after GS Treatment
4.6. Content of Ergosterol in Mycelium after GS Treatment
4.7. Ergosterol Binding Assay
4.8. Effects of GS on Fungal Morphology by Scanning Electron Microscopy
4.9. Mathematical Calculation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| GS | Gemini surfactant |
| MIC | Minimal Inhibitory Concentration |
| HLB | Hydrophilic-Lipophilic Balance |
| ATCC | American Type Culture Collection |
References
- Hait, S.K.; Moulik, S.P. Gemini surfactants: A distinct class of self-assembling molecules. Curr. Sci. Bangalore 2002, 82, 1101–1111. [Google Scholar]
- Sekhon, B.S. Gemini (dimeric) surfactants. Resonance 2004, 9, 42–49. [Google Scholar] [CrossRef]
- Shukla, D.; Tyagi, V.K. Cationic gemini surfactants: A review. J. Oleo Sci. 2006, 55, 381–390. [Google Scholar] [CrossRef]
- Caillier, L.; Taffin de Givenchy, E.; Levy, R.; Vandenberghe, Y.; Geribaldi, S.; Guittard, F. Polymerizable semi-fluorinated gemini surfactants designed for antimicrobial materials. J. Colloid Interface Sci. 2009, 332, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Brycki, B.; Kowalczyk, I.; Koziróg, A. Synthesis, molecular structure, spectral properties and antifungal activity of polymethylene-α,ω-bis(N,N-dimethyl-N-dodecyloammonium bromides). Molecules 2011, 16, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Brycki, B.; Kowalczyk, I.; Szulc, A.; Kaczerewska, O.; Pakiet, M. Multifunctional gemini surfactants: Structure, synthesis, properties and application. In Application and Characterization of Surfactants, 1st ed.; Najjae, R., Ed.; InTech: Rijeka, Croatia, 2017; ISBN 978-953-51-3326-1. [Google Scholar]
- Sumitomo, T.; Maeda, T.; Nagamune, H.; Kourai, H. Bacterioclastic action of bis-quaternary ammonium compound against Escherichia coli. Biocontrol Sci. 2004, 9, 1–9. [Google Scholar] [CrossRef]
- Shirai, A.; Sumitomo, T.; Kurimoto, M.; Maeda, H.; Kourai, H. The mode of the antifungal activity of gemini-pyridinium salt against yeast. Biocontrol Sci. 2009, 14, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Obłąk, E.; Piecuch, A.; Dworniczek, E.; Olejniczak, T. The influence of biodegradable gemini surfactants, N,N′-bis(1-Decyloxy-1-Oxopronan-2-yl)-N,N,N′,N′ tetramethylpropane-1,3-diammonium dibromide and N,N′-bis(1-dodecyloxy-1-oxopronan-2-yl)-N,N,N′,N′-tetramethylethane-1,2-diammonium dibromide, on fungal biofilm and adhesion. J. Oleo Sci. 2015, 64, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Laska, U.; Wilk, A.; Maliszewska, I.; Syper, L. Novel glucose-derived gemini surfactants with a 1,1′-ethylenebisurea spacer: Preparation, thermotropic behavior, and biological properties. J. Surfactants Deterg. 2006, 9, 115–124. [Google Scholar] [CrossRef]
- Ding, Z.; Fang, S. Synthesis, surface and antimicrobial activities of novel cationic gemini surfactants. J. Surfactants Deterg. 2015, 18, 1051–1057. [Google Scholar] [CrossRef]
- Fothergill, A.W. Antifungal susceptibility testing: Clinical laboratory and standards institute (CLSI) Methods. In Interaction of Yeasts, Moulds and Antifungal Agent: How to Detect Resistance; Hall, G.S., Ed.; Springer: New York, NY, USA, 2012; p. 131. [Google Scholar]
- European Normative 1650+A1. Chemical Disinfectants and Antiseptics—Quantitative Suspension Test for the Evaluation of Fungicidal or Yeasticidal Activity of Chemical Disinfectants and Antiseptics Used in Food, Industrial, Domestic and Institutional Areas—Test Method and Requirements (Phase 2, Step 1); BSI: London, UK, 2010. [Google Scholar]
- Plumridge, A.; Hesse, S.J.A.; Watson, A.J.; Lowe, K.C.; Stratford, M.; Archer, D.B. The weak acid preservative sorbic acid inhibits conidial germination and mycelial growth of Aspergillus niger through intracellular acidification. Appl. Environ. Microbiol. 2004, 70, 3506–3511. [Google Scholar] [CrossRef] [PubMed]
- Van de Sande, W.W.; Tavakol, M.; van Vianen, W.; Bakker-Woudenberg, I.A. The effects of antifungal agents to conidial and hyphal forms of Aspergillus fumigatus. Med. Mycol. 2010, 48, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Wills, E.A.; Rebindo, M.R.; Perfect, J.R.; Del Poeta, M. New potential targets for antifungal development. Emerg. Ther. Targets 2000, 4, 265–296. [Google Scholar] [CrossRef]
- Chamilos, G.; Kontoyiannis, D.P. Update on antifungal drug resistance mechanisms of Aspergillus fumigatus. Drug Resist. Updates 2005, 8, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Pasanen, A.L.; Yli-Pietilä, K.; Pasanen, P.; Kalliokoski, P.; Tarhanen, J. Ergosterol content in various fungal species and biocontaminated building materials. Appl. Environ. Microbiol. 1999, 65, 138–142. [Google Scholar] [PubMed]
- Gutarowska, B.; Żakowska, Z. Elaboration and application of mathematical model for estimation of mould contamination of some building materials based on ergosterol content. Int. Biodeter. Biodeg. 2002, 49, 299–305. [Google Scholar] [CrossRef]
- Gutarowska, B.; Piotrowska, M. Methods of mycological analysis in buildings. Build. Environ. 2007, 42, 1843–1850. [Google Scholar] [CrossRef]
- Hippelein, M.; Rugamer, M. Ergosterol as an indicator of mould growth on building materials. Int. J. Hyg. Environ. Health 2004, 207, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Saxena, J.; Munimbazi, C.; Bullerman, L.B. Relationship of mould count, ergosterol and ochratoxin A production. Int. J. Food Microbiol. 2001, 71, 29–34. [Google Scholar] [CrossRef]
- Gutarowska, B.; Żakowska, Z. Estimation of fungal contamination of various plant materials with UV-determination of fungal ergosterol. Ann. Microbiol. 2010, 60, 415–422. [Google Scholar] [CrossRef]
- Suberkropp, K. Fungal growth, production, and sporulation during leaf decomposition in two streams. Appl. Environ. Microbiol. 2001, 67, 5063–5068. [Google Scholar] [CrossRef] [PubMed]
- Barajas-Aceves, M.; Hassan, M.; Tinoco, R.; Vazquez-Duhalt, R. Effect of pollutants on the ergosterol content as indicator of fungal biomass. J. Microbiol. Methods 2002, 50, 227–236. [Google Scholar] [CrossRef]
- Zill, G.; Engelhardt, G.; Wallnöfer, P.R. Determination of ergosterol as a measure of fungal growth using Si 60 HPLC. Z. Lebensm. Unters. Forsch. 1988, 187, 246–249. [Google Scholar] [CrossRef] [PubMed]
- Gutarowska, B.; Skóra, J.; Pielech-Przybylska, K. Evaluation of ergosterol content in the air of various environments. Aerobiologia 2015, 31, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.F.; Madsen, J.O. Determination of ergosterol on mouldy building materials using isotope dilution and gas chromatography-tandem mass spectrometry. J. Chromatogr. A 2000, 898, 227–234. [Google Scholar] [CrossRef]
- Hąc-Wydro, K.; Wydro, P.; Dynarowicz-Łątka, P. Interactions between Dialkyl-dimethylammonium bromides (DXDAB) and Sterols—A Monolayer Study. J. Colloid Interface Sci. 2005, 286, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.A.; Ahmad, I.; Cameotra, S.S. Phenyl aldehyde and propanoids exert multiple sites of action towards cell membrane and cell wall targetingergosterol in Candida albicans. AMB Express 2013, 3, 54–69. [Google Scholar] [CrossRef]
- Gutarowska, B. Moulds in biodeterioration of technical materials. Acta Univ. Lodz. Folia Biol. Oecol. 2014, 10, 27–39. [Google Scholar] [CrossRef]
- Balgavy, P.; Devinsky, F. Cut-off effect in biological activities of surfactant. Adv. Colloid Interface Sci. 1996, 66, 23–63. [Google Scholar] [CrossRef]
- Pisárčik, M.; Pupák, M.; Devinsky, F.; Almásy, L.; Tian, Q.; Bukovsky, M. Urea-based gemini surfactants: Synthesis, aggregation behavior and biological activity. Colloid Surface A 2016, 497, 385–396. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.; Xiao, H. Synthesis and antimicrobial characterization of novel l-lysine gemini surfactants pended with reactive groups. Tetrahedron Lett. 2008, 49, 1759–1761. [Google Scholar] [CrossRef]
- Colomer, A.; Pinazo, A.; Manresa, M.A.; Vinardell, M.P.; Mitjans, M.; Infante, M.R.; Perez, L. Cationic surfactants derived from lysine: Effect of their structure and chargé type on antimicrobial and hemolytic activities. J. Med. Chem. 2011, 54, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Obłąk, E.; Piecuch, A.; Krasowska, A.; Łuczyński, J. Antifungal activity of gemini Quaternery ammonium salts. Microbiol. Res. 2013, 168, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Koziróg, A.; Brycki, B. Monomeric and gemini surfactants as antimicrobial agents—Influence on environmental and reference strains. Acta Biochim. Pol. 2015, 62, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Shirai, A.; Sumitomo, T.; Yoshida, M.; Kaimura, T.; Nagamune, H.; Maeda, T.; Kourai, H. Synthesis and Biological Properties of Gemini Quaternary Ammonium Compounds, 5,5′-[2,2′-(α,ω-Polymethylnedicarbonyldioxy)diethyl]bis-(3-alkyl-4-methylthiazolium iodide) and 5,5′-[2,2′-(p-Phenylenedi-carbonyldioxy)diethyl]bis(3-alkyl-4-methylthiazolium bromide). Chem. Pharm. Bull. 2006, 54, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Lass-Flörl, C.; Nagl, M.; Speth, C.; Ulmer, H.; Dierich, M.P.; Würzner, R. Studies in vitro activities of voriconazole and itraconazole against Aspergillus hyphae using viability staining. Antimicrob. Agents Chemother. 2001, 45, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Guarro, J.; Llop, C.; Aguilar, C.; Pujol, I. Comparison of in vitro antifungal susceptibilities of conidia and hyphae of filamentous fungi. Antimicrob. Agents Chemother. 1997, 41, 2760–2762. [Google Scholar] [PubMed]
- Manavathu, E.K.; Cutright, J.; Chandrasekar, P.H. Comparative study of susceptibilities of germinated and ungerminated conidia of Aspergillus fumigatus to various antifungal agents. J. Clin. Microbiol. 1999, 37, 858–861. [Google Scholar] [PubMed]
- Rodríguez-Rojas, A.; Rodríguez-Beltrán, J.; Couce, A.; Blázquez, J. Antibiotics and antibiotic resistance: A bitter fight against evolution. Int. J. Med. Microbiol. 2013, 303, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Tezel, U.; Pavlostathis, S.G. Quaternary ammonium disinfectants: Microbial adaptation, degradation and ecology. Curr. Opin. Biotechnol. 2015, 33, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.D. Biocide use and antibiotic resistance: The relevance of laboratory findings to clinical and environmental situations. Lancet Infect. Dis. 2003, 3, 794–803. [Google Scholar] [CrossRef]
- Obłąk, E.; Gamian, A. The biological activity of quaternary ammonium salts. Postepy Hig. Med. Dosw. 2010, 64, 201–211. [Google Scholar]
- Valiante, V.; Macheleidt, J.; Föge, M.; Brakhage, A.A. The Aspergillus fumigatus cell wall integrity signaling pathway: Drug target, compensatory pathways, and virulence. Front. Microbiol. 2015, 6, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Campoy, S.; Adrio, J.L. Antifungals. Biochem. Pharmacol. 2017, 133, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Gessner, M.O. Ergosterol as a measure of fungal biomass. In Methods to Study Litter Decomposition—A Practical Guide, 1st ed.; Graça, M.A., Bärlocher, F., Gessner, M.O., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 189–195. ISBN 978-1-4020-3466-4. [Google Scholar]
- Pawiroharsono, S.; Naji, B.; Bonaly, R.; Tonetti, F.; Chasseboeuf, C.; Richter, J.P. Permeability and membrane sterol distribution in Saccharomyces uvarum and Kluyveromyces bulgaricus grown in presence of polyoxyalkylene glycol-oleic acid condensates. Appl. Microbiol. Biotechnol. 1987, 27, 181–185. [Google Scholar] [CrossRef]
- Mille-Lindblom, C.; von Wachenfeldt, E.; Tranvik, L.J. Ergosterol as a measure of living fungal biomass: Persistence in environmental samples after fungal death. J. Microbiol. Methods 2004, 59, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Koziróg, A.; Brycki, B.; Michlewska, S. Changes in Aspergillus niger mycelium after N,N-bis(3-aminopropyl)dodecylamine treatment. Postępy Mikrobiol. 2017, 3. in press. [Google Scholar]
- Devinsky, F.; Lacko, I.; Krasnec, L. Synthesis, IR and 1H-NMR spectra of N,N,N′,N′-tetraalkyl-l,6-hexanediamine dioxides. Collect. Czech. Chem. Commun. 1979, 44, 773–780. [Google Scholar] [CrossRef]
- Campanac, C.; Pineau, L.; Payard, A.; Baziard-Mouysset, G.; Roques, C. Interaction between biocide cationic agents and bacterial biofilms. Antimicrob. Agents Chemother. 2002, 46, 1469–1474. [Google Scholar] [CrossRef] [PubMed]
- Seitz, L.M.; Sauer, D.B.; Burroughs, R.; Mohr, H.E.; Hubbard, J.D. Ergosterol as a measure of fungal growth. Phytopathology 1979, 69, 1202–1203. [Google Scholar] [CrossRef]
- Escalante, A.; Gattuso, M.; Perez, P.; Zacchino, S. Evidence for the mechanism of action of the antifungal phytolaccoside B isolated from Phytolacca tetramera Hauman. J. Nat. Prod. 2008, 71, 1720–1725. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biocide | Conidia | Mycelium |
|---|---|---|
| C5 | 0.12 | 0.31 |
| C6 | 0.12 | 0.31 |
| A5 | 0.38 | 30 |
| A6 | 0.38 | 25 |
| Biocide Concentration (mM) | Biocide | Time (h) | ||||
|---|---|---|---|---|---|---|
| 4 | 8 | 24 | 48 | |||
| control | 0.00 | K | 6.20 ± 0.05 | 6.18 ± 0.10 | mycelium | Mycelium |
| ½ MIC | 0.06 | C5 | 5.93 ± 0.53 | 5.46 ± 0.15 # | mycelium | Mycelium |
| 0.06 | C6 | 5.51 ± 0.37 # | 4.18 ± 0.11 # | mycelium | Mycelium | |
| 0.19 | A5 | 5.90 ± 0.60 | 5.87 ± 0.22 # | mycelium | Mycelium | |
| 0.19 | A6 | 5.92 ± 0.10 | 5.71 ± 0.20 # | mycelium | Mycelium | |
| MIC | 0.12 | C5 | 4.49 ± 0.03 # | 4.72 ± 0.13 # | nd | nd |
| 0.12 | C6 | 3.87 ± 0.02 # | 2.91 ± 0.23 # | nd | nd | |
| 0.38 | A5 | 5.11 ± 0.03 # | 2.73 ± 0.16 # | 1.64 ± 0.05 # | nd | |
| 0.38 | A6 | 4.92 ± 0.03 # | 4.20 ± 0.31 # | 1.04 ± 0.02 # | nd | |
| 2 MIC | 0.24 | C5 | 2.91 ± 0.10 # | 1.85 ± 0.08 # | nd | nd |
| 0.24 | C6 | 2.38 ± 0.24 # | nd | nd | nd | |
| 0.76 | A5 | 4.30 ± 0.32 # | 2.97 ± 0.15 # | 1.25 ± 0.01 # | nd | |
| 0.76 | A6 | 3.08 ± 0.28 # | 2.48 ± 0.21 # | nd | nd | |
| Biocide Concentration (mM) | Biocide | Correlation Coefficient r | |
|---|---|---|---|
| K | 0.00 | - | 0.999 |
| ½ MIC | 0.06 | C5 | 0.987 |
| 0.06 | C6 | 0.947 | |
| 0.19 | A5 | 0.999 | |
| 0.19 | A6 | 0.966 | |
| MIC | 0.12 | C5 | 0.958 |
| 0.12 | C6 | 0.958 | |
| 0.38 | A5 | −0.869 | |
| 0.38 | A6 | −0.750 | |
| 2 MIC | 0.24 | C5 | 0.996 |
| 0.24 | C6 | 0.990 | |
| 0.76 | A5 | 0.808 | |
| 0.76 | A6 | 0.837 | |
| Biocide | Control | With Ergosterol |
|---|---|---|
| C5 | 0.12 | 0.36 |
| C6 | 0.12 | 0.36 |
| A5 | 0.38 | 0.19 |
| A6 | 0.38 | 0.19 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koziróg, A.; Brycki, B.; Pielech-Przybylska, K. Impact of Cationic and Neutral Gemini Surfactants on Conidia and Hyphal Forms of Aspergillus brasiliensis. Int. J. Mol. Sci. 2018, 19, 873. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030873
Koziróg A, Brycki B, Pielech-Przybylska K. Impact of Cationic and Neutral Gemini Surfactants on Conidia and Hyphal Forms of Aspergillus brasiliensis. International Journal of Molecular Sciences. 2018; 19(3):873. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030873
Chicago/Turabian StyleKoziróg, Anna, Bogumił Brycki, and Katarzyna Pielech-Przybylska. 2018. "Impact of Cationic and Neutral Gemini Surfactants on Conidia and Hyphal Forms of Aspergillus brasiliensis" International Journal of Molecular Sciences 19, no. 3: 873. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030873