Kluyveromyces marxianus, an Attractive Yeast for Ethanolic Fermentation in the Presence of Imidazolium Ionic Liquids

 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Yeast Morphology in Ionic Liquid (IL)
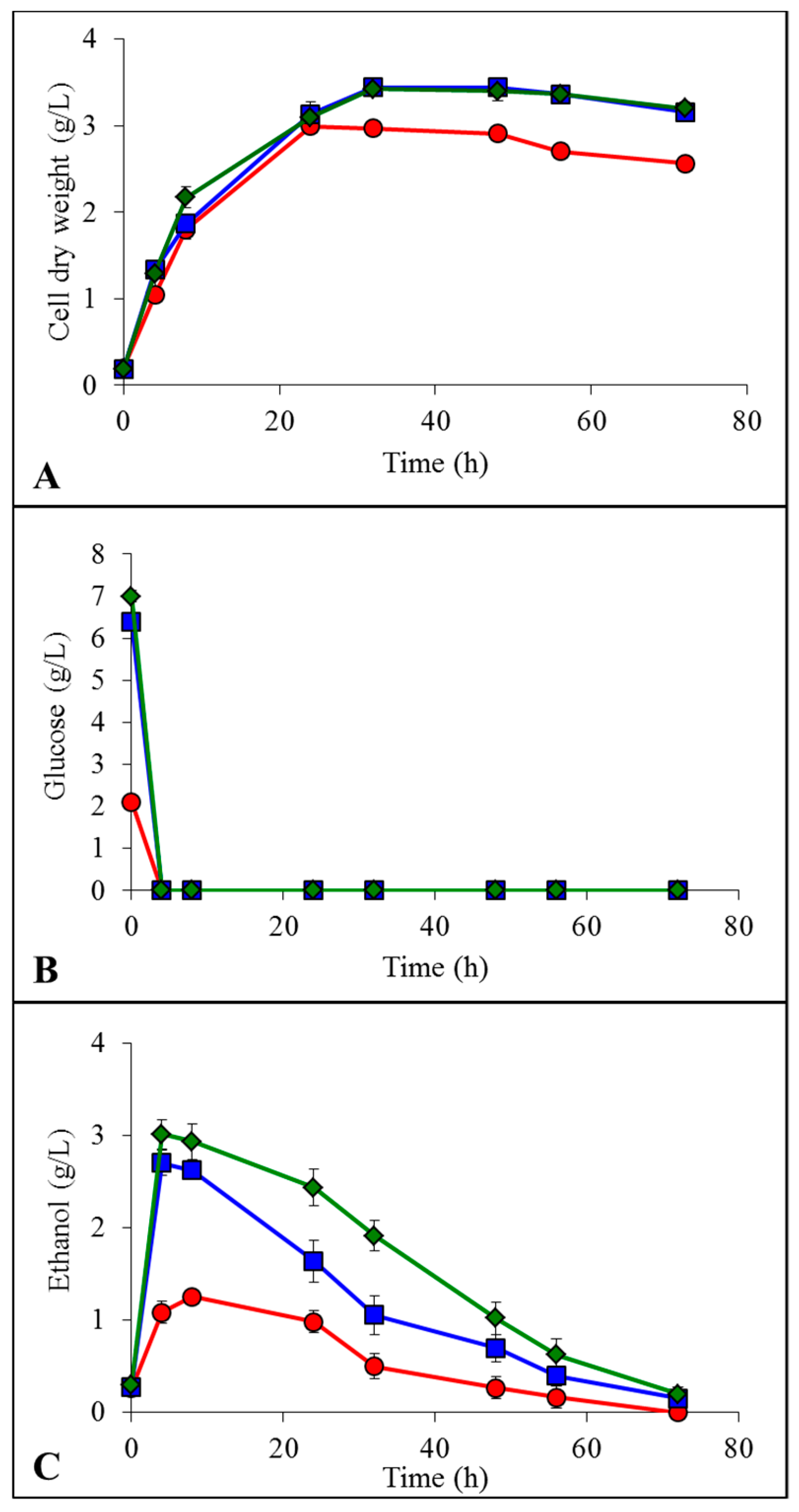
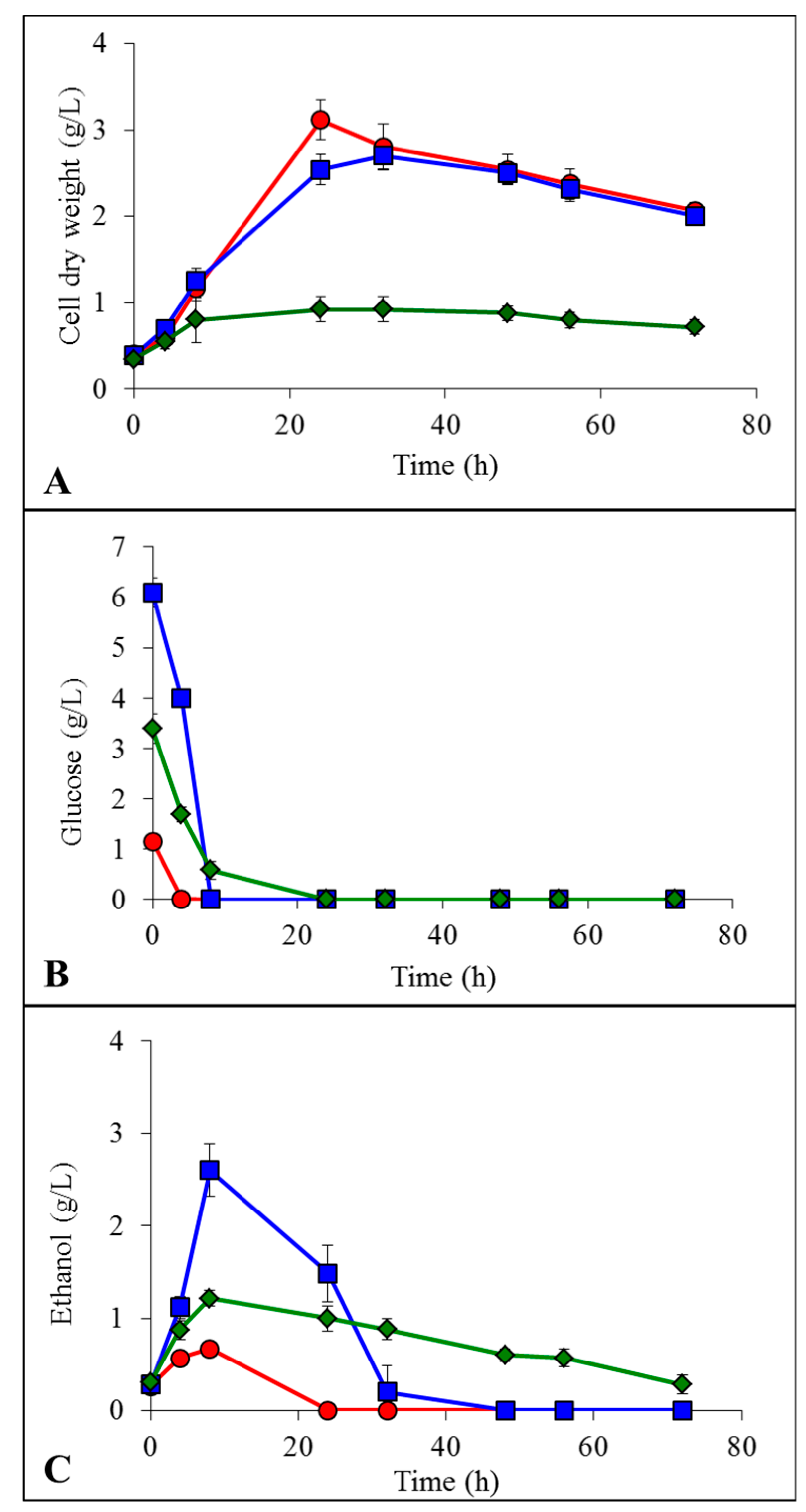
2.2. Growth, Glucose Consumption and Ethanol Formation in IL
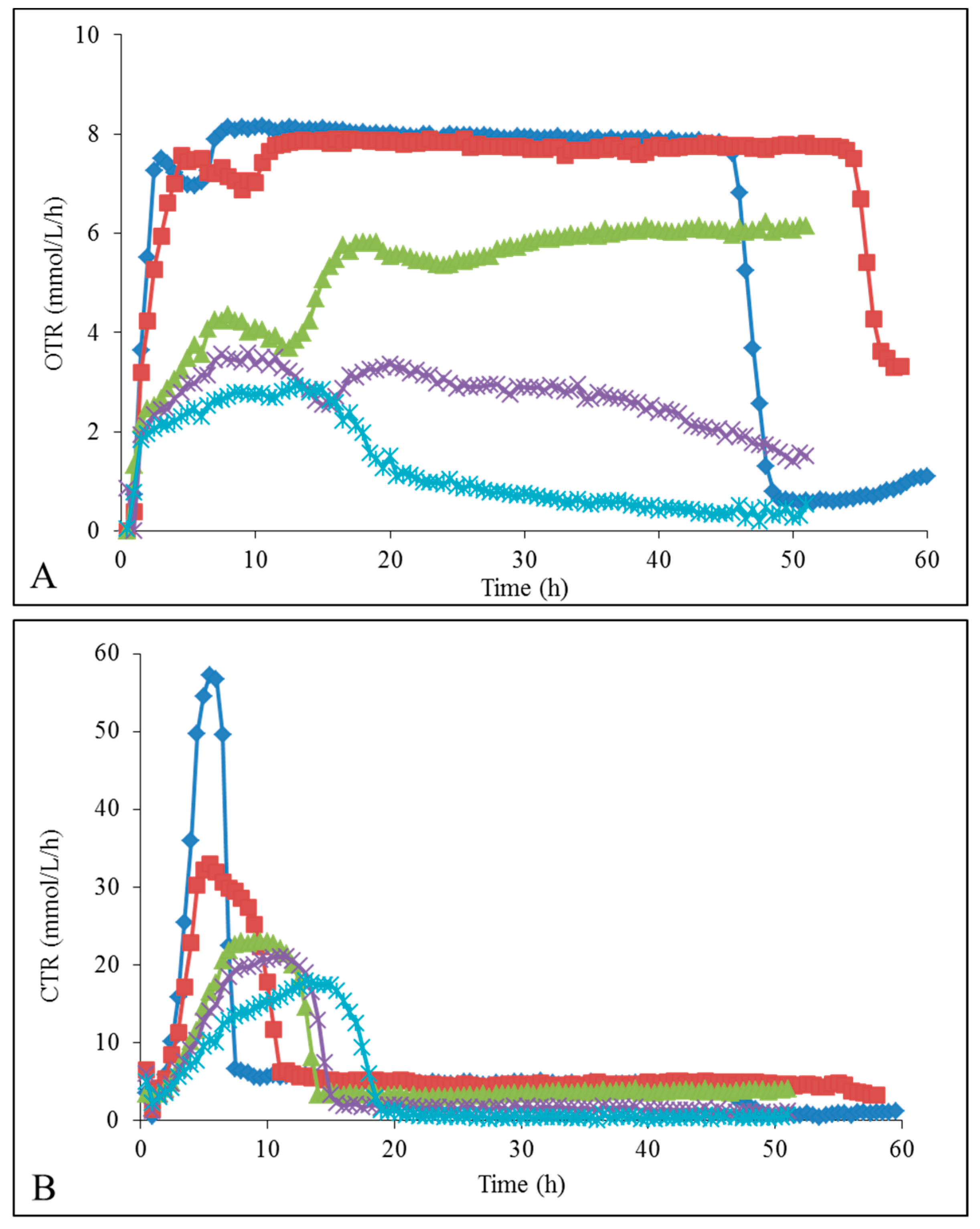
2.3. Oxygen Transfer Rate (OTR) and Carbon Dioxide Transfer Rate (CTR) Profiles
2.4. Application to Lignocellulosic Biomasses (LCBs)
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Strain and Culture Conditions
4.3. Lignocellulosic Biomasses (LCBs)
4.4. Morphology Observations
4.5. Glucose and Ethanol Quantification
4.6. Oxygen Transfer Rate (OTR) and Carbon Dioxide Transfer Rate (CTR) Measurements
4.7. IL Pretreatment of LCBs
4.8. Enzymatic Hydrolysis of LCBs
4.9. Fermentation of Enzymatically Hydrolysed LCBs
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CDW | Cell Dry Weight |
| CTR | Carbon Dioxide Transfer Rate |
| [Emim][OAc] | 1-Ethyl-3-methylimidazolium Acetate |
| [Emim][MeO(H)PO2] | 1-Ethyl-3-methylimidazolium Methylphosphonate |
| IL | Ionic Liquid |
| LCB | Ligno Cellulosic Biomass |
| OTR | Oxygen Transfer Rate |
| RAMOS | Respiratory Activity Monitoring System |
| SEM | Scanning Electron Microscopy |
| SHF | Separate Hydrolysis and Fermentation |
| YM | Yeast-Malt |
| YMD | Yeast-Malt-Dextrose |
References
- Balat, M. Production of bioethanol from lignocellulosic materials via the biochemical pathway: A review. Energy Convers. Manag. 2011, 52, 858–875. [Google Scholar] [CrossRef]
- Valdivia, M.; Galan, J.L.; Laffarga, J.; Ramos, J.-L. Biofuels 2020: Biorefineries based on lignocellulosic materials. Microb. Biotechnol. 2016, 9, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.L.; García-Lorente, F.; Valdivia, M.; Duque, E. Green biofuels and bioproducts: Bases for sustainability analysis. Microb. Biotechnol. 2017, 10, 1111–1113. [Google Scholar] [CrossRef] [PubMed]
- Badgujar, K.C.; Bhanage, B.M. Factors governing dissolution process of lignocellulosic biomass in ionic liquid: Current status, overview and challenges. Bioresour. Technol. 2015, 178, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Jönsson, L.J.; Martín, C. Pretreatment of lignocellulose: Formation of inhibitory by-products and strategies for minimizing their effects. Bioresour. Technol. 2016, 199, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhang, R.; Cheng, T.; Guo, J.; Xian, M.; Liu, H. Imidazolium-based ionic liquids for cellulose pretreatment: Recent progresses and future perspectives. Appl. Microbiol. Biotechnol. 2017, 101, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Sitepu, I.R.; Shi, S.; Simmons, B.A.; Singer, S.W.; Boundy-Mills, K.; Simmons, C.W. Yeast tolerance to the ionic liquid 1-ethyl-3-methylimidazolium acetate. FEMS Yeast Res. 2014, 14, 1286–1294. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, Q.; Bottoms, S.; Hinchman, L.; McIlwain, S.; Li, S.; Myers, C.L.; Boone, C.; Coon, J.J.; Hebert, A.; Sato, T.K.; et al. Mechanism of imidazolium ionic liquids toxicity in Saccharomyces cerevisiae and rational engineering of a tolerant, xylose fermenting strain. Microb. Cell Fact. 2016, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, M.; Datta, S.; Dibble, D.C.; Tamrakar, P.R.; Benke, P.I.; Li, C.; Singh, S.; Sale, K.L.; Adams, P.D.; Keasling, J.D.; et al. Impact of ionic liquid pretreated plant biomass on Saccharomyces cerevisiae growth and biofuel production. Green Chem. 2011, 13, 2743–2749. [Google Scholar] [CrossRef]
- Santos, A.G.; Ribeiro, B.D.; Alviano, D.S.; Coelho, M.A.Z. Toxicity of ionic liquids toward microorganisms interesting to the food industry. RSC Adv. 2014, 4, 37157–37163. [Google Scholar] [CrossRef]
- Isik, M.; Sardon, H.; Mecerreyes, D. Ionic liquids and cellulose: Dissolution, chemical modification and preparation of new cellulosic materials. Int. J. Mol. Sci. 2014, 15, 11922–11940. [Google Scholar] [CrossRef] [PubMed]
- Olivier-Bourbigou, H.; Magna, L.; Morvan, D. Ionic liquids and catalysis: Recent progress from knowledge to applications. Appl. Catal. A Gen. 2010, 373, 1–56. [Google Scholar] [CrossRef]
- Auxenfans, T.; Husson, E.; Sarazin, C. Simultaneous pretreatment and enzymatic saccharification of (ligno)celluloses in aqueous-ionic liquid media: A compromise. Biochem. Eng. J. 2017, 117, 77–86. [Google Scholar] [CrossRef]
- Fonseca, G.G.; Heinzle, E.; Wittmann, C.; Gombert, A.K. The yeast Kluyveromyces marxianus and its biotechnological potential. Appl. Microbiol. Biotechnol. 2008, 79, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, G.G.; de Carvalho, N.M.; Gombert, A.K. Growth of the yeast Kluyveromyces marxianus CBS 6556 on different sugar combinations as sole carbon and energy source. Appl. Microbiol. Biotechnol. 2013, 97, 5055–5067. [Google Scholar] [CrossRef] [PubMed]
- Radecka, D.; Mukherjee, V.; Mateo, R.Q.; Stojiljkovic, M.; Foulquié-Moreno, M.R.; Thevelein, J.M. Looking beyond Saccharomyces: The potential of non-conventional yeast species for desirable traits in bioethanol fermentation. FEMS Yeast Res. 2015, 15, fov053. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, N.; Husson, E.; Jacquard, C.; Wewetzer, S.; Büchs, J.; Sarazin, C.; Gosselin, I. Impact of two ionic liquids, 1-ethyl-3-methylimidazolium acetate and 1-ethyl-3-methylimidazolium methylphosphonate, on Saccharomyces cerevisiae: Metabolic, physiologic, and morphological investigations. Biotechnol. Biofuels 2015, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Anderlei, T.; Büchs, J. Device for sterile online measurement of the oxygen transfer rate in shaking flasks. Biochem. Eng. J. 2001, 7, 157–162. [Google Scholar] [CrossRef]
- Meier, K.; Klöckner, W.; Bonhage, B.; Antonov, E.; Regestein, L.; Büchs, J. Correlation for the maximum oxygen transfer capacity in shake flasks for a wide range of operating conditions and for different culture media. Biochem. Eng. J. 2016, 109, 228–235. [Google Scholar] [CrossRef]
- Husson, E.; Buchoux, S.; Avondo, C.; Cailleu, D.; Djellab, K.; Gosselin, I.; Wattraint, O.; Sarazin, C. Enzymatic hydrolysis of ionic liquid-pretreated celluloses: Contribution of CP-MAS 13C NMR and SEM. Bioresour. Technol. 2011, 102, 7335–7342. [Google Scholar] [CrossRef] [PubMed]
- Auxenfans, T.; Buchoux, S.; Larcher, D.; Husson, G.; Husson, E.; Sarazin, C. Enzymatic saccharification and structural properties of industrial wood sawdust: Recycled ionic liquids pretreatments. Energy Convers. Manag. 2014, 88, 1094–1103. [Google Scholar] [CrossRef]
- Petersen, R.C. The chemical composition of wood. In The Chemistry of Solid Wood; Rowell, R.M., Ed.; American Chemical Society: Washington, DC, USA, 1984; Chapter 2; pp. 57–126. [Google Scholar]
- Nakashima, K.; Yamaguchi, K.; Taniguchi, N.; Arai, S.; Yamada, R.; Katahira, S.; Ishida, N.; Takahashi, H.; Ogino, C.; Kondo, A. Direct bioethanol production from cellulose by the combination of cellulase-displaying yeast and ionic liquid pretreatment. Green Chem. 2011, 13, 2948–2953. [Google Scholar] [CrossRef]
- Blumer-Schuette, S.E.; Brown, S.D.; Sander, K.B.; Bayer, E.A.; Kataeva, I.; Zurawski, J.V.; Conway, J.M.; Adams, M.W.W.; Kelly, R.M. Thermophilic lignocellulose deconstruction. FEMS Microbiol. Rev. 2014, 38, 393–448. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kondo, A. Cell surface engineering of industrial microorganisms for biorefining applications. Biotechnol. Adv. 2015, 33, 1403–1411. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.P.; Simmons, C.W.; Claypool, J.; Jabusch, L.; Burd, H.; Hadi, M.Z.; Simmons, B.A.; Singer, S.W.; VanderGheynst, J.S. Thermophilic enrichment of microbial communities in the presence of the ionic liquid 1-ethyl-3-methylimidazolium acetate. J. Appl. Microbiol. 2012, 113, 1362–1370. [Google Scholar] [CrossRef] [PubMed]
- Frederix, M.; Hütter, K.; Leu, J.; Batth, T.S.; Turner, W.J.; Rüegg, T.L.; Blanch, H.W.; Simmons, B.A.; Adams, P.D.; Keasling, J.D.; et al. Development of a native Escherichia coli induction system for ionic liquid tolerance. PLoS ONE 2014, 9, e101115. [Google Scholar] [CrossRef] [PubMed]
- Ruegg, T.L.; Kim, E.-M.; Simmons, B.A.; Keasling, J.D.; Singer, S.W.; Soon Lee, T.; Thelen, M.P. An auto-inducible mechanism for ionic liquid resistance in microbial biofuel production. Nat. Commun. 2014, 5, 3490. [Google Scholar] [CrossRef] [PubMed]
- Portillo, M.D.C.; Saadeddin, A. Recent trends in ionic liquid (IL) tolerant enzymes and microorganisms for biomass conversion. Crit. Rev. Biotechnol. 2015, 35, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Simmons, B.A.; Singer, S.W.; Thelen, M.P.; VanderGheynst, J.S. Ionic liquid-tolerant microorganisms and microbial communities for lignocellulose conversion to bioproducts. Appl. Microbiol. Biotechnol. 2016, 100, 10237–10249. [Google Scholar] [CrossRef] [PubMed]
- Lesage, G.; Bussey, H. Cell wall assembly in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2006, 70, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-P.; Zong, M.-H.; Linhardt, R.J.; Lou, W.-Y.; Li, N.; Huang, C.; Wu, H. Mechanistic insights into the effect of imidazolium ionic liquid on lipid production by Geotrichum fermentans. Biotechnol. Biofuels 2016, 9, 266. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.S.A.; Lee, S.-H.; Endo, T.; Bon, E.P.S. Major improvement in the rate and yield of enzymatic saccharification of sugarcane bagasse via pretreatment with the ionic liquid 1-ethyl-3-methylimidazolium acetate ([Emim] [Ac]). Bioresour. Technol. 2011, 102, 10505–10509. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Knierim, B.; Manisseri, C.; Arora, R.; Scheller, H.V.; Auer, M.; Vogel, K.P.; Simmons, B.A.; Singh, S. Comparison of dilute acid and ionic liquid pretreatment of switchgrass: Biomass recalcitrance, delignification and enzymatic saccharification. Bioresour. Technol. 2010, 101, 4900–4906. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; He, Y.-C.; Xian, M.; Jun, G.; Xu, X.; Yang, J.-M.; Li, L.-Z. Improving enzymatic hydrolysis of wheat straw using ionic liquid 1-ethyl-3-methyl imidazolium diethyl phosphate pretreatment. Bioresour. Technol. 2009, 100, 3570–3575. [Google Scholar] [CrossRef] [PubMed]
- Binder, J.B.; Raines, R.T. Fermentable sugars by chemical hydrolysis of biomass. Proc. Natl. Acad. Sci. USA 2010, 107, 4516–4521. [Google Scholar] [CrossRef] [PubMed]
- Soudham, V.P.; Raut, D.G.; Anugwom, I.; Bradberg, T.; Larsson, C.; Mikkola, J.-P. Coupled enzymatic hydrolysis and ethanol fermentation: Ionic liquid pretreatment for enhanced yields. Biotechnol. Biofuels 2015, 8, 135. [Google Scholar] [CrossRef] [PubMed]
- Lienqueo, M.E.; Ravanal, M.C.; Pezoa-Conte, R.; Cortínez, V.; Martínez, L.; Niklitschek, T.; Salazar, O.; Carmona, R.; García, A.; Hyvärinen, S.; et al. Second generation bioethanol from Eucalyptus globulus Labill and Nothofagus pumilio: Ionic liquid pretreatment boosts the yields. Ind. Crops Prod. 2016, 80, 148–155. [Google Scholar] [CrossRef]
- Anderlei, T.; Zang, W.; Papaspyrou, M.; Büchs, J. Online respiration activity measurement (OTR, CTR, RQ) in shake flasks. Biochem. Eng. J. 2004, 17, 187–194. [Google Scholar] [CrossRef]

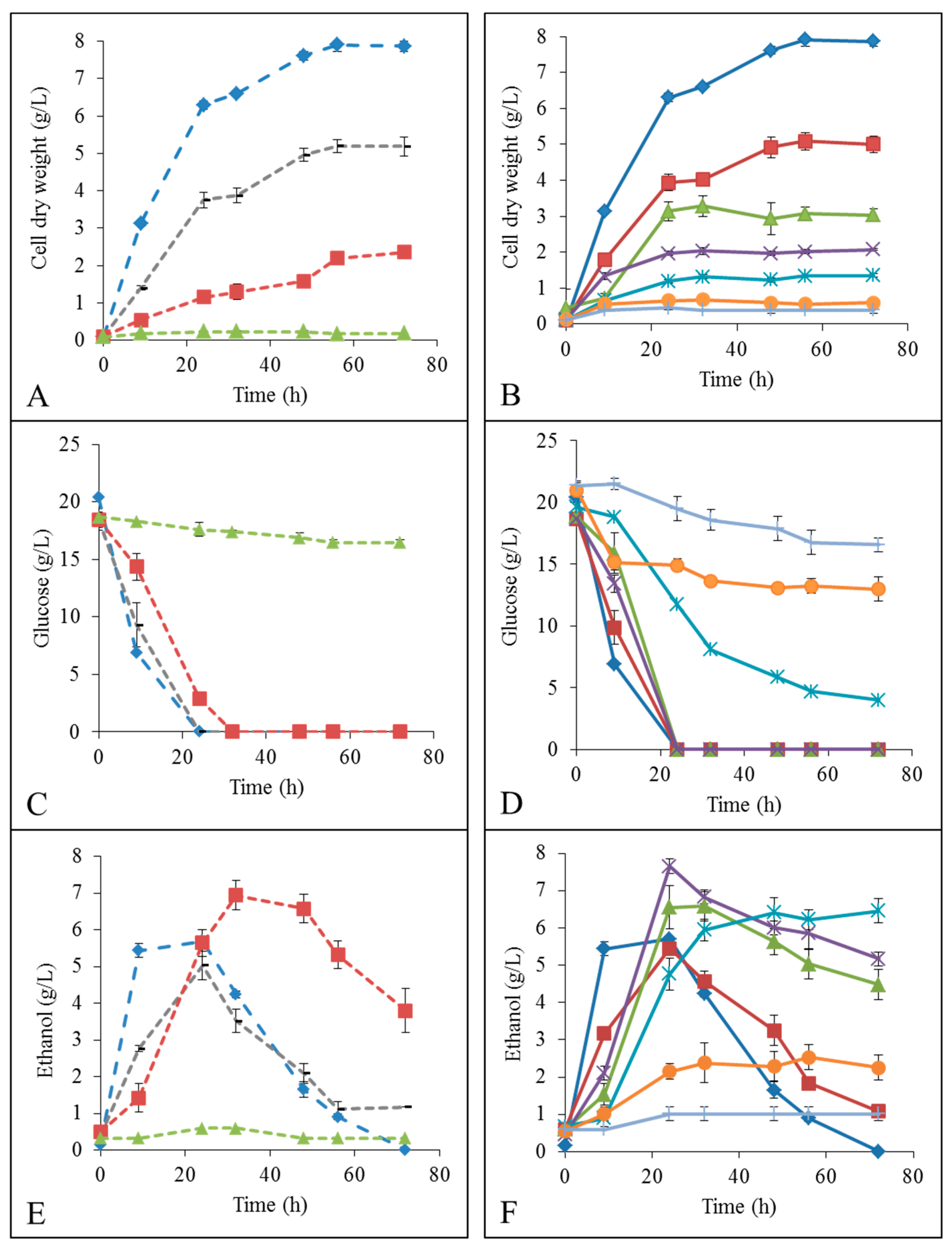
 YMD;
YMD;  YMD + 0.5% ionic liquid (IL);
YMD + 0.5% ionic liquid (IL);  YMD + 1% IL;
YMD + 1% IL;  YMD + 2% IL;
YMD + 2% IL;  YMD + 3% IL;
YMD + 3% IL;  YMD + 4% IL;
YMD + 4% IL;  YMD + 5% IL and
YMD + 5% IL and  YMD + 6% IL (means ± S.D.). Dashed lines for [Emim][OAc] (A,C,E) and full lines for [Emim][MeO(H)PO2] (B,D,F).
YMD; YMD + 0.5% ionic liquid (IL); YMD + 1% IL; YMD + 2% IL; YMD + 3% IL; YMD + 4% IL; YMD + 5% IL and YMD + 6% IL (means ± S.D.). Dashed lines for [Emim][OAc] (A,C,E) and full lines for [Emim][MeO(H)PO2] (B,D,F).
YMD + 6% IL (means ± S.D.). Dashed lines for [Emim][OAc] (A,C,E) and full lines for [Emim][MeO(H)PO2] (B,D,F).
YMD; YMD + 0.5% ionic liquid (IL); YMD + 1% IL; YMD + 2% IL; YMD + 3% IL; YMD + 4% IL; YMD + 5% IL and YMD + 6% IL (means ± S.D.). Dashed lines for [Emim][OAc] (A,C,E) and full lines for [Emim][MeO(H)PO2] (B,D,F). YMD; YMD + 1% [Emim][MeO(H)PO2]; YMD + 2% [Emim][MeO(H)PO2]; YMD + 3% [Emim][MeO(H)PO2]; YMD + 4% [Emim][MeO(H)PO2]; YMD + 5% [Emim][MeO(H)PO2] and YMD + 6% [Emim][MeO(H)PO2]. Error bars are not represented to avoid overloading the figure.
YMD; YMD + 1% [Emim][MeO(H)PO2]; YMD + 2% [Emim][MeO(H)PO2]; YMD + 3% [Emim][MeO(H)PO2]; YMD + 4% [Emim][MeO(H)PO2]; YMD + 5% [Emim][MeO(H)PO2] and YMD + 6% [Emim][MeO(H)PO2]. Error bars are not represented to avoid overloading the figure.
YMD; YMD + 1% [Emim][MeO(H)PO2]; YMD + 2% [Emim][MeO(H)PO2]; YMD + 3% [Emim][MeO(H)PO2]; YMD + 4% [Emim][MeO(H)PO2]; YMD + 5% [Emim][MeO(H)PO2] and YMD + 6% [Emim][MeO(H)PO2]. Error bars are not represented to avoid overloading the figure.
YMD; YMD + 1% [Emim][MeO(H)PO2]; YMD + 2% [Emim][MeO(H)PO2]; YMD + 3% [Emim][MeO(H)PO2]; YMD + 4% [Emim][MeO(H)PO2]; YMD + 5% [Emim][MeO(H)PO2] and YMD + 6% [Emim][MeO(H)PO2]. Error bars are not represented to avoid overloading the figure.
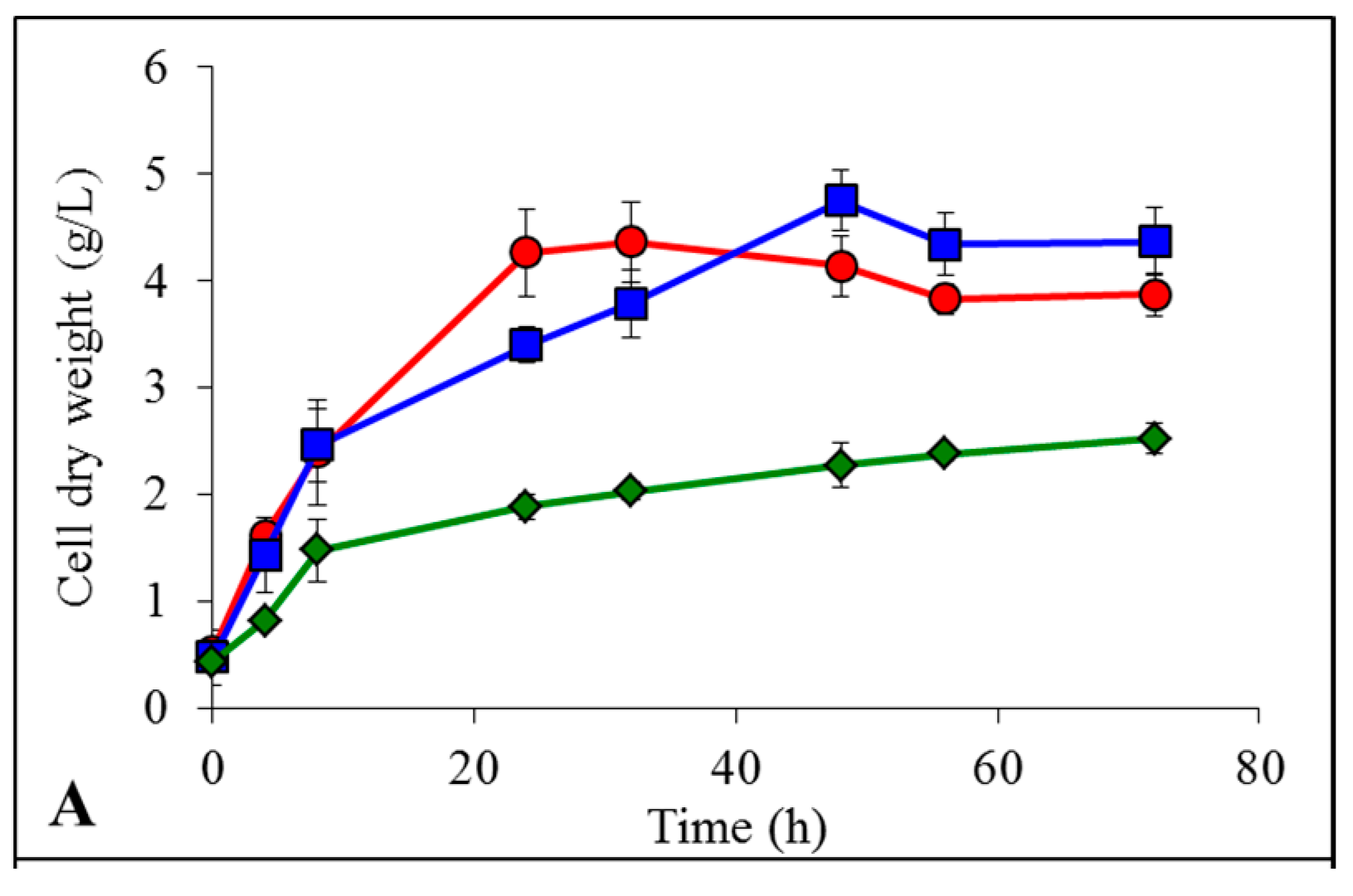
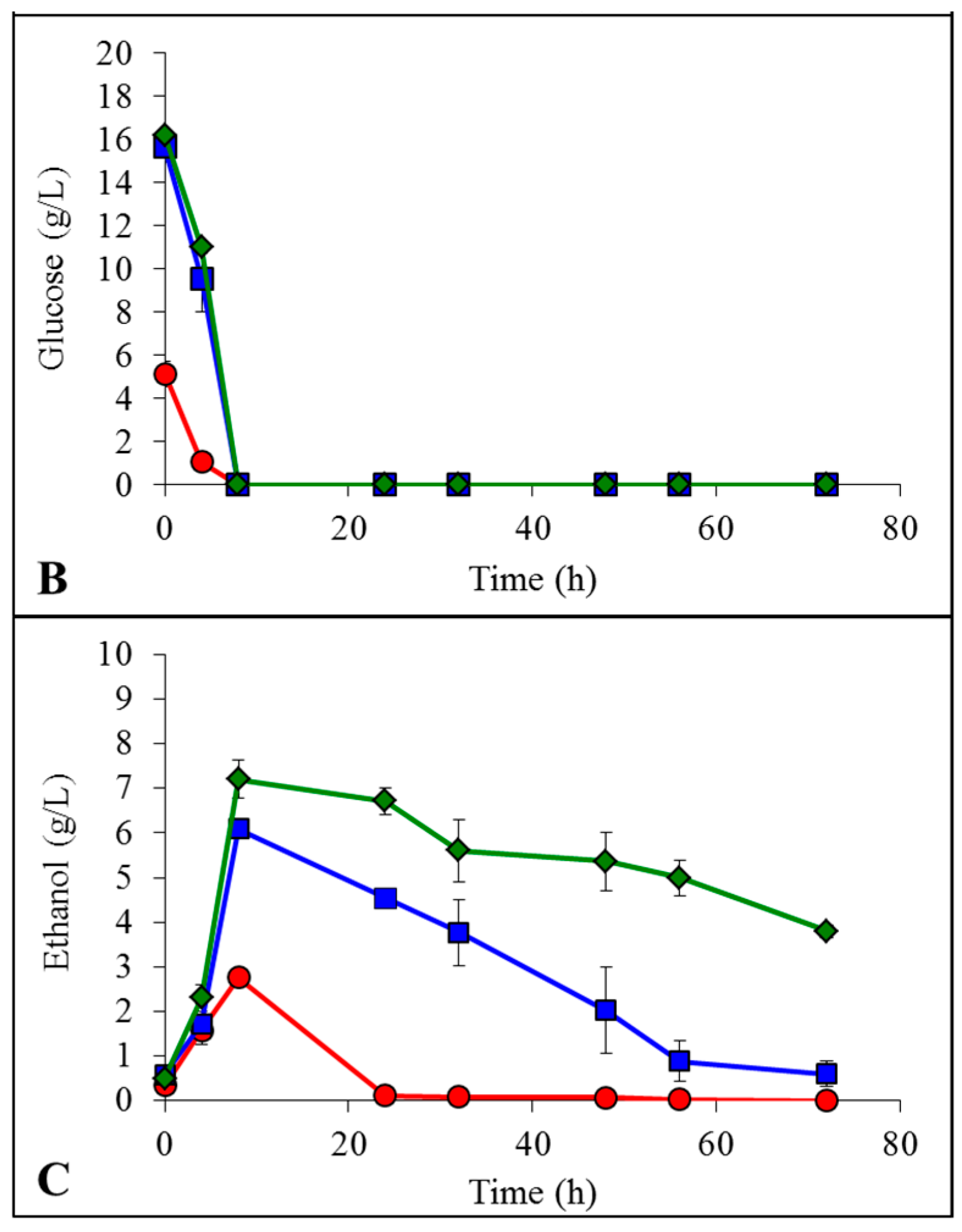
 non-pretreated,
non-pretreated,  pretreated with [Emim][OAc] and
pretreated with [Emim][OAc] and  pretreated with [Emim][MeO(H)PO2].
non-pretreated, pretreated with [Emim][OAc] and pretreated with [Emim][MeO(H)PO2].
pretreated with [Emim][MeO(H)PO2].
non-pretreated, pretreated with [Emim][OAc] and pretreated with [Emim][MeO(H)PO2].
 non-pretreated, pretreated with [Emim][OAc] and pretreated with [Emim][MeO(H)PO2].
non-pretreated, pretreated with [Emim][OAc] and pretreated with [Emim][MeO(H)PO2].
non-pretreated, pretreated with [Emim][OAc] and pretreated with [Emim][MeO(H)PO2].
non-pretreated, pretreated with [Emim][OAc] and pretreated with [Emim][MeO(H)PO2]. non-pretreated, pretreated with [Emim][OAc] and pretreated with [Emim][MeO(H)PO2].
non-pretreated, pretreated with [Emim][OAc] and pretreated with [Emim][MeO(H)PO2].
non-pretreated, pretreated with [Emim][OAc] and pretreated with [Emim][MeO(H)PO2].
non-pretreated, pretreated with [Emim][OAc] and pretreated with [Emim][MeO(H)PO2].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Pretreatment | Residue Recovery (% w/w) | Cellulose | Lignin | Xylose | Arabinose | Extractives |
|---|---|---|---|---|---|---|---|
| Spruce sawdust | None | 100 | 55.4 ± 1.3 | 28.7 ± 1.0 | 4.2 ± 0.4 | 1.4 ± 0.2 | 3.3 ± 1.0 |
| [Emim][OAc] | 85.8 ± 1.2 | 58.2 ± 0.4 | 24.6 ± 0.5 | 3.7 ± 0.8 | 1.2 ± 0.1 | 2.0 ± 0.3 | |
| [Emim][MeO(H)PO2] | 85.1 ± 2.3 | 56.6 ± 1.6 | 23.2 ± 1.4 | 3.5 ± 1.0 | 1.1 ± 0.2 | 2.6 ± 0.8 | |
| Oak sawdust | None | 100 | 44.7 ± 0.3 | 26.7 ± 0.8 | 14.8 ± 1.3 | 1.2 ± 0.1 | 4.6 ± 1.1 |
| [Emim][OAc] | 85.3 ± 0.8 | 46.5 ± 0.7 | 23.5 ± 1.8 | 18.5 ± 1.3 | 1.1 ± 0.1 | 2.5 ± 0.7 | |
| [Emim][MeO(H)PO2] | 89.2 ± 1.1 | 46.3 ± 1.6 | 25.8 ± 1.6 | 16.3 ± 1.1 | 1.1 ± 0.1 | 3.8 ± 0.8 |
| Pretreatment | None | [Emim][OAc] | [Emim][MeO(H)PO2] |
|---|---|---|---|
| Long fiber cellulose | 24.9 ± 0.5 | 68.2 ± 1.6 | 73.5 ± 1.7 |
| Spruce sawdust | 25.5 ± 1.7 | 49.3 ± 0.8 | 54.3 ± 3.5 |
| Oak sawdust | 11.6 ± 0.7 | 59.3 ± 2.9 | 32.1 ± 3.0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehmood, N.; Alayoubi, R.; Husson, E.; Jacquard, C.; Büchs, J.; Sarazin, C.; Gosselin, I. Kluyveromyces marxianus, an Attractive Yeast for Ethanolic Fermentation in the Presence of Imidazolium Ionic Liquids. Int. J. Mol. Sci. 2018, 19, 887. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030887
Mehmood N, Alayoubi R, Husson E, Jacquard C, Büchs J, Sarazin C, Gosselin I. Kluyveromyces marxianus, an Attractive Yeast for Ethanolic Fermentation in the Presence of Imidazolium Ionic Liquids. International Journal of Molecular Sciences. 2018; 19(3):887. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030887
Chicago/Turabian StyleMehmood, Nasir, Ranim Alayoubi, Eric Husson, Cédric Jacquard, Jochen Büchs, Catherine Sarazin, and Isabelle Gosselin. 2018. "Kluyveromyces marxianus, an Attractive Yeast for Ethanolic Fermentation in the Presence of Imidazolium Ionic Liquids" International Journal of Molecular Sciences 19, no. 3: 887. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030887