Furosine Induced Apoptosis by the Regulation of STAT1/STAT2 and UBA7/UBE2L6 Genes in HepG2 Cells

Abstract
:1. Introduction
2. Results
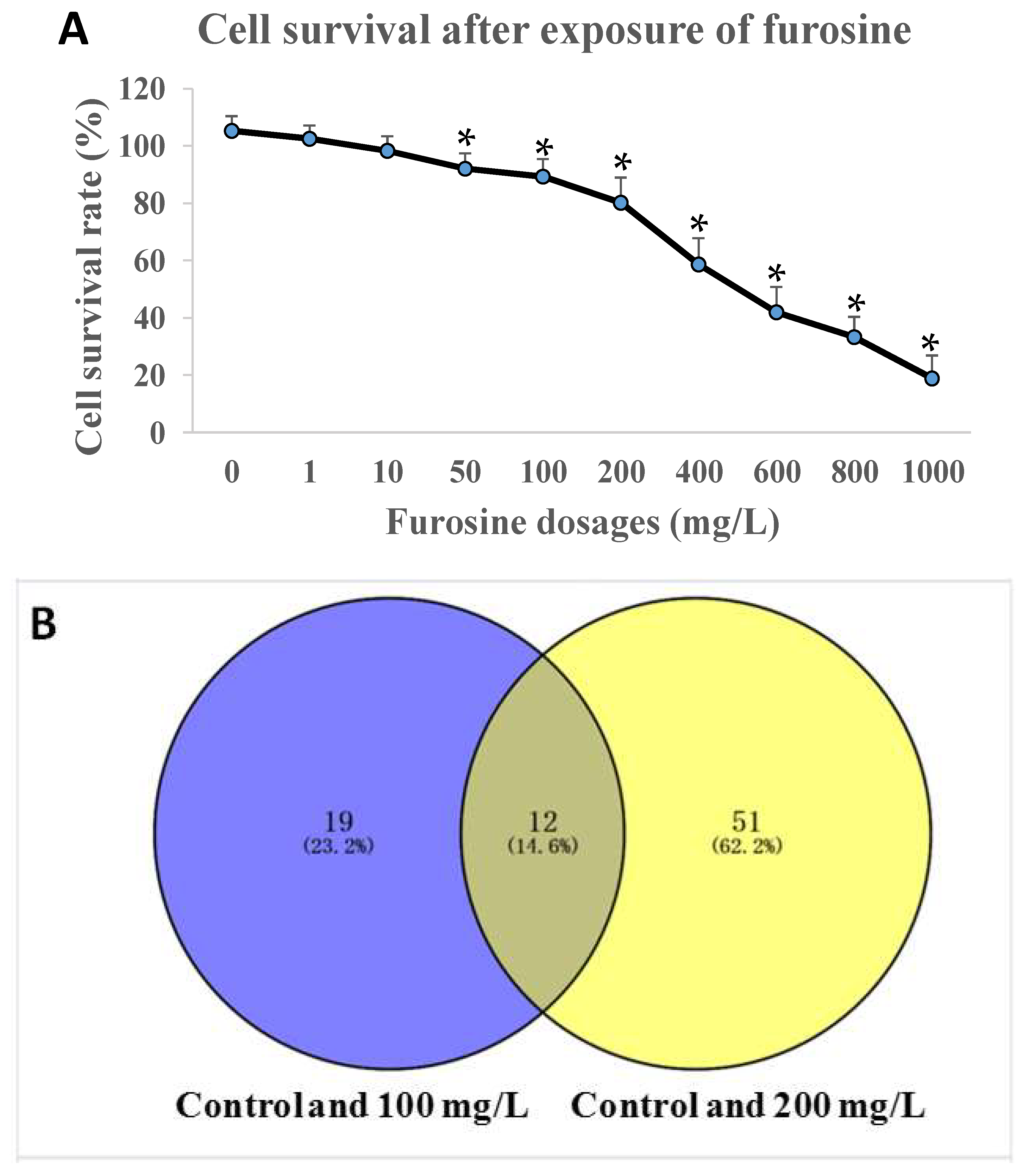
2.1. Furosine Inhibited HepG2 Cell Viability
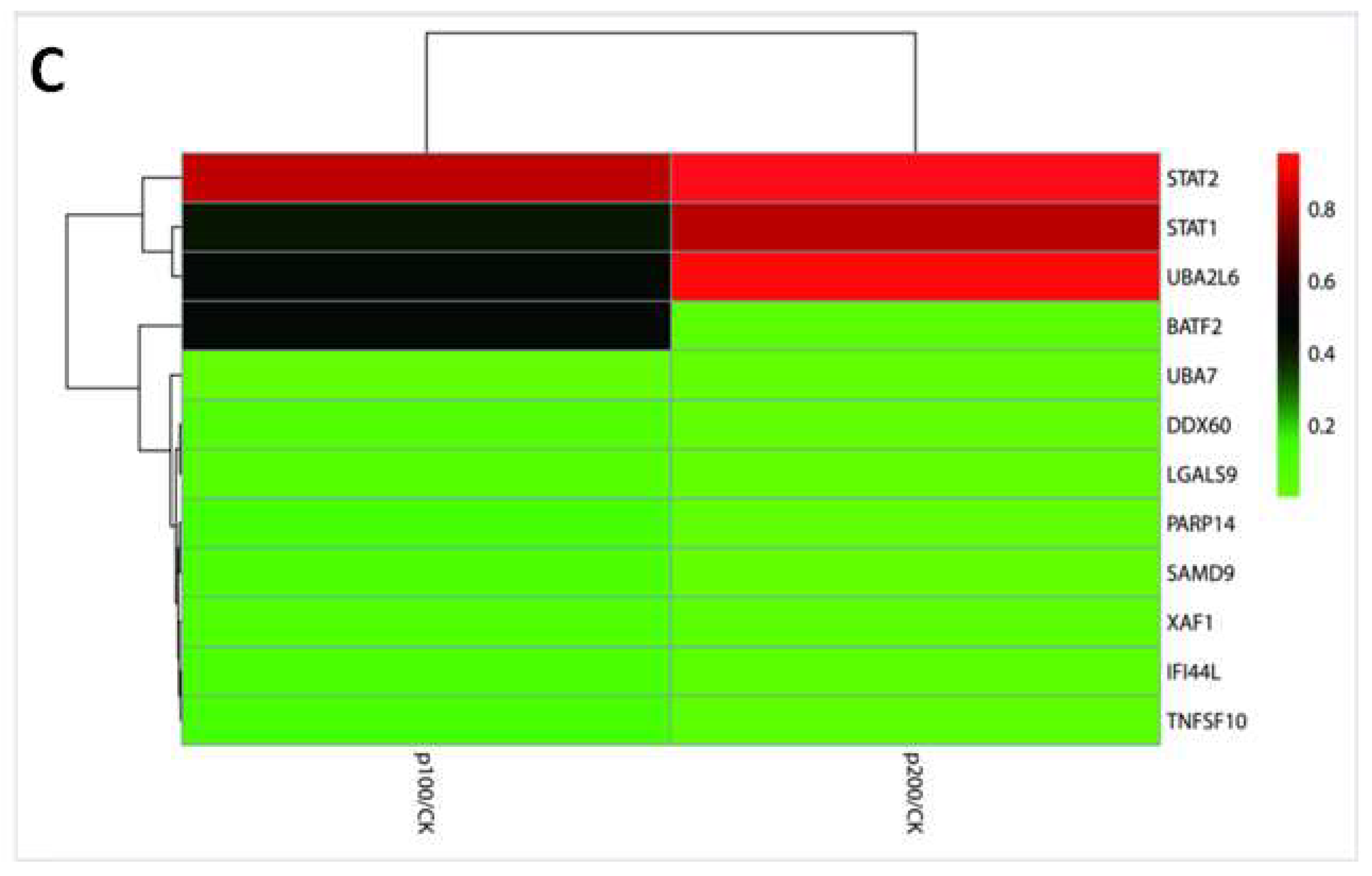
2.2. Transcriptomics Detection of HepG2 Cells
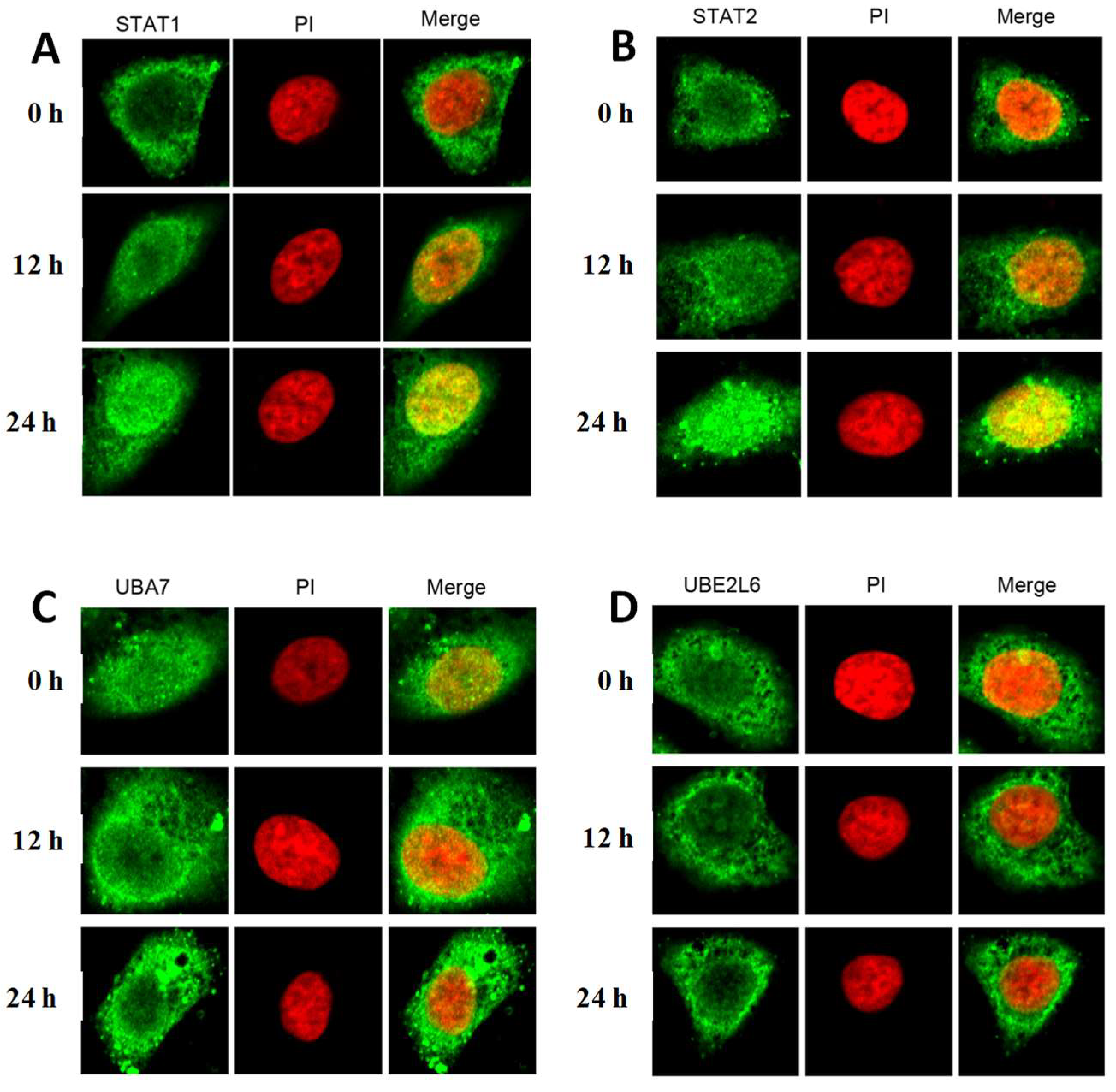
2.3. The Effect of Furosine on Cellular Location of STAT1, STAT2, UBA7, and UBE2L6
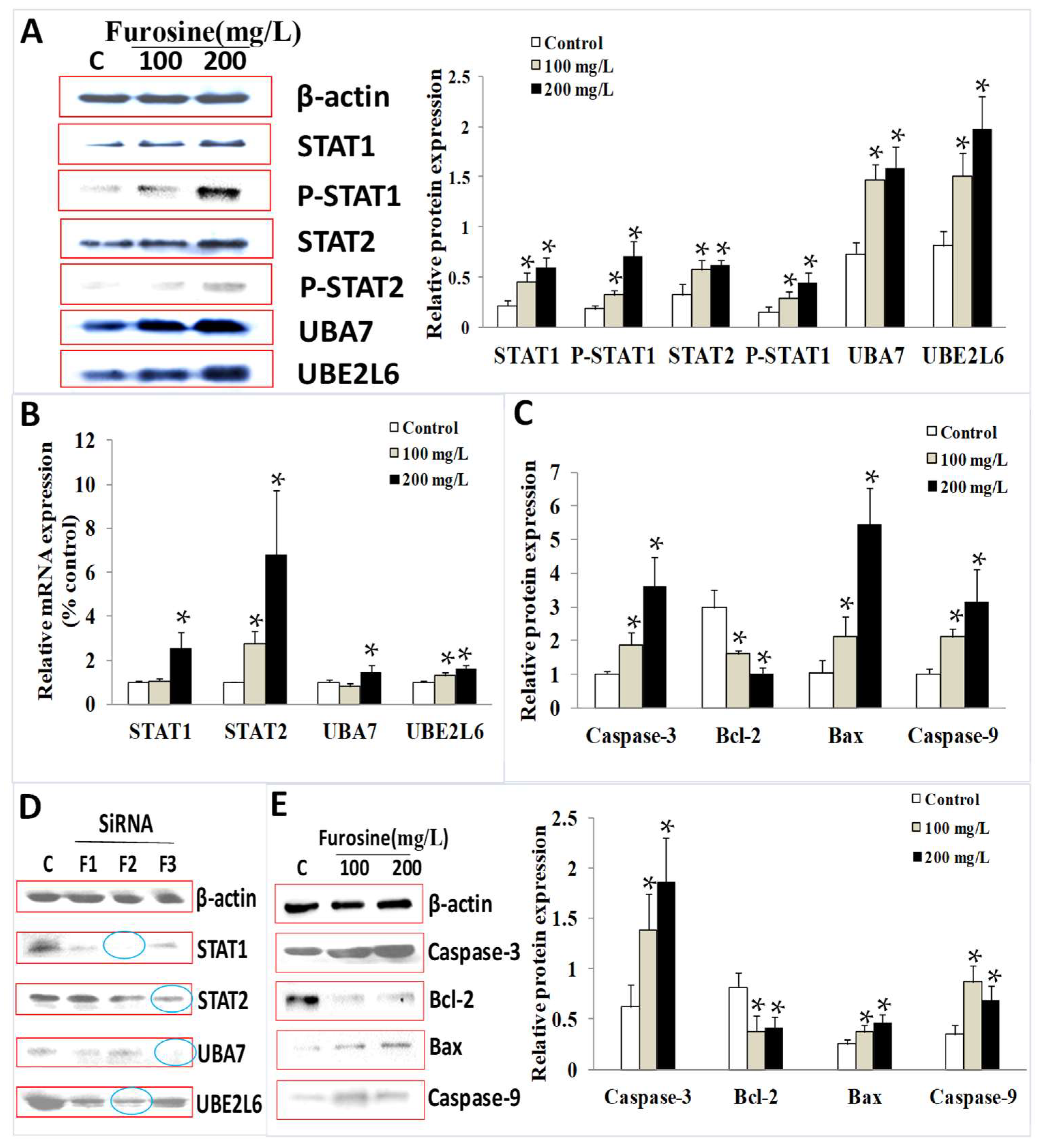
2.4. The Effect of Furosine on mRNA and Protein Expression of STAT1, STAT2, UBA7, and UBE2L6
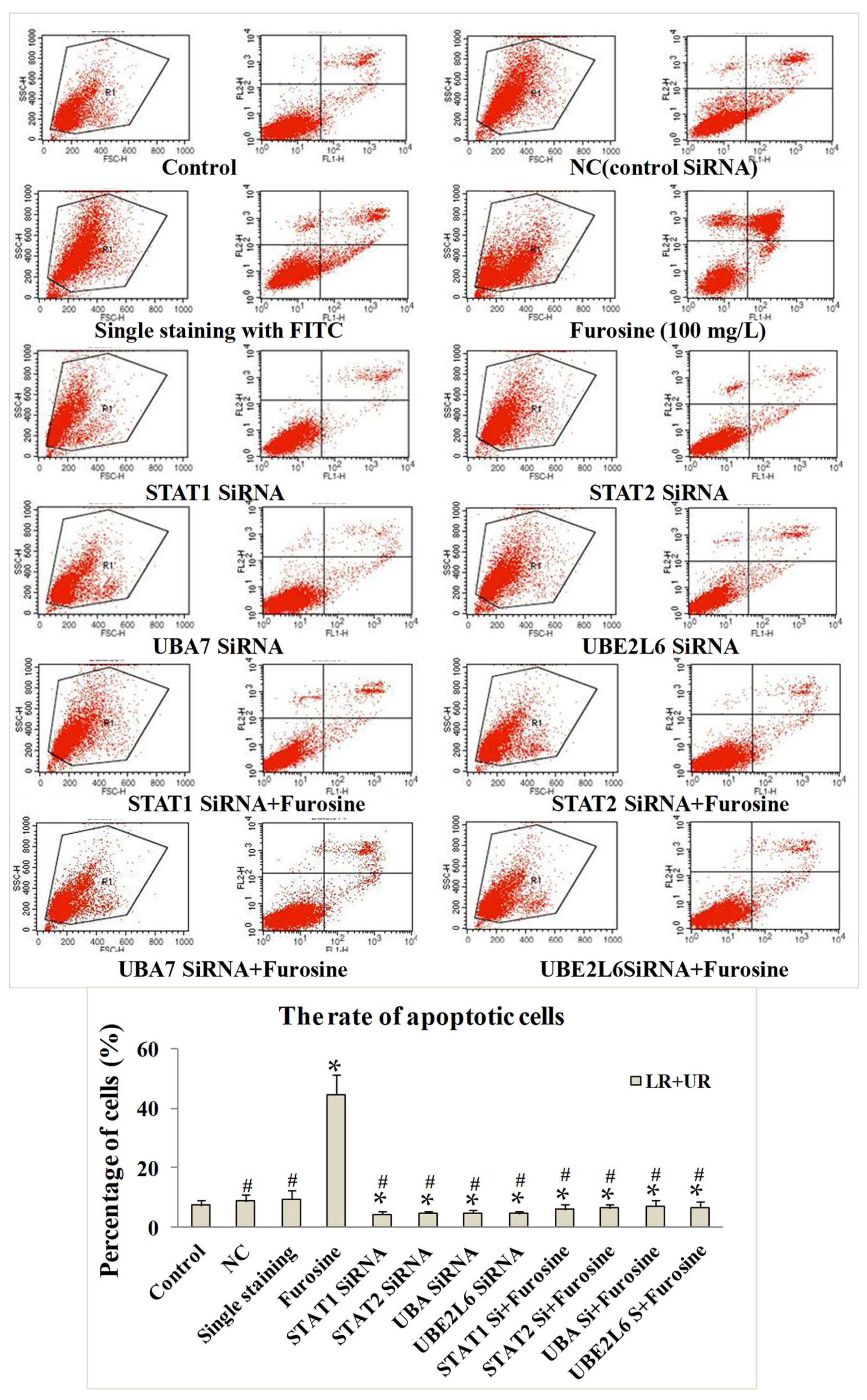
2.5. The Role of Furosine in Inducing Cell Apoptosis
2.6. The Role of STAT1, STAT2, UBA, and UBE2L6 in Participating in Apoptosis Induced by Furosine
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture and Viability Detection
4.3. Transcriptomics Detection and Data Analysis
4.4. Cell SiRNA Treatment
4.5. Quantitative Real-Time PCR (q-PCR) Analysis
4.6. Western Blot Analysis
4.7. Immunofluorescence Cell Staining
4.8. Cell Apoptosis Detection
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HepG2 | liver hepatocellular cells |
| STAT1 | signal transducer and activator of transcription 1 |
| STAT2 | signal transducer and activator of transcription 2 |
| UBA7 | ubiquitin-like modifier activating enzyme 7 |
| UBE2L6 | ubiquitin-conjugating enzyme E2L6 |
References
- Henle, T. Protein-bound advanced glycation endproducts (AGEs) as bioactive amino acid derivatives in foods. Amino Acids 2005, 29, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Ledl, F.; Schleicher, E. New aspects of the Maillard reaction in foods and in the human body. Angew. Chem. Int. Ed. Engl. 1990, 29, 565–594. [Google Scholar] [CrossRef]
- Radamendoza, M.; Olano, A.; Villamiel, M. Furosine as indicator of Maillard reaction in jams and fruit-based infant foods. J. Agric. Food Chem. 2002, 50, 4141–4145. [Google Scholar] [CrossRef]
- Rajchl, A.; Cizkova, H.; Voldrich, M.; Jiruskova, M.; Sevcik, R. Evaluation of shelf life and heat treatment of tomato products. Czech. J. Food Sci. 2009, 27, 130–133. [Google Scholar] [CrossRef]
- Villamiel, M.; Castillo, M.D.; Corzo, N.; Olano, A. Presence of furosine in honeys. J. Sci. Food Agric. 2001, 81, 790–793. [Google Scholar] [CrossRef]
- Seiquer, I.; Diaz-Alguacil, J.; Delgado-Andrade, C.; Lopez-Frias, M.; Munoz Hoyos, A.; Galdo, G.; Navarro, M.P. Diets rich in Maillard reaction products affect protein digestibility in adolescent males aged 11–14. Am. J. Clin. Nutr. 2006, 83, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Troise, A.D.; Fiore, A.; Wiltafsky, M.; Fogliano, V. Quantification of N-epsilon (2-Furoylmethyl)-l-lysine (furosine), Nepsilon-(Carboxymethyl)-l-lysine (CML), N-epsilon-(Carboxyethyl)-l-lysine (CEL) and total lysine through stable isotope dilution assay and tandem mass spectrometry. Food Chem. 2015, 188, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Xing, L.; Wang, J.Q.; Zheng, N. Toxicology studies of furosine in vitro/in vivo and exploration of the related mechanism. Toxicol. Lett. 2018, 291, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Baris, S.; Alroqi, F.; Kiykim, A.; Karakoc-Aydiner, E.; Ogulur, I.; Ozen, A.; Charbonnier, L.M.; Bakır, M.; Boztug, K.; Chatila, T.A.; et al. Severe Early-Onset Combined Immunodeficiency due to Heterozygous Gain-of-Function Mutations in STAT1. J. Clin. Immunol. 2016, 36, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Kaur, T.; Mukherjea, D.; Sheehan, K.; Jajoo, S.; Rybak, L.P.; Ramkumar, V. Short interfering RNA against STAT1 attenuates cisplatin-induced ototoxicity in the rat by suppressing inflammation. Cell Death Dis. 2011, 2, e180. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.H.; Park, O.; Henderson, L.E.; Abdelmegeed, M.A.; Moon, K.H.; Song, B.J. Lack of PPARα exacerbates lipopolysaccharide-induced liver toxicity through STAT1 inflammatory signaling and increased oxidative/nitrosative stress. Toxicol. Lett. 2011, 202, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Feng, D.; Wang, Y.; Lin, S.; Xu, L. Sodium tanshinone IIA sulfonate protects mice from ConA-induced hepatitis via inhibiting NF-κB and IFN-γ/STAT1 pathways. J. Clin. Immunol. 2008, 28, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Moore, F.; Naamane, N.; Colli, M.L.; Bouckenooghe, T.; Ortis, F.; Gurzov, E.N.; Igoillo-Esteve, M.; Mathieu, C.; Bontempi, G.; Thykjaer, T.; et al. STAT1 is a master regulator of pancreatic β-cell apoptosis and islet inflammation. J. Biol. Chem. 2011, 286, 929–941. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Waguespack, M.; Barker, S.A.; Li, S. Doxorubicin directs the accumulation of interleukin-12 induced IFN γ into tumors for enhancing STAT1 dependent antitumor effect. Clin. Cancer Res. 2007, 13, 4252–4260. [Google Scholar] [CrossRef] [PubMed]
- Alazawi, W.; Heath, H.; Petts, G.; Kudu, H.; Feakins, R.; O’Brien, A.; Goldin, R.; Foster, G.R. PTH-102 Stat2 is a key inflammatory molecule in human non-alcoholic fatty liver disease and murine liver injury. Gut 2015, 64, A452–A453. [Google Scholar] [CrossRef]
- Lee, J.W.; Kang, H.S.; Lee, J.Y.; Lee, E.J.; Rhim, H.; Yoon, J.H.; Seo, S.R.; Chung, C.K. The transcription factor STAT2 enhances proteasomal degradation of RCAN1 through the ubiquitin E3 ligase FBW7. Biochem. Biophys. Res. Commun. 2012, 420, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, M.K.; Chaudhuri, G. Down-regulation of UCRP and UBE2L6 in BRCA2 knocked-down human breast cells. Biochem. Biophys. Res. Commun. 2005, 328, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Hoyo-Becerra, C.; Liu, Z.; Yao, J.; Kaltwasser, B.; Gerken, G.; Hermann, D.M.; Schlaak, J.F. Rapid regulation of depression-associated genes in a new mouse model mimicking interferon-α-related depression in hepatitis C virus infection. Mol. Neurobiol. 2014, 52, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Borsini, A.; Cattaneo, A.; Malpighi, C.; Thuret, S.; Harrison, N.A.; Zunszain, P.A.; Pariante, C.M. Interferon-alpha reduces human hippocampal neurogenesis and increases apoptosis via activation of distinct STAT1-dependent mechanisms. Int. J. Neuropsychopharmacol. 2017, 21, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Qiao, L.; Wang, X.C.; Ding, J.; Chen, J. UHRF1 epigenetically down-regulates UbcH8 to inhibit apoptosis in cervical cancer cells. Cell Cycle 2018, 31, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wei, J.; Chen, F.; Xiao, X.; Huang, T. Epigenetic downregulation of the ISG15–conjugating enzyme UbcH8 impairs lipolysis and correlates with poor prognosis in nasopharyngeal carcinoma. Oncotarget 2015, 6, 41077–41091. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.J.; Chen, F.Z.; Chen, C.H.; Wan, X.X.; Zhou, X.; Fang, Q.; Zhang, D.Z. ISG15 inhibits cancer cell growth and promotes apoptosis. Int. J. Mol. Med. 2017, 39, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Salla, R.; Tuija, H.K.; Anne, P.; Sarhadi, V.K.; Jaakko, H.; Sakari, K.; Juha, S.; Harriet, W.; Sisko, A. Pathways affected by asbestos exposure in normal and tumour tissue of lung cancer patients. BMC Med. Genom. 2008, 1, 55. [Google Scholar]
- Klimmeck, D.; Hansson, J.; Raffel, S.; Vakhrushe, S.Y.; Trumpp, A.; Krijgsveld, J. Proteomic cornerstones of hematopoietic stem cell differentiation: Distinct signatures of multipotent progenitors and myeloid committed cells. Mol. Cell. Proteom. 2012, 11, 286–302. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.D.; Wu, K.H.; Lu, S.; Xiong, J.L.; Tan, L.J. UBE2L6 expression is highly correlated with an osteoporotic candidate gene STAT-1. Osteoporos. Int. 2013, 24, 566–567. [Google Scholar]
- Mittal, M.K.; Chaudhuri, G. Requirement of functionally active BRCA2 protein for the expression of IRF9-regulated genes in human breast cells. Cancer Res. 2009, 69, A16. [Google Scholar] [CrossRef]
- Przanowski, P.; Loska, S.; Cysewski, D.; Dabrowski, M.; Kaminska, B. ISG’ylation increases stability of numerous proteins including Stat1, which prevents premature termination of immune response in LPS-stimulated microglia. Neurochem. Int. 2018, 112, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Stefan, M.; Wei, C.; Lombardi, A.; Li, C.W.; Concepcion, E.S. Genetic epigenetic dysregulation of thymic TSH receptor gene expression triggers thyroid autoimmunity. Proc. Natl. Acad. Sci. USA 2014, 111, 12562–12567. [Google Scholar] [CrossRef] [PubMed]
- Sobh, A.; Chou, J.; Schneider, L.; Geha, R.S.; Massaad, M.J. Chronic mucocutaneous candidiasis associated with an SH2 domain gain-of-function mutation that enhances STAT1 phosphorylation. Allergy Clin. Immun. 2016, 138, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Sun, M.; Yuan, X.; Ji, L.; Jin, Y. Enterovirus 71 suppresses interferon responses by blocking Janus kinase (JAK)/signal transducer and activator of transcription (STAT) signaling through inducing karyopherin-α1 degradation. J. Biol. Chem. 2017, 292, 10262–10274. [Google Scholar] [CrossRef] [PubMed]
- Sdnk, B.; Umasuthan, N.; Priyathilaka, T.T.; Thulasitha, W.S.; Jdhe, J. Molecular cloning, transcriptional profiling, and subcellular localization of signal transducer and activator of transcription 2 (STAT2) ortholog from rock bream, Oplegnathus fasciatus. Gene 2017, 626, 95–105. [Google Scholar]
- Cohen, P.; Tcherpakov, M. Will the ubiquitin system furnish as many drug targets as protein kinases. Cell 2010, 143, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Salajegheh, M.; Kong, S.W.; Pinkus, J.L. Interferon-stimulated gene 15 (ISG15) cnjugates proteins in dermatomyositis muscle with perifascicular atrophy. Ann. Neurol. 2010, 67, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Moudry, P.; Lukas, C.; Macurek, L.; Hanzlikova, H.; Hodny, Z.; Lukas, J.; Bartek, J. Ubiquitin. Cell Cycle 2012, 11, 1573–1582. [Google Scholar] [CrossRef] [PubMed]
- Mitupatum, T.; Aree, K.; Kittisenachai, K.S.; Roytrakul, S.; Puthong, S.; Kangsadalampai, S.; Rojpibulstit, P. mRNA Expression of Bax, Bcl-2, p53, Cathepsin B, Caspase-3 and Caspase-9 in the HepG2 cell line following induction by a novel monoclonal Ab Hep88 mAb: Cross-Talk for paraptosis and apoptosis. Asian Pac. J. Cancer Prev. 2016, 2, 703–712. [Google Scholar] [CrossRef]
- Wu, R.; Tang, S.; Wang, M.; Xu, X.; Yao, C.; Wang, S.M. MicroRNA-497 induces apoptosis and suppresses proliferation via the Bcl-2/Bax-Caspase9-Caspase3 pathway and cyclin D2 protein in HUVECs. PLoS ONE 2016, 11, e0167052. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Sun, K.; Wang, Y.; Dong, S.; Wang, C.; Liu, L.; Wu, Y. Role of Cyt-C/caspases-9,3, Bax/Bcl-2 and the FAS death receptor pathway in apoptosis induced by zinc oxide nanoparticles in human aortic endothelial cells and the protective effect by alpha-lipoic acid. Chem. Biol. Interact. 2016, 258, 40–51. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequences (5′ → 3′) | |
|---|---|---|
| Forward Primer | Reverse Primer | |
| STAT1 | GCTGCTGCTCCACAAGATGTT | TCTGCTGCCTTCGCTTCCA |
| STAT2 | TGGCAGTGACAGAGGAGTTACA | CAGGCAATGGAGAGTTGGTTCA |
| UBA7 | TCCAGAAGATGAGACGCTCCTT | AGCAACAGTCACACCTCCATTC |
| UBE2L6 | GGAAGCCTTACACCAAGCCTTA | GAGTCAGGAGGTCAGCAAGTTC |
| Caspase-3 | TGGAGGCTGACTTCCTGTATGC | TTCCGTTGCCACCTTCCTGTTA |
| Bax | CCAGGATGCGTCCACCAAGA | GAAGTCCAGTGTCCAGCCCAT |
| Bcl-2 | GCATCTTCTCCTTCCAGCCTGA | TCTGCGAAGTCACGACGGTAG |
| Caspase-9 | TGAATGACCACCTAGAGCCTTG | AGAACCACACCAGCCACAGT |
| GAPDH | CGTCCCGTAGACAAAATGGT | TTGATGGCAACAATCTCCAC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Xing, L.; Zhao, N.; Wang, J.; Zheng, N. Furosine Induced Apoptosis by the Regulation of STAT1/STAT2 and UBA7/UBE2L6 Genes in HepG2 Cells. Int. J. Mol. Sci. 2018, 19, 1629. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061629
Li H, Xing L, Zhao N, Wang J, Zheng N. Furosine Induced Apoptosis by the Regulation of STAT1/STAT2 and UBA7/UBE2L6 Genes in HepG2 Cells. International Journal of Molecular Sciences. 2018; 19(6):1629. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061629
Chicago/Turabian StyleLi, Huiying, Lei Xing, Nan Zhao, Jiaqi Wang, and Nan Zheng. 2018. "Furosine Induced Apoptosis by the Regulation of STAT1/STAT2 and UBA7/UBE2L6 Genes in HepG2 Cells" International Journal of Molecular Sciences 19, no. 6: 1629. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061629