Methyl 3,4-Dihydroxybenzoate Enhances Resistance to Oxidative Stressors and Lifespan in C. elegans Partially via daf-2/daf-16

Abstract
:1. Introduction
2. Results
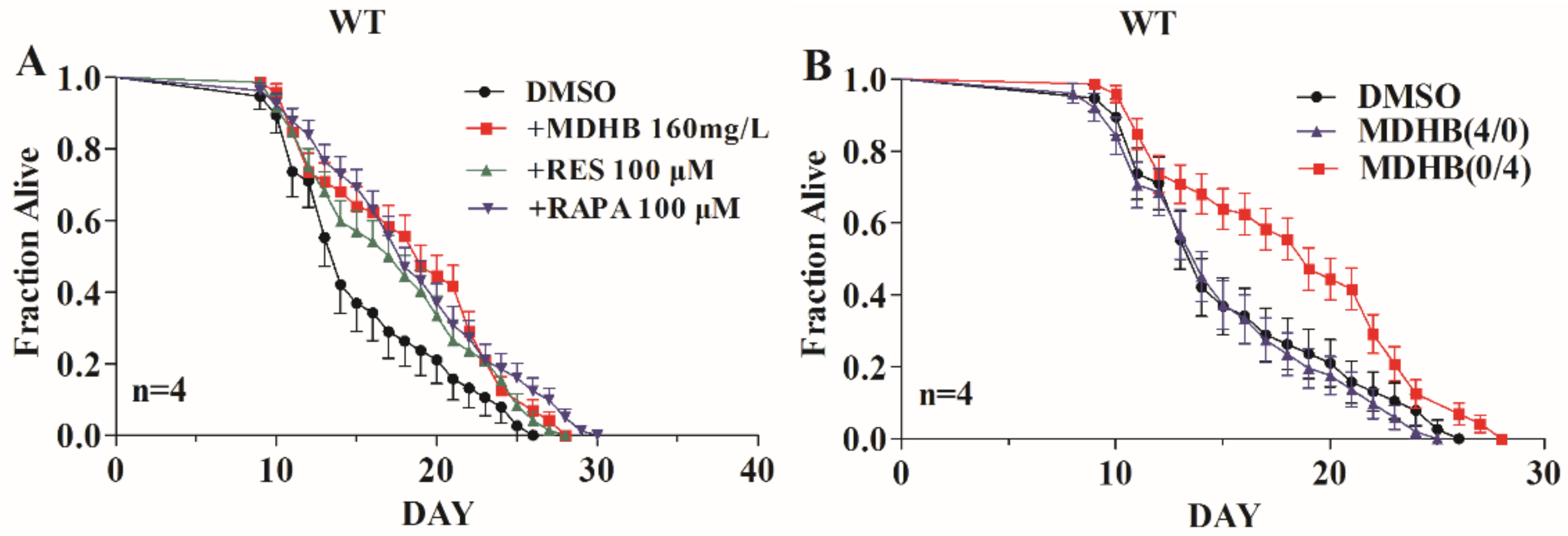
2.1. Exposure to MDHB during Adulthood Extends the Mean Lifespan of Wild-Type C. elegans
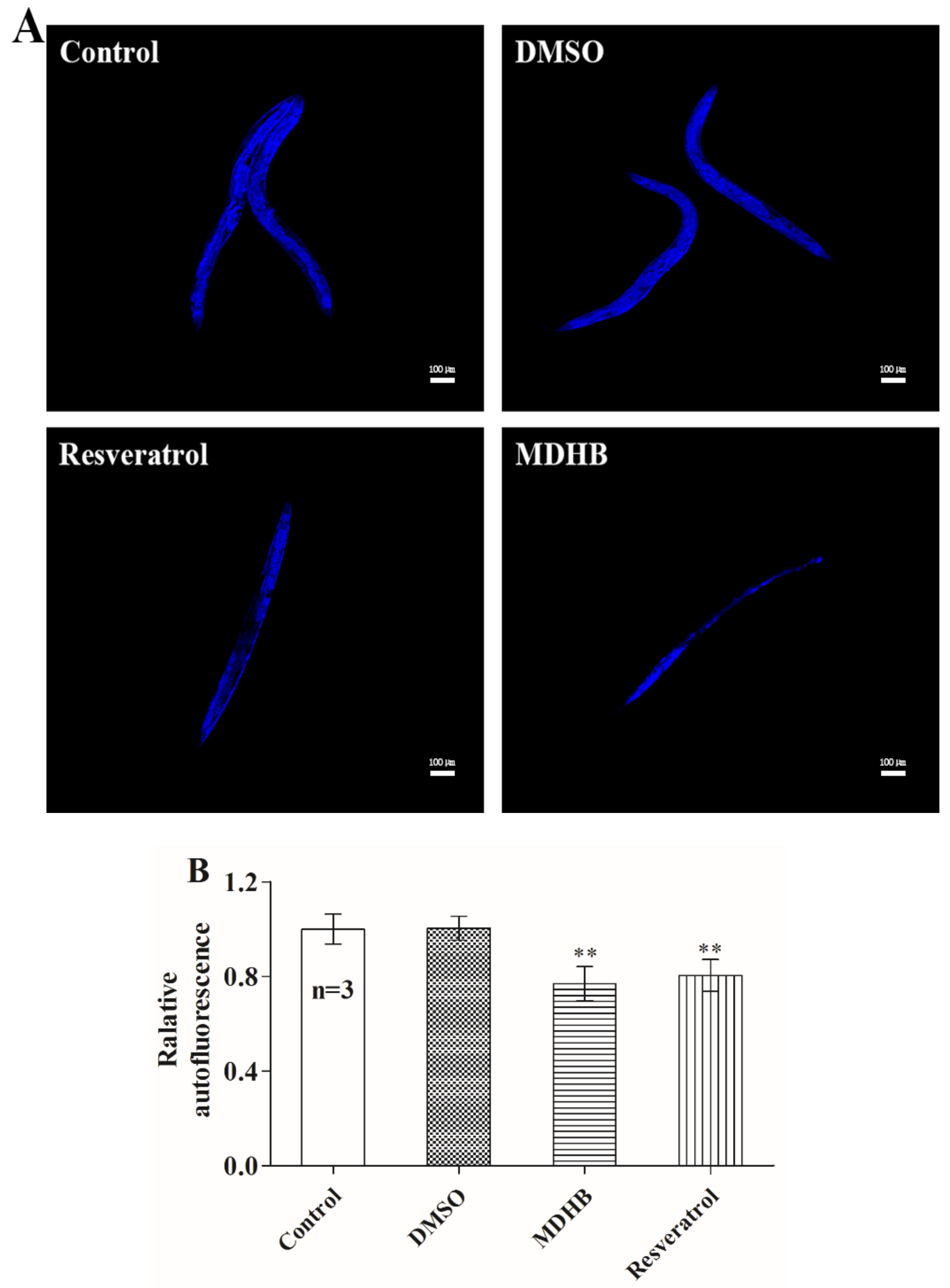
2.2. MDHB Reduces Lipofuscin Accumulation
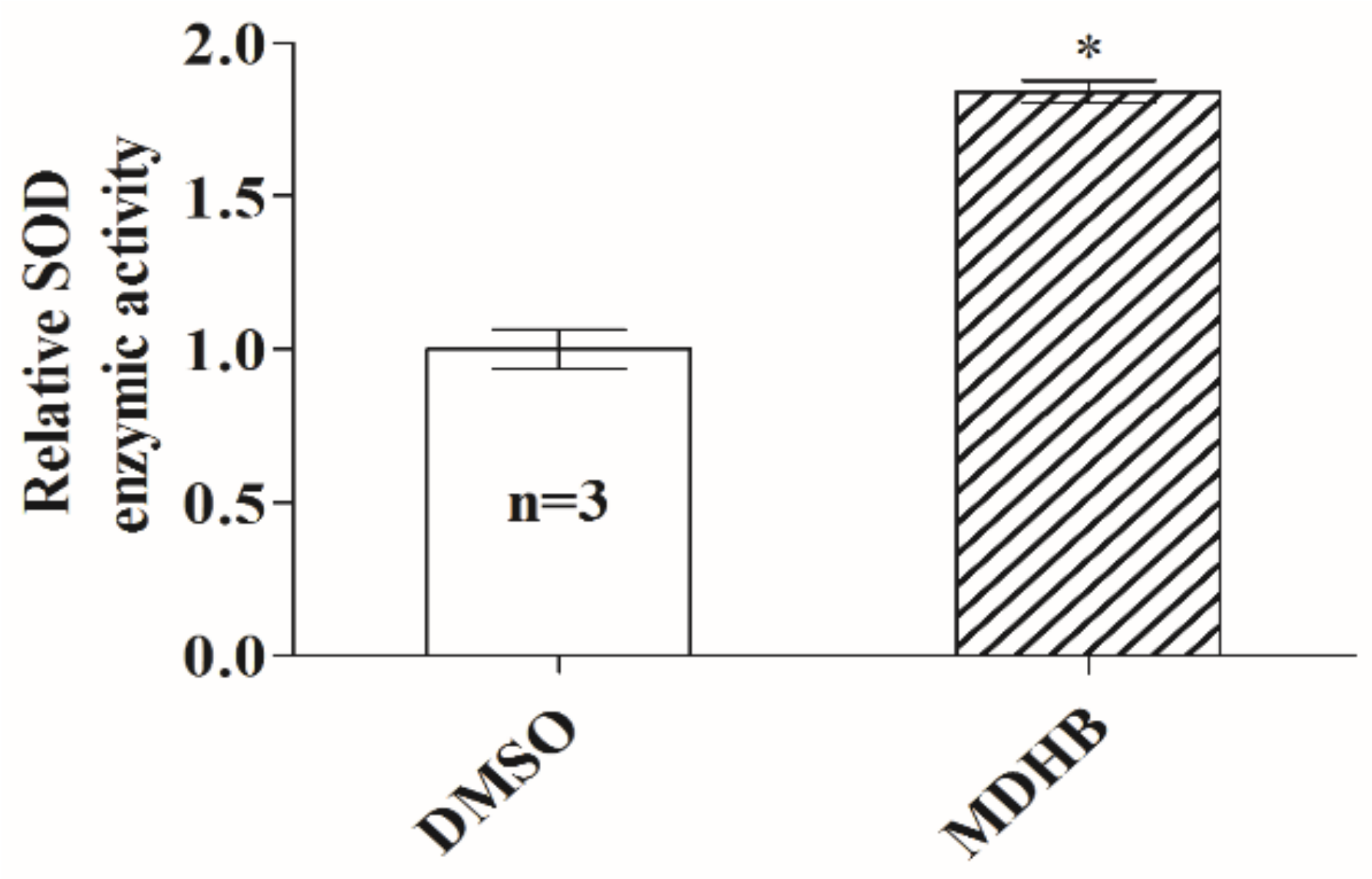
2.3. Effects of MDHB on SOD Activity in C. elegans
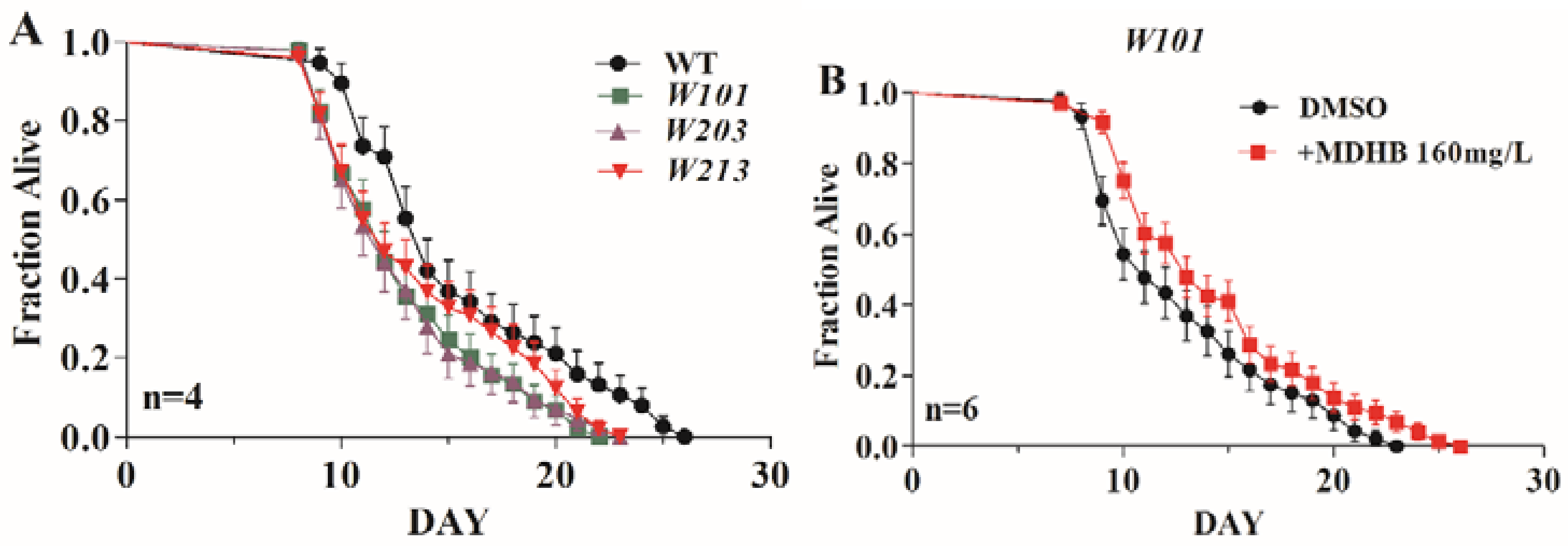
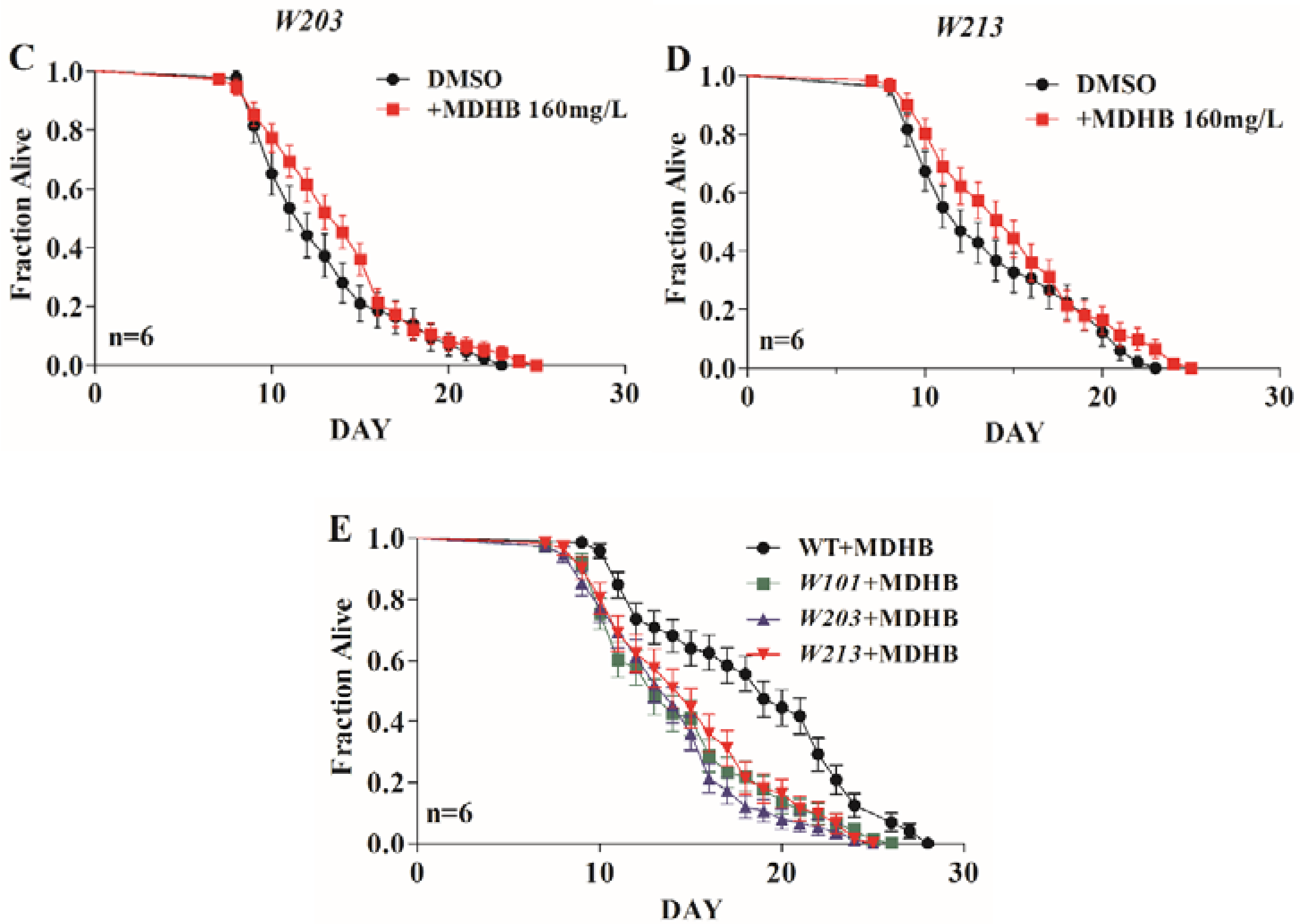
2.4. MDHB Extends the Lifespan of W06A7.4 Loss-of-Function Mutants
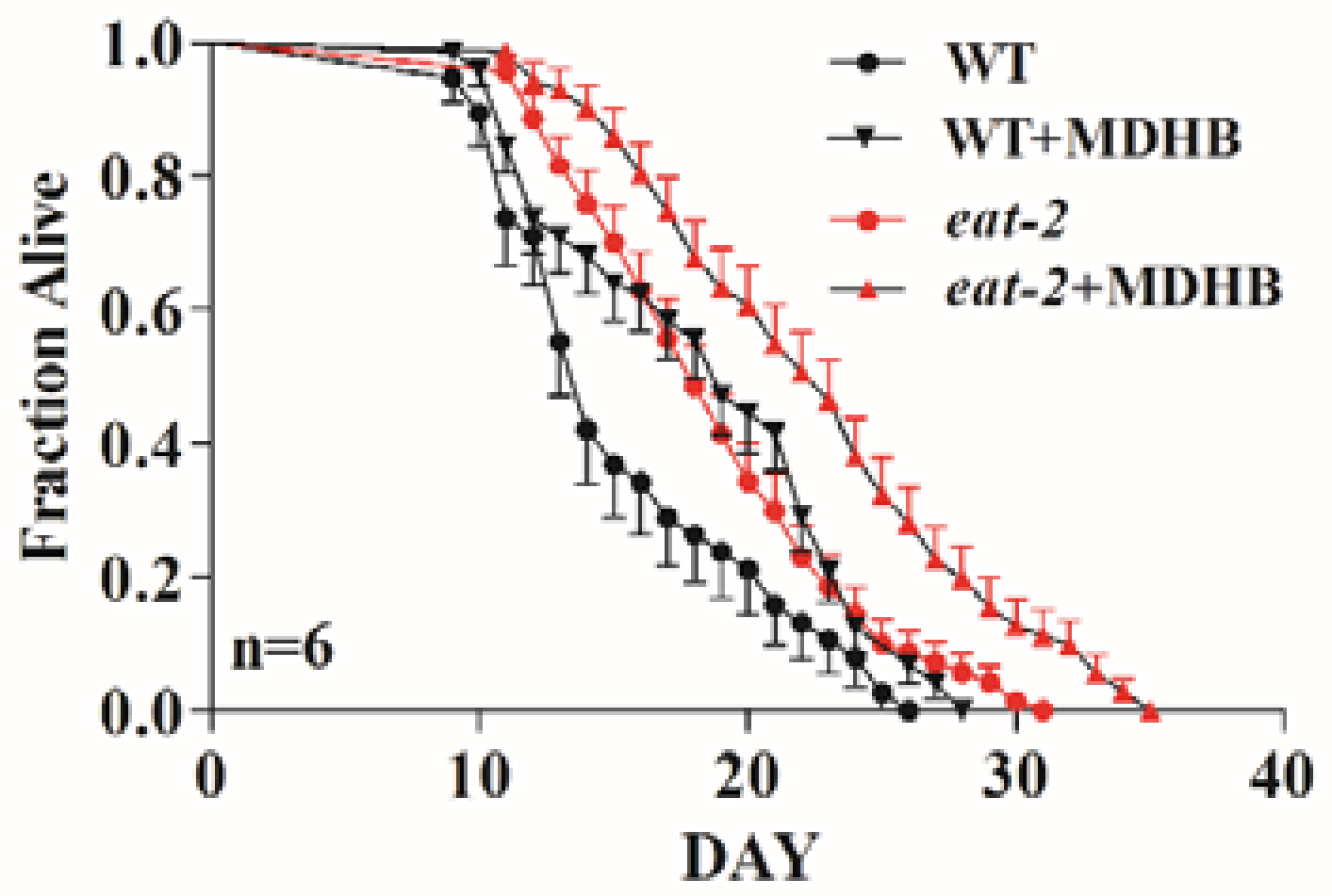
2.5. MDHB Does Not Appear to Lengthen Lifespan via Caloric Restriction
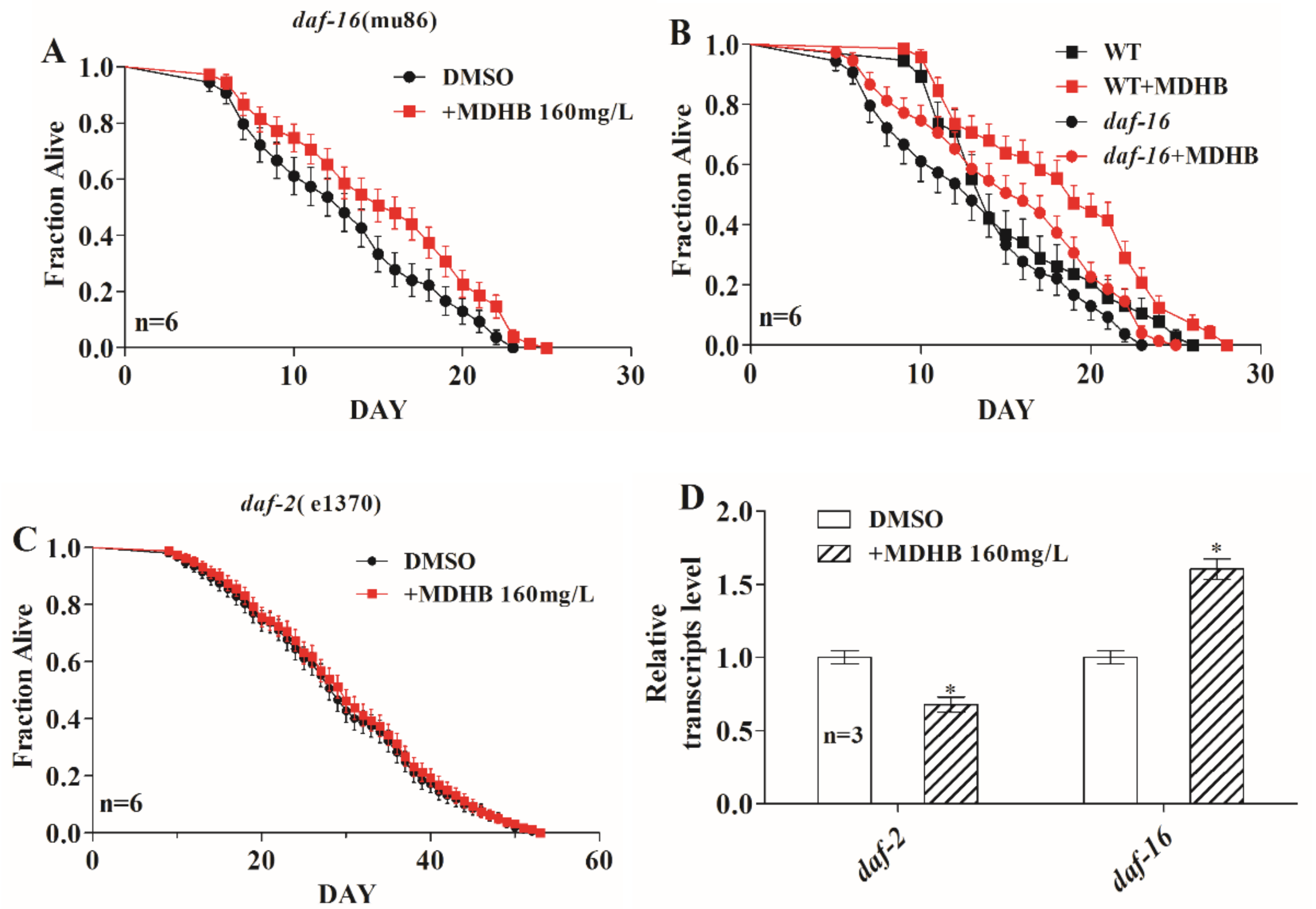
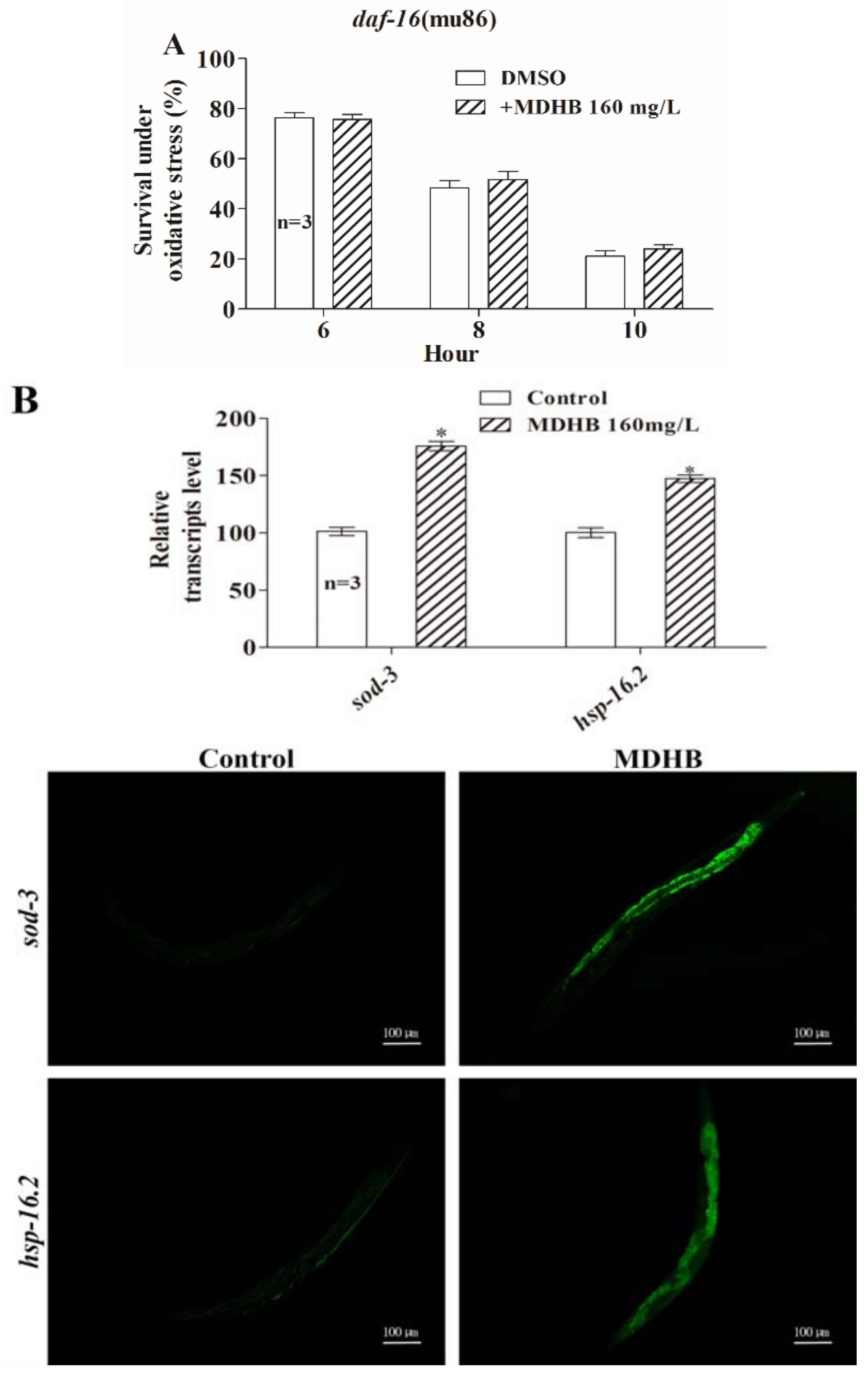
2.6. MDHB Prolongs the Lifespan of C. elegans Partially through the daf-2/daf-16 Signaling Pathway
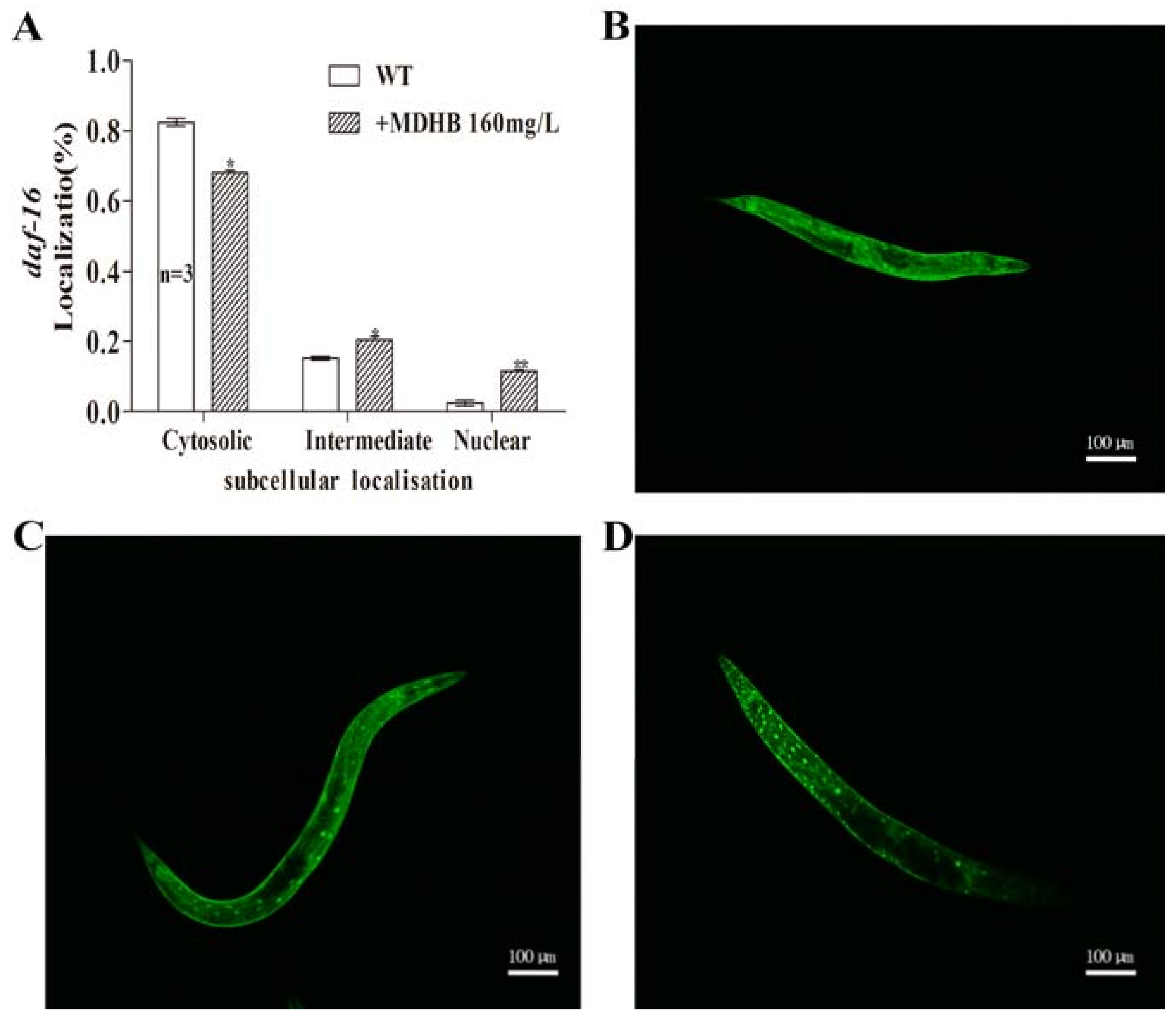
2.7. MDHB Promotes daf-16/FoxO Nuclear Localization
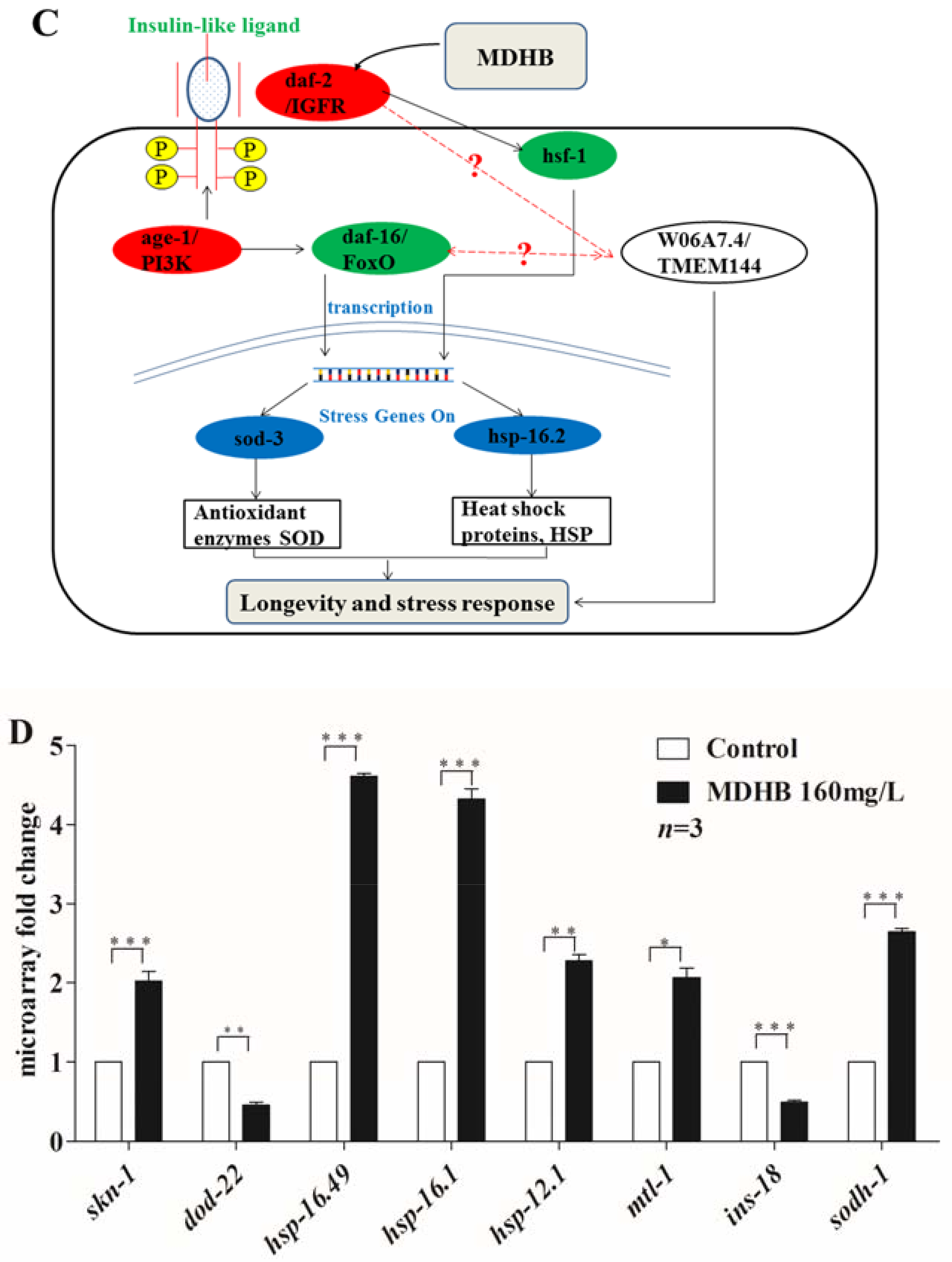
2.8. Effects of MDHB on the Expression of daf-2/daf-16 Target Genes
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. General Methods and Strains
4.3. W06A7.4 Gene Knockout Through a CRISPR-Cas9 System
4.4. Lifespan Assay
4.5. Lipofuscin Accumulation Assay
4.6. Measurement of SOD Activity
4.7. Oxidative Stress Resistance Assay
4.8. Expression of sod-3 and hsp-16.2
4.9. daf-16 Localization
4.10. Microarray Profiling Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MDHB | Methyl 3,4-dihydroxybenzoate |
| C. elegans | Caenorhabditis elegans |
| IIS | Insulin/Insulin-like growth factor-1 signaling pathway |
| daf-16 | Orthology of the FoxO in C. elegans |
| FoxO | Forxhead box protein O |
| CR | Calorie restriction |
| DMSO | Dimethyl sulfoxide |
| ROS | Reactive oxygen species |
| SOD | Superoxide Dismutase |
| RES | Resveratrol |
| RAPA | Rapamycin |
| 5-FuDR | 5-fluorouracil deoxyribotside |
| E. coli | Escherichia coli |
| NGM | Nematode growth medium |
| LB | Luria-Bertani |
| RIN | RNA integrity |
References
- Honda, Y.; Honda, S. Life span extensions associated with upregulation of gene expression of antioxidant enzymes in Caenorhabditis elegans; studies of mutation in the age-1, PI3 kinase homologue and exposure to hyperoxia. J. Am. Aging Assoc. 2002, 25, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Boccardi, V.; Mecocci, P. Telomerase activation and human health-span: An open issue. Aging Clin. Exp. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Evason, K.; Huang, C.; Yamben, I.; Covey, D.F.; Kornfeld, K. Anticonvulsant Medications Extend Worm Life-Span. Science 2005, 307, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Regitz, C.; Fitzenberger, E.; Mahn, F.L.; Dußling, L.M.; Wenzel, U. Resveratrol reduces amyloid-β (Aβ1–42)-induced paralysis through targeting proteostasis in an Alzheimer model of Caenorhabditiselegans. Eur. J. Nutr. 2016, 55, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The Genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [PubMed]
- Sulston, J.E.; Brenner, S. The DNA of Caenorhabditis elegans. Genetics 1974, 77, 95–104. [Google Scholar] [PubMed]
- Johnson, T.E.; Wood, W.B. Genetic analysis of life-span in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 1982, 97, 6603–6607. [Google Scholar] [CrossRef]
- Tissenbaum, H.A. Using C. elegans for aging research. Invertebr. Reprod. Dev. 2015, 59 (Suppl. 1), 59–63. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Wang, L.F.; Pan, J.P.; Mi, X.N.; Zhang, Z.; Geng, H.J.; Wang, J.H.; Hu, S.H.; Zhang, W.; Gao, Q.; et al. Neuroprotective effects of methyl 3,4-Dihydroxybenzoate against TBHP-induced oxidative damage in SH-SY5Y cells. Molecules 2016, 21, 1071. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Su, C.F.; Zhang, Z.; Wang, C.Y.; Luo, J.Q.; Zhou, X.W.; Cai, L.; Yan, L.; Zhang, W.; Luo, H.M. Neuroprotective Effects of Methyl 3,4-dihydroxybenzoate Against H2O2-Induced Apoptosis in RGC-5 Cells. J. Pharmacol. Sci. 2014, 125, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhou, X.; Zhou, X.; Xu, X.; Liao, M.; Yan, L.; Lv, R.; Luo, H. Methyl 3,4-dihydroxybenzoate promotes neurite outgrowth of cortical neurons cultured in vitro. Neural Regen. Res. 2012, 7, 971–977. [Google Scholar] [PubMed]
- Zhang, Z.; Cai, L.; Zhou, X.; Su, C.; Xiao, F.; Gao, Q.; Luo, H. Methyl 3,4-dihydroxybenzoate promote rat cortical neurons survival and neurite outgrowth through the adenosine A2a receptor/PI3K/Akt signaling pathway. Neuroreport 2015, 26, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Cai, L.; Geng, H.J.; Su, C.F.; Yan, L.; Wang, J.H.; Gao, Q.; Luo, H.M. Methyl 3,4-dihydroxybenzoate extends the lifespan of Caenorhabditis elegans, partly via W06A7.4 gene. Exp. Gerontol. 2014, 60, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.T.; Hu, P.J. Insulin/insulin-like growth factor signaling in C. elegans. WormBook 2013, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Yen, K.; Mobbs, C.V. Evidence for only two independent pathways for decreasing senescence in Caenorhabditis elegans. AGE. 2010, 32, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Henderson, S.T.; Johnson, T.E. daf-16 integrates developmental and environmental inputs to mediate aging in the nematode Caenorhabditis elegans. Brief Commun. 2001, 11, 1975–1980. [Google Scholar] [CrossRef]
- Grala, T.M.; Kay, J.K.; Walker, C.G.; Sheahan, A.J.; Littlejohn, M.D.; Lucy, M.C.; Roche, J.R. Expression analysis of key somatotropic axis and liporegulatory genes in ghreline and obestatin-infused dairy cows. Domest. Anim. Endocrinol. 2010, 39, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C. The plasticity of aging: Insights from long-lived mutants. Cell 2005, 120, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Baumeister, R.; Schaffitzel, E.; Hertweck, M. Endocrine signaling in Caenorhabditis elegans controls stress response and longevity. J. Endocrinol. 2006, 190, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Lakowski, B.; Hekimi, S. The genetics of caloric restriction in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 1998, 95, 13091–13096. [Google Scholar] [CrossRef] [PubMed]
- Brunk, U.T.; Terman, A. Lipofuscin: Mechanisms of age-related accumulation and influence on cell function. Free Radic. Biol. Med. 2002, 33, 611–619. [Google Scholar] [CrossRef]
- Berdichevsky, A.; Nedelcu, S.; Boulias, K.; Bishop, N.A.; Guarente, L.; Horvitz, H.R. 3-Ketoacyl thiolase delays aging of Caenorhabditis elegans and is required for lifespan extension mediated by sir-2.1. Proc. Natl. Acad. Sci. USA 2010, 107, 18927–18932. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.D.; Wilson, M.A.; Zhu, M.; Wolkow, C.A.; de Cabo, R.; Ingram, D.K.; Zou, S. Dietary deprivation extends lifespan in Caenorhabditis elegans. Aging Cell 2006, 5, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Mair, W.; Dillin, A. Aging and survival: The genetics of life span extension by dietary restriction. Annu. Rev. Biochem. 2008, 77, 727–754. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, T.L.; Smith, E.D.; Tsuchiya, M.; Welton, K.L.; Thomas, J.H.; Fields, S.; Kennedy, B.K.; Kaeberlein, M. Lifespan extension in Caenorhabditis elegans by complete removal of food. Aging Cell 2006, 5, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Bishop, N.A.; Guarente, L. Two neurons mediate diet-restriction-induced longevity in C. elegans. Nature 2007, 447, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, R.; Rui, Q.; Wang, D. Intestinal Insulin Signaling Encodes Two Different Molecular Mechanisms for the Shortened Longevity Induced by Graphene Oxide in Caenorhabditis elegans. Sci. Rep. 2016, 6, 24024. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.S.; Narasimhan, S.D.; Yen, K.; Tissenbaum, H.A. A new DAF-16 isoform regulates longevity. Nature 2010, 466, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.L. Fat synthesis and adiposity regulation in Caenorhabditis elegans. Trends Endocrinol. Metab. 2009, 20, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.; Snoek, L.B.; De Bono, M.; Kammenga, J.E. Worms under stress: C. elegans stress response and its relevance to complex human disease and aging. Trends Genet. 2013, 29, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Li, J.; Wang, W.; Fu, Y. Gengnianchun, a Traditional Chinese Medicine, Enhances Oxidative Stress Resistance and Lifespan in Caenorhabditis elegans by Modulating daf-16/FoxO. Evid. Based Complement. Altern. Med. 2017. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, A.; Oh, S.W.; Tissenbaum, H.A. Worming pathways to and from DAF-16/FoxO. Exp. Gerontol. 2006, 41, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Tullet, J.M.A.; Green, J.W.; Au, C.; Benedetto, A.; Thompson, M.A.; Clark, E.; Gilliat, A.F.; Young, A.; Schmeisser, K.; Gems, D. The SKN-1/Nrf2 transcription factor can protect against oxidative stress and increase lifespan in C. elegans by distinct mechanisms. Aging Cell 2017, 16, 1191–1194. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.W.; Cheon, S.M.; Lee, M.Y.; Kim, H.J.; Jeon, H.; Cha, D.S. Catalpol Modulates Lifespan via DAF-16/FoxO and SKN-1/Nrf2 Activation in Caenorhabditis elegans. Evid. Based Complement. Altern. Med. 2015, 2015, 524878. [Google Scholar] [CrossRef] [PubMed]
- Senchuk, M.M.; Dues, D.J.; Schaar, C.E.; Johnson, B.K.; Madaj, Z.B.; Bowman, M.J.; Winn, M.E.; Van Raamsdonk, J.M. Activation of DAF-16/FoxO by reactive oxygen species contributes to longevity in long-lived mitochondrial mutants in Caenorhabditis elegans. PLoS Genet. 2018. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.T.; McCarroll, S.A.; Bargmann, C.I.; Fraser, A.; Kamath, R.S.; Ahringer, J.; Li, H.; Kenyon, C. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nat. Genet. 2003, 424, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Artyukhin, A.B.; Yim, J.J.; Cheong, M.; Avery, L. Starvation-induced collective behavior in C. elegans. Sci. Rep. 2015. [Google Scholar] [CrossRef] [PubMed]
- Gengyo-Ando, K.; Mitani, S. Characterization of mutations induced by ethyl methanesulfonate, UV, and trimethylpsoralen in the nematode Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 2000, 269, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Friedland, A.E.; Tzur, Y.B.; Esvelt, K.M.; Colaiácovo, M.P.; Church, G.M.; Calarco, J.A. Heritable genome editing in C. elegans via a CRISPR-Cas9 system. Nat. Methods 2013, 10, 741–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiernagle, T. Maintenance of C. elegans. WormBook 2006, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; Lea, K.; Zucker-Aprison, E.; Blumenthal, T. The spliceosomal snRNAs of Caenorhabditis elegans. Nucleic Acids Res. 1999, 18, 2633–2642. [Google Scholar] [CrossRef]
- Kadandale, P.; Chatterjee, I.; Singson, A. Germline transformation of Caenorhabditis elegans by injection. Methods Mol. Biol. 2009, 518, 123–133. [Google Scholar] [PubMed]
- Meyer, L.R.; Zweig, A.S.; Hinrichs, A.S.; Karolchik, D.; Kuhn, R.M.; Wong, M.; Sloan, C.A.; Rosenbloom, K.R.; Roe, G.; Rhead, B.; et al. The UCSC Genome Browser database: Extensions and updates 2013. Nucleic Acids Res. 2013, 41, D64–D69. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.A.; Fleming, J.T. Basic Culture Methods. Methods Cell Biol. 1995, 48, 3–29. [Google Scholar] [PubMed]
- Won, S.M.; Cha, H.U.; Yi, S.S.; Kim, S.J.; Park, S.K. Tenebrio molitor Extracts Modulate the Response to Environmental Stressors and Extend Lifespan in Caenorhabditis elegans. J. Med. Food 2016. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.B.; Xing, M.M.; Kim, D.K. Lifespan-extending and stress resistance properties of brazilin from Caesalpinia sappan in Caenorhabditis elegans. Arch. Pharm. Res. 2017, 40, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Possik, E.; Pause, A. Measuring oxidative stress resistance of Caenorhabditis elegans in 96-well microtiter plates. J. Vis. Exp. 2015, e52746. [Google Scholar] [CrossRef] [PubMed]
- Walker, G.A.; Lithgow, G.J. Lifespan extension in C. elegans by a molecular chaperone dependent upon insulin-like signals. Aging Cell 2003, 2, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Rea, S.L.; Wu, D.; Cypser, J.R.; Vaupel, J.W.; Johnson, T.E. A stress-sensitive reporter predicts longevity in isogenic populations of Caenorhabditis elegans. Nat. Genet. 2005, 37, 894–898. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Treatment | Mean ± SEM (Days) | % Change | N (Censored) | EXP | p | 95% CI for Mean | |
|---|---|---|---|---|---|---|---|---|
| Lower Bound | Upper Bound | |||||||
| WT, 20 °C | None | 15.47 ± 0.118 | 1132 (168) | 13 | 15.11 | 15.83 | ||
| MDHB (0/4) 160 mg/L | 18.41 ± 0.323 | +19 | 346 (54) | 4 | 0.008 | 17.76 | 19.06 | |
| MDHB (4/0) 160 mg/L | 15.02 ± 0.235 | −2.9 | 372 (28) | 4 | 0.618 | 14.56 | 15.49 | |
| RES 100 μM | 17.56 ± 0.316 | +13.5 | 352 (48) | 4 | 0.012 | 16.94 | 18.18 | |
| RAPA 100 μM | 18.76 ± 0.332 | +21.3 | 367 (33) | 4 | 0.003 | 18.11 | 19.40 | |
| W101 | None | 13.30 ± 0.240 | −16 | 512 (88) | 6 | 12.83 | 13.78 | |
| MDHB | 14.84 ± 0.276 | +11.6 | 489 (111) | 6 | 0.018 | 14.30 | 15.38 | |
| W203 | None | 13.01 ± 0.260 | −16 | 508 (92) | 6 | 12.50 | 13.52 | |
| MDHB | 14.03 ± 0.235 | +7.9 | 492 (108) | 6 | 0.042 | 13.57 | 14.52 | |
| W213 | None | 13.78 ± 0.270 | −11 | 522 (78) | 6 | 13.25 | 14.32 | |
| MDHB | 15.01 ± 0.281 | +9 | 501 (99) | 6 | 0.038 | 14.46 | 15.56 | |
| daf-16 (mu86) | None | 13.15 ± 0.337 | −15 | 546 (54) | 6 | 12.55 | 13.75 | |
| MDHB | 14.07 ± 0.306 | +7 | 523 (77) | 6 | 0.036 | 13.41 | 14.72 | |
| daf-2 (e1370) | None | 29.24 ± 0.469 | +89 | 532 (68) | 6 | 28.32 | 30.16 | |
| MDHB | 30.06 ± 0.459 | +2.8 | 513 (87) | 6 | 0.512 | 29.16 | 30.96 | |
| eat-2 (ad1113) | None | 18.77 ± 0.265 | +21.3 | 576 (24) | 6 | 18.25 | 19.29 | |
| MDHB | 22.60 ± 0.316 | +20.4 | 543 (57) | 6 | 0.039 | 21.98 | 23.22 | |
| Cosmid No. | Gene Symbol | Brief Description | MDHB Treatment | |
|---|---|---|---|---|
| Fold Change | p Value | |||
| T19E7.2 | skn-1 | orthologous to the mammalian Nrf (Nuclear factor-erythroid-related factor) transcription factors | 2.01 | <0.001 |
| F55G11.5 | dod-22 | DUF141 domain of unknown function, high similarity to uncharacterized C. elegans K10D11.2 | −2.22 | 0.0023 |
| T22A3.2 | hsp-12.1 | a member of the small heat shock family of proteins | 2.27 | 0.0045 |
| T27E4.8 | hsp-16.1 | Member of the C. elegans hsp-16 family | 4.32 | <0.001 |
| T27E4.3 | hsp-16.49 | Hsp20/alpha crystallin family, similar to alpha-B crystalline | 4.61 | <0.001 |
| K11G9.6 | mtl-1 | Metallothionein-related cadmium-binding protein | 2.06 | 0.0083 |
| T28B8.2 | ins-18 | Insulin-like protein of the type-β subfamily; may be a ligand for the daf-2 receptor | −2.04 | 0.0421 |
| K12G11.3 | sodh-1 | Alcohol dehydrogenase 1, have oxidoreductase activity | 2.64 | <0.001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mi, X.-N.; Wang, L.-F.; Hu, Y.; Pan, J.-P.; Xin, Y.-R.; Wang, J.-H.; Geng, H.-J.; Hu, S.-H.; Gao, Q.; Luo, H.-M. Methyl 3,4-Dihydroxybenzoate Enhances Resistance to Oxidative Stressors and Lifespan in C. elegans Partially via daf-2/daf-16. Int. J. Mol. Sci. 2018, 19, 1670. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061670
Mi X-N, Wang L-F, Hu Y, Pan J-P, Xin Y-R, Wang J-H, Geng H-J, Hu S-H, Gao Q, Luo H-M. Methyl 3,4-Dihydroxybenzoate Enhances Resistance to Oxidative Stressors and Lifespan in C. elegans Partially via daf-2/daf-16. International Journal of Molecular Sciences. 2018; 19(6):1670. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061670
Chicago/Turabian StyleMi, Xiang-Nan, Li-Fang Wang, Yang Hu, Jun-Ping Pan, Yi-Rong Xin, Jia-Hui Wang, Hai-Ju Geng, Song-Hui Hu, Qin Gao, and Huan-Min Luo. 2018. "Methyl 3,4-Dihydroxybenzoate Enhances Resistance to Oxidative Stressors and Lifespan in C. elegans Partially via daf-2/daf-16" International Journal of Molecular Sciences 19, no. 6: 1670. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061670