Suppression of Plant Defenses by Herbivorous Mites Is Not Associated with Adaptation to Host Plants

 ,
,  and
and 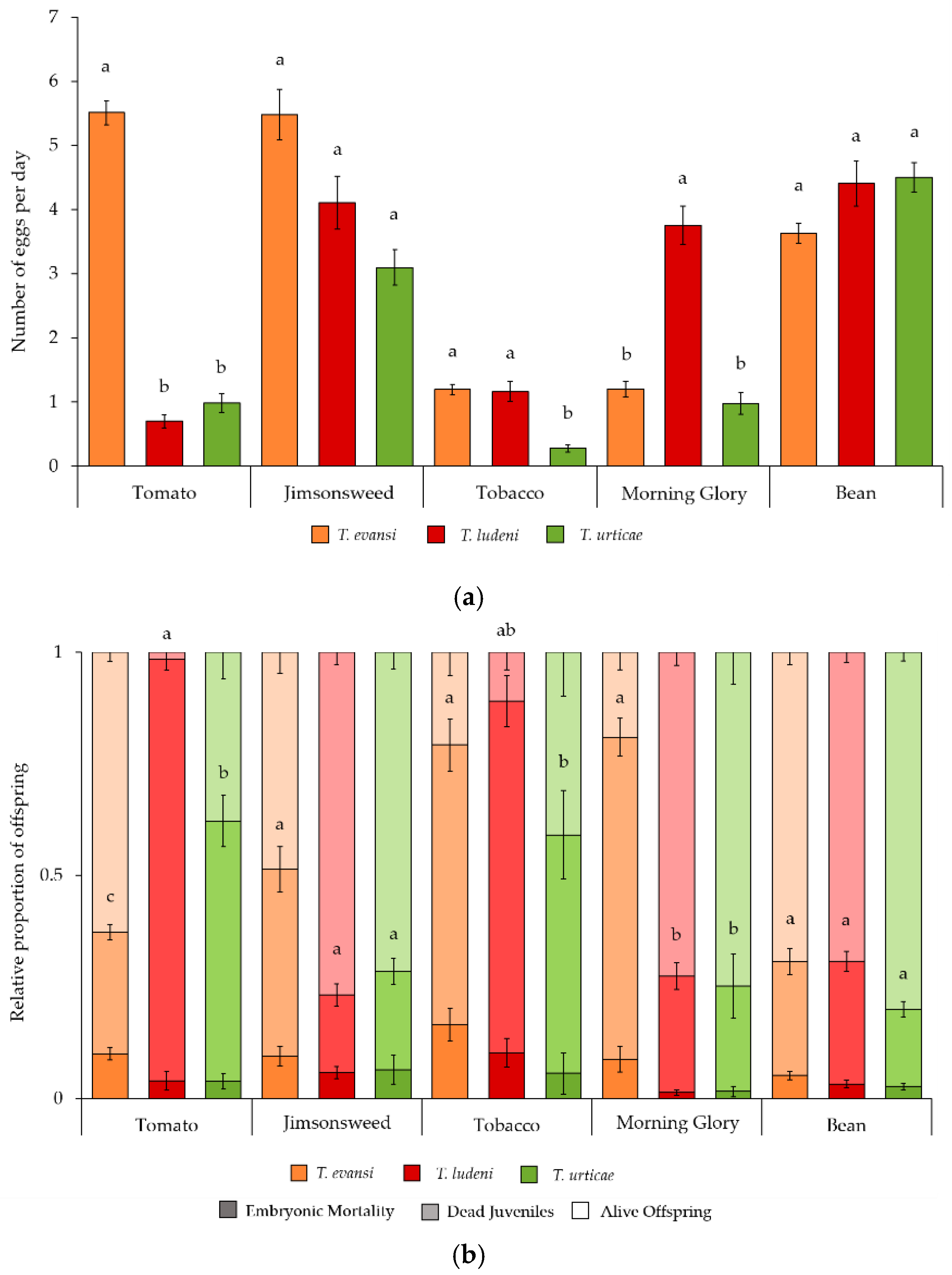{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Performance of Tetranychus Species on Several Host Plants
2.2. The Effect of T. evansi Infestations on Plant Defences
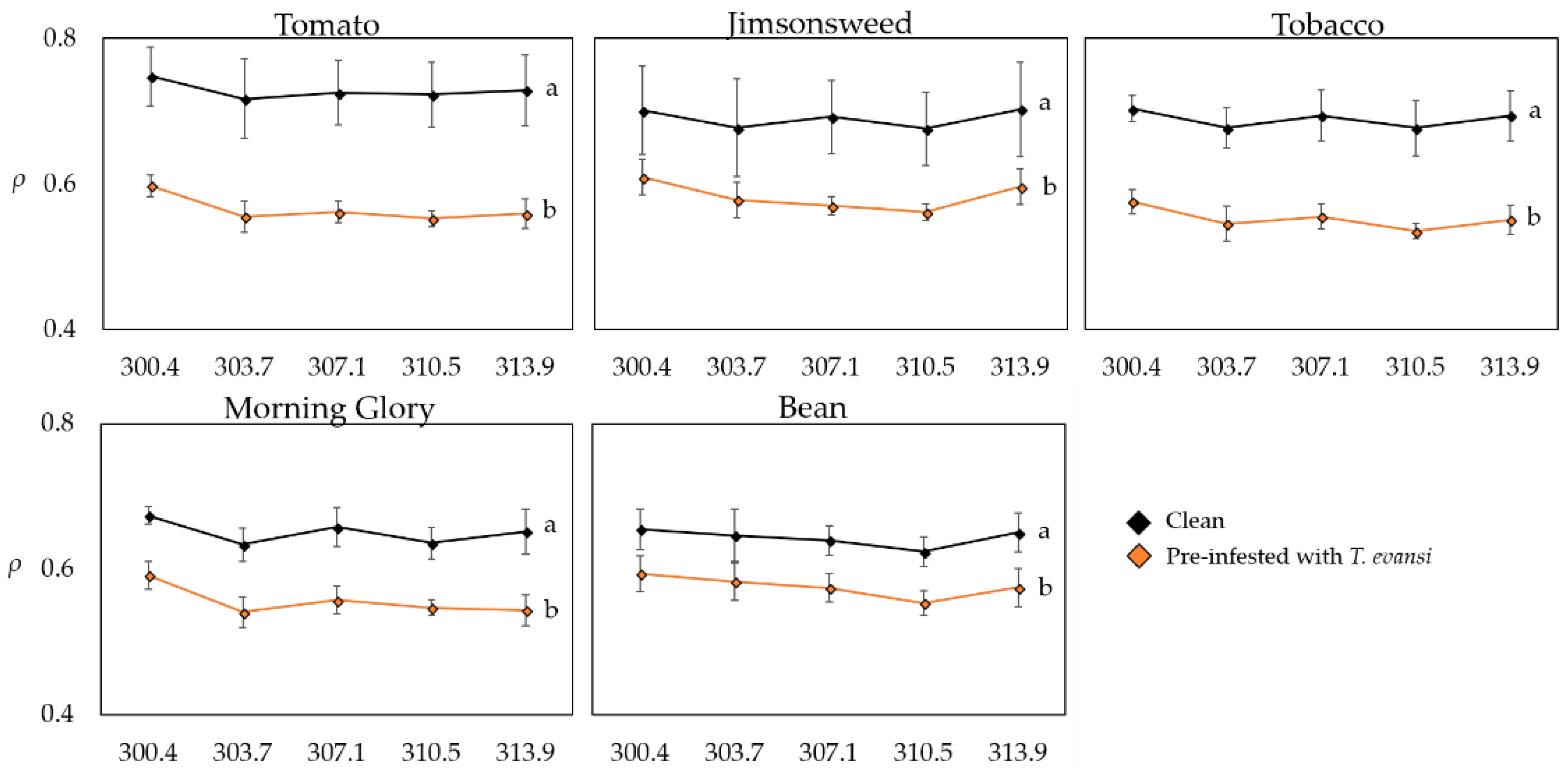
2.2.1. Reflectance Spectroscopy Analysis
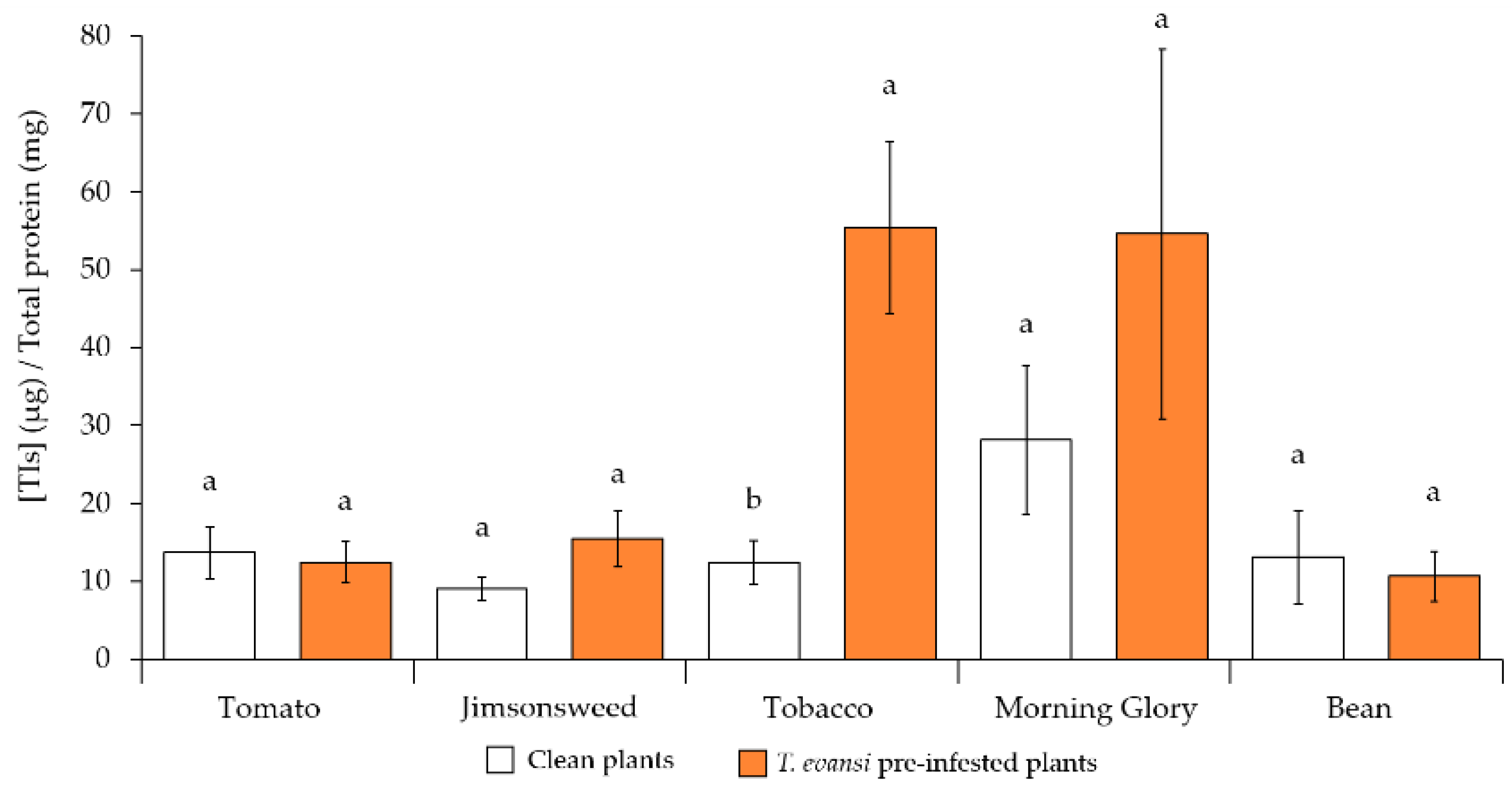
2.2.2. Trypsin Inhibitors Quantification
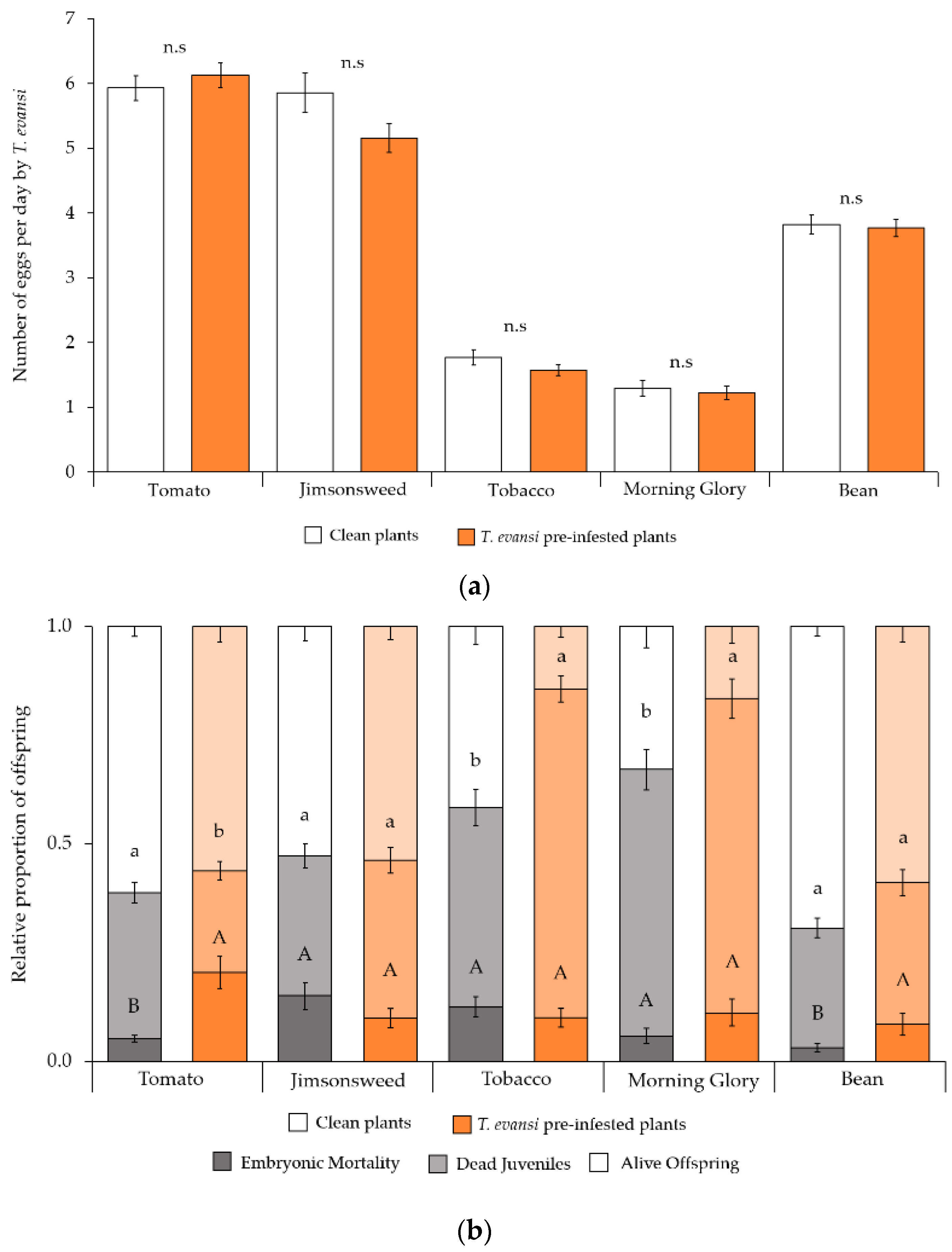
2.2.3. Conspecific Performance
3. Discussion
4. Materials and Methods
4.1. Plants
4.2. Spider Mite Cultures
4.3. Performance of Tetranychus Species on Several Host Plants
4.4. The Effect of T. evansi Infestations on Plant Defences
4.4.1. Reflectance Spectroscopy Analysis
4.4.2. Trypsin Inhibitors Quantification
4.4.3. Conspecific Performance
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| JA | Jasmonic Acid |
| TI | Trypsin Inhibitors |
| BApNA | N-Benzoyl-d,l-arginin-4 nitroamilide hydrochloride |
References
- Strauss, S.Y.; Agrawal, A.A. The ecology and evolution of plant tolerance to herbivory. Trends Ecol. Evol. 1999, 14, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Karban, R.; Myers, J.H. Induced Plant Responses to Herbivory. Annu. Rev. Ecol. Syst. 1989, 20, 331–348. [Google Scholar] [CrossRef]
- Karban, R.; Agrawal, A.A. Herbivore Offense. Annu. Rev. Ecol. Syst. 2002, 33, 641–664. [Google Scholar] [CrossRef]
- Ehrlich, P.R.; Haven, P.H. Butterflies and Plants: A Study in Coevolution. Evolution 1964, 18, 586–608. [Google Scholar] [CrossRef]
- Chapman, R.F. Contact chemoreception in feeding by Phytophagous Insects. Annu. Rev. Entomol. 2003, 48, 455–484. [Google Scholar] [CrossRef] [PubMed]
- Zagrobelny, M.; Bak, S.; Vinther, A. Cyanogenic glucosides and plant–insect interactions. Phytochemistry 2004, 65, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Paschold, A.; Halitschke, R.; Baldwin, I.T. Using ‘mute’ plants to translate volatile signals. Plant J. 2006, 45, 275–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, J.L.; Snyder, M.J.; Koener, J.F.; Feyereisen, R. Inducible P450s of the CYP9 family from larval Manduca sexta midgut. Insect Biochem. Mol. Biol. 2000, 30, 559–568. [Google Scholar] [CrossRef]
- Cianfrogna, J.A.; Zangerl, A.R.; Berenbaum, M.R. Dietary and developmental influences on induced detoxification in an oligophage. J. Chem. Ecol. 2002, 28, 1349–1364. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Jasmonate and salicylate induce expression of herbivore cytochrome P450 genes. Nature 2002, 419, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Musser, R.O.; Hum-musser, S.M.; Eichenseer, H.; Peiffer, M.; Murphy, J.B.; Felton, G.W. Caterpillar saliva beats plant defences. Nature 2002, 416, 599–600. [Google Scholar] [CrossRef] [PubMed]
- Kant, M.R.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Intraspecific variation in a generalist herbivore accounts for differential induction and impact of host plant defences. Proc. R. Soc. B 2008, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Oliver, K.M.; Xie, W.; Wu, Q.; Wang, S.; Zhang, Y. The whitefly-associated facultative symbiont Hamiltonella defensa suppresses induced plant defences in tomato. Funct. Ecol. 2015, 29, 1007–1018. [Google Scholar] [CrossRef]
- Soler, R.; Badenes-Pérez, F.R.; Broekgaarden, C.; Zheng, S.J.; David, A.; Boland, W.; Dicke, M. Plant-mediated facilitation between a leaf-feeding and a phloem-feeding insect in a brassicaceous plant: From insect performance to gene transcription. Funct. Ecol. 2012, 26, 156–166. [Google Scholar] [CrossRef]
- Sarmento, R.A.; Lemos, F.; Bleeker, P.M.; Schuurink, R.C.; Pallini, A.; Oliveira, M.G.A.; Lima, E.R.; Kant, M.; Sabelis, M.W.; Janssen, A. A herbivore that manipulates plant defence. Ecol. Lett. 2011, 14, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godinho, D.P.; Janssen, A.; Dias, T.; Cruz, C.; Magalhães, S. Down-regulation of plant defence in a resident spider mite species and its effect upon con- and heterospecifics. Oecologia 2016, 180, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Bos, J.I.B. Effector proteins that modulate plant-insect interactions. Curr. Opin. Plant Biol. 2011, 14, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Alba, J.M.; Glas, J.J.; Schimmel, B.C.J.; Kant, M.R. Avoidance and suppression of plant defenses by herbivores and pathogens. J. Plant Interact. 2011, 6, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Lortzing, T.; Steppuhn, A. Jasmonate signalling in plants shapes plant-insect interaction ecology. Curr. Opin. Insect Sci. 2016, 14, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A.; Baldwin, I.T. Plant Responses to Insect Herbivory: The EmergingMolecular Analysis. Annu. Rev. Plant Biol. 2002, 53, 299–328. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Saona, C.R.; Musser, R.O.; Vogel, H.; Hum-Musser, S.M.; Thaler, J.S. Molecular, Biochemical, and Organismal Analyses of Tomato Plants Simultaneously Attacked by Herbivores from Two Feeding Guilds. J. Chem. Ecol. 2010, 36, 1043–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidart-Bouzat, M.G.; Kliebenstein, D. An ecological genomic approach challenging the paradigm of differential plant responses to specialist versus generalist insect herbivores. Oecologia 2011, 167, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Musser, R.O.; Cipollini, D.F.; Hum-musser, S.M.; Williams, S.A.; Brown, J.K.; Felton, G.W. Evidence That the Caterpillar Salivary Enzyme Glucose Oxidase Provides Herbivore Offense in Solanaceous Plants. Arch. Insect Biochem. Physiol. 2005, 137, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Mothes, U.; Seitz, K.A. Fine structure and function of the prosomal glands of the two-spotted spider mite, Tetranychus urticae (Acari, Tetranychidae). Cell Tissue Res. 1981, 221, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Helle, W.; Sabelis, M.W. Spider Mites Their Biology, Natural Enemies and Control; Elsevier: Amsterdam, The Netherlands, 1985; Volume 1A, ISBN 0444423729. [Google Scholar]
- Kant, M.R.; Ament, K.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Differential Timing of Spider Mite-Induced Direct and Indirect Defenses in Tomato Plants. Plant Physiol. 2004, 135, 483–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wybouw, N.; Zhurov, V.; Martel, C.; Bruinsma, K.A.; Hendrickx, F.; Grbić, V.; Van Leeuwen, T. Adaptation of a polyphagous herbivore to a novel host plant extensively shapes the transcriptome of herbivore and host. Mol. Ecol. 2015, 24, 4647–4663. [Google Scholar] [CrossRef] [PubMed]
- Kant, M.R.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.C.J.; Villarroel, C.A.; Ataide, L.M.S.; Dermauw, W.; Glas, J.J.; et al. Mechanisms and ecological consequences of plant defence induction and suppression in herbivore communities. Ann. Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C. Resistance of Cultivated Tomato to Cell Content-Feeding Herbivores Is Regulated by the Octadecanoid-Signaling Pathway. Plant Physiol. 2002, 130, 494–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ament, K.; Kant, M.R.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Jasmonic acid is a key regulator of spider mite-induced volatile terpenoid and methyl salicylate emission in tomato. Plant Physiol. 2004, 135, 2025–2037. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, M.E.; Cambra, I.; Martinez, M.; Pozancos, C.; González-Melendi, P.; Grbic, V.; Castañera, P.; Ortego, F.; Diaz, I. Gene pyramiding of peptidase inhibitors enhances plant resistance to the spider mite Tetranychus urticae. PLoS ONE 2012, 7, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Alba, J.M.; Schimmel, B.C.J.; Glas, J.J.; Ataide, L.M.S.; Pappas, M.L.; Villarroel, C.A.; Schuurink, R.C.; Sabelis, M.W.; Kant, M.R. Spider mites suppress tomato defenses downstream of jasmonate and salicylate independently of hormonal crosstalk. New Phytol. 2015, 205, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Eich, E. Solanaceae and Convolvulaceae: Secondary Metabolites; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008; ISBN 9788578110796. [Google Scholar]
- Migeon, A.; Nouguier, E.; Dorkeld, F.; Cbgp, U.M.R.; Ird, I.; Montpellier, C.; Cedex, M. Spider Mites Web: A comprehensive database for the Tetranychidae Alain. Trends Acarol. 2010, 557–560. [Google Scholar] [CrossRef]
- Zélé, F.; Santos, I.; Olivieri, I.; Weill, M.; Duron, O.; Magalhães, S. Endosymbiont diversity and prevalence in herbivorous spider mite populations in South-Western Europe. FEMS Microbiol. Ecol. 2018, 94, fiy015. [Google Scholar] [CrossRef] [PubMed]
- Izaguirre, M.M.; Scopel, A.L.; Baldwin, I.T.; Ballare, C.L. Convergent Responses to Stress. Solar Ultraviolet-B Radiation and Manduca sexta Herbivory Elicit Overlapping Transcriptional Responses in Field-Grown Plants of Nicotiana longiflora. Plant Physiol. 2003, 132, 1755–1767. [Google Scholar] [CrossRef] [PubMed]
- Izaguirre, M.M. Solar Ultraviolet-B Radiation and Insect Herbivory Trigger Partially Overlapping Phenolic Responses in Nicotiana attenuata and Nicotiana longiflora. Ann. Bot. 2007, 99, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Zavala, J.A.; Patankar, A.G.; Gase, K.; Baldwin, I.T. Constitutive and inducible trypsin proteinase inhibitor production incurs large fitness costs in Nicotiana attenuata. Proc. Natl. Acad. Sci. USA 2004, 101, 1607–1612. [Google Scholar] [CrossRef] [PubMed]
- Haruta, M.; Major, I.T.; Christopher, M.E.; Patton, J.J.; Constabel, C.P.A. Kunitz trypsin inhibitor gene family from trembling aspen (Populus tremuloides Michx.): Clonning, functional expression, and induction by wounding and herbivory. Plant Mol. Biol. 2001, 1, 181–191. [Google Scholar]
- Reinbothe, S.; Mollenhauer, B.; Reinbothe, C. JIPs and RIPs: The Regulation of Plant Gene Expression by Jasmonates in Response to Environmental Cues and Pathogens. Plant Cell 1994, 6, 1197–1209. [Google Scholar] [CrossRef] [PubMed]
- Walker-Simmons, M.; Ryan, C.A. Wound-induced Accumulation of Trypsin Inhibitor Activities in Plant Leaves: Survey of Several Plant Genera. Plant Physiol. 1977, 59, 437–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, K.-W.; Lin, M.-I.; Tuan, S.-J.; Chen, Y.-M.; Lin, C.-J.; Kao, S.-S. Sweet potato (Ipomoea batatas) trypsin inhibitors expressed in transgenic tobacco plants confer resistance against Spodoptera litura. Plant Cell Rep. 1997, 16, 696–699. [Google Scholar] [CrossRef]
- Thaler, J.S. Jasmonate-inducible plant defences cause increased parasitism of herbivores. Nature 1999, 399, 17–19. [Google Scholar] [CrossRef]
- Cipollini, D. Environmental and developmental regulation of trypsin inhibitor activity in Brassica napus. J. Chem. Ecol. 2000, 26, 1411–1422. [Google Scholar] [CrossRef]
- Cipollini, D.F.; Bergelson, J. Plant density and nutrient availability constrain constitutive and wound-induced expression of trypsin inhibitors in Brassica napus. J. Chem. Ecol. 2001, 27, 593–610. [Google Scholar] [CrossRef] [PubMed]
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, I.T.; Jena, D.; Jagger, M. An Ecologically Motivated Analysis of Plant-Herbivore Interactions in Native Tobacco. Plant Physiol. 2001, 127, 1449–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchet, E.; Egas, M.; Olivieri, I.; Magalha, S. Environmental effects on the detection of adaptation. J. Evol. Biol. 2011, 24, 2653–2662. [Google Scholar] [CrossRef] [Green Version]
- Santamaría, M.E.; González-Cabrera, J.; Martínez, M.; Grbic, V.; Castañera, P.; Díaz, I.; Ortego, F. Digestive proteases in bodies and faeces of the two-spotted spider mite, Tetranychus urticae. J. Insect Physiol. 2015, 78, 69–77. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, E.F.; Pallini, A.; Janssen, A. Herbivores with similar feeding modes interact through the induction of different plant responses. Oecologia 2016, 180, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kassell, B.; Radicevic, M.; Berlow, S.; Pea, R.J.; Laskowski, M. Bovine Trypsin-Kallikrein Inhibitor (Kunitz Inhibitor, Basic Pancreatic Trypsin Inhibitor, Polyvalent Inhibitor from Bovine Organs); Academic Press: San Francisco, CA, USA, 1970; Volume 230. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Therneau, T.M. coxme: Mixed Effects Cox Models. R-Package Descr. 2012, 1–14. [Google Scholar]
- Crawley, M.J. The R Book; John Wiley & Sons: Chichester, UK, 2012; ISBN 9780470510247. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2014, 67, 1–48. [Google Scholar] [CrossRef]
- Fournier, D.A. AD Model Builder: Using automatic differentiation for statistical inference of highly parameterized complex nonlinear models. Optim. Methods Softw. 2012, 27, 233–249. [Google Scholar] [CrossRef] [Green Version]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paulo, J.T.; Godinho, D.P.; Silva, A.; Branquinho, C.; Magalhães, S. Suppression of Plant Defenses by Herbivorous Mites Is Not Associated with Adaptation to Host Plants. Int. J. Mol. Sci. 2018, 19, 1783. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061783
Paulo JT, Godinho DP, Silva A, Branquinho C, Magalhães S. Suppression of Plant Defenses by Herbivorous Mites Is Not Associated with Adaptation to Host Plants. International Journal of Molecular Sciences. 2018; 19(6):1783. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061783
Chicago/Turabian StylePaulo, Jéssica T., Diogo P. Godinho, Anabela Silva, Cristina Branquinho, and Sara Magalhães. 2018. "Suppression of Plant Defenses by Herbivorous Mites Is Not Associated with Adaptation to Host Plants" International Journal of Molecular Sciences 19, no. 6: 1783. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061783